(Actinopterygii Klein, 1885) Класс Актиноптеригии, Лучепёрые рыбы, Class Actinopterygii Klein, 1885 (Ray-finned fishes) 46 отрядов
Материал подготовил Фактистов И. Е., 2022 г. Сайт www.fishbiosystem.ru
Лучепёрые рыбы (лат. Actinopterygii) — класс рыб из группы костных рыб. Подавляющее большинство известных современных видов рыб (свыше 20 000 или около 95 %) относятся к лучепёрым. Живут в морских и пресных водах по всему миру. Многие лучепёрые рыбы являются объектом промысла. Размеры живущих ныне видов колеблются от 8 мм (Paedocypris) до 11 м (ремнетелые), а вес достигает 2300 кг (рыба-луна).
Научное название класса Actinopterygii происходит от греческих слов ακτις (aktis) — «луч» и πτερον (pteron) — «перо», «крыло» и связано со строением плавников этих рыб (центральная ось базальных элементов скелета в парных плавниках отсутствует).
Наиболее древняя находка ископаемых лучепёрых рыб имеет возраст 420 млн лет (конец Силурийского периода). Это хищная рыба Andreolepis hedei из отряда палеонискообразных (Palaeonisciformes). Останки этого вида были найдены в России, Швеции и Эстонии.
В пермском и триасовом периодах были широко распространены лучепёрые рыбы, обладавшие ганоидной чешуей. На смену им около 200 млн лет назад появились костистые рыбы, к которым в настоящий момент относят подавляющее большинство (95 %) современных рыб. В ходе длительной эволюции, сопровождавшейся широкой адаптивной радиацией, возникло большое разнообразие лучепёрых рыб.
Класс лучепёрых рыб разделяют на два подкласса: хрящевые ганоиды, или хрящекостные рыбы (Chondrostei) и новопёрые рыбы (Neopterygii). Первый подкласс включает в себя более древние по происхождению отряды рыб. Большинство же отрядов принадлежит к более молодой группе — новопёрым рыбам. В этом подклассе выделяются 2 инфракласса: костные ганоиды (Holostei) и костистые рыбы (Teleostei).
Лучепёрые рыбы встречающиеся в водах Российской Федерации (см. Рыбы России). Среди самых крупных таксонов отечественной ихтиофауны следует назвать отряды Perciformes — окунеобразные (418 видов, 4,2%) и Scorpaeniformes — скорпенообразные (378 видов, 25,6%). Представители этих двух отрядов составляют более половины (54,8%) всей отечественной ихтиофауны, хотя и здесь немало тех, которые очень редки или встречаются в наших территориальных водах только в летний период и не каждый год. Большинство из них морские обитатели и представляют дальневосточную фауну. Среди пресноводных представителей первенство удерживает отряд Cypriniformes — карпообразные (149 видов, 6,2%).
Различные размеры и измерения рыбы играют очень важную роль в ихтиологии, систематике и таксономии рыб. Часть признаков, отличающих различные виды или роды рыб — это разные размеры рыб и их соотношение.
Существуют определённые постоянные измерения:
SL (standart length) — стандартная длина. Длина рыбы от начала головы (с закрытым ртом) до начала хвостового плавника (если быть более точным — до окончания гипуральной кости). В большинстве специализированных статей (включая статьи с первичным описанием вида) используется именно стандартная длина.
TL (total length) — общая длина. Длина рыбы от начала головы (с закрытым ртом) до конца хвостового плавника (в сложенном положении). У рыб с различными отростками на хвостовом плавнике (например, меченосец) измерение проводится до конца основной части хвостового плавника. Дополнительные отростки в расчет не принимаются.
Два этих показателя, SL и TL, являются основными показателями для описания рыб. В профессиональной литературе в подавляющем большинстве случаев используется стандартная длина рыбы (SL). В аквариумистике очень часто переписывается эта длина для указания размера рыбки, но не бозначается, что это стандартная длина. Часто она выдается, как общая длина (TL).

Кроме двух основных, существует немало вспомогательных размеров рыб. Здесь приводится только часть из них.
1. (fork length) — длина рыбки от начала головы (с закрытым ртом) до выемки хвостового плавника (у рыб с раздвоённым хвостовым плавником).
2. Длина основания спинного плавника. Измерение производится от основания первого луча спинного плавника до основания последнего луча.
3. Длина головы. Измеряется от начала головы (с закрытым ртом) до заднего края жаберной крышки.
4. Посторбитальная длина (postorbital length). Измеряется от заднего края глаза до заднего края жаберной крышки.
5. Диаметр глаза.
6. Длина носа (рыла). Измеряется от начала головы (с закрытым ртом) до переднего края глаза.
7. Длина челюсти. Измеряется от начала рта (закрытого) до заднего края верхней челюсти.
8. Добрюшная длина (prepelvic length). Измеряется от начала головы (с закрытым ртом) до переднего края основания брюшных плавников.
9. Длина грудного плавника. Измеряется от основания до края грудного плавника.
10. Длина основания анального плавника. Измеряется от основания первого луча до основания последнего луча анального плавника.
11. Длина хвостового стебля. Измеряется от заднего края основания анального плавника до начала хвостового плавника (правильнее — до края гипуральной кости).
12. Наименьшая высота корня хвоста.
13. Высота тела рыбы. Измеряется в наиболее высоком месте тела рыбы.
При описании нового вида рыбы обязательно даются большинство из этих показателей, а также соотношение их друг с другом (чаще всего со стандартной длиной — SL). Без знания этих терминов понять описание нового вида и отличие его от похожих видов довольно трудно.
Ихтиопланктон Черного моря. Т.В.Дехник 1973 г.
СИСТЕМАТИЗАЦИЯ РЫБ ЧЕРНОГО МОРЯ ПО СПОСОБАМ РАЗМНОЖЕНИЯ
По новейшим данным ихтиофауна Черного и Азовского морей включает 163 вида и подвида (Расс, 1949, 1965; Виноградов, 1960; Драпкин, 1961; Nalbant, 1962; Стоянов, 1963; Световидов, 1964). Н. М. Книпович (1933) и Т. С. Расс (1949, 1965) различают в составе ихтиофауны по происхождению и экологии три группы: проходные и полупроходные (24 вида), солоноватоводные (22 вида) и собственно морские виды , которых Т. С. Расс (1949, 1965) разделяет на умеренно-водные — бореально-атлантические реликты (8 видов) и тепловодные — южнобореальные, субтропические и тропические (109 видов).
Собственно морских рыб Черного моря В. А. Водяницкий (1930б) делит по способам размножения на четыре группы: 1) живородящие; 2) вынашивающие икру в специальных выводковых камерах; 3) рыбы с пелагической икрой; 4) рыбы с демерсальной икрой. В список рыб, размножение которых в Черном море достоверно известно или очень вероятно, В. А. Водяницкий (1930б) включил 75 видов. В дальнейшем этот список уточнялся и дополнялся (Водяницкий, 1936; Пчелина, 1936, 1940; Косякина, 1936, 1938; Дехник и Павловская, 1950; Зайцев, 1959а; Овен, 1959). Накапливались новые данные по видовому составу, распределению, условиям развития икры и личинок рыб. Проведенные автором в течение 10 лет (1957—1967 гг.) исследования размножения и развития рыб Черного моря и имеющиеся литературные данные позволили систематизировать рыб по их экологическим особенностям размножения и уточнить список рыб, достоверно размножающихся в Черном море.
По способам размножения собственно морские рыбы Черного моря могут быть разделены на пять групп. Три первые группы соответствуют выделенным В. А. Водяницким (1930б); четвертую группу составляют рыбы, откладывающие (разбрасывающие) икру (яйца) на грунт, водоросли или различные подводные предметы. К пятой группе отнесены гнездовые рыбы, охраняющие потомство и имеющие тоже донную икру, не отличающиеся от рыб предыдущей группы рядом биологических особенностей. В частности, у самцов этой группы рыб в период нереста обычно появляется брачный наряд; в их поведении проявляется «агрессивность», определяемая заботой о потомстве; вырабатывается «строительный» инстинкт, который у некоторых видов выражается в постройке искусных сооружений (губановые, бычки, колюшка).
В прилагаемом ниже списке видов приняты следующие условные обозначения:
1) рыбы, достоверно размножающиеся в Черном море (+);
2) размножающиеся в Черном и Азовском морях (+ X);
3) размножающиеся только в Азовском море (X);
4) единичные случаи нахождения в Черном море икры, личинок или текучих особей (1);
5) размножение в Черном море не установлено (?);
6) рыбы, не размножающиеся в Черном море (0);
7) нахождение в Черном море нуждается в подтверждении (!);
8) принадлежность к указанной группе нуждается в подтверждении (п).



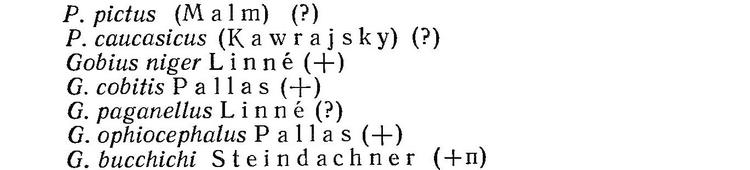
Изучение строения, развития и экологии икры и личинок рыб позволило уточнить имеющиеся в литературе представления о размножении некоторых видов в Черном море.
Так, выявленное сходство в строении икринок и только что выклюнувшихся личинок луфаря (Pomatomus saltatrix) и тунца (Thunnus thymus) позволяет предположить, что в определении были допущены ошибки. Отнесенные к тунцу икра и личинки (Водяницкий, 1936; Дука, 1959; Овен, 1959; Зайцев, 19596) в действительности принадлежат луфарю.
Выявленная морфологическая изменчивость выклюнувшихся личинок двух близких видов — Serranus scriba и Serranus cabrilla — заставляет с осторожностью подойти к имеющимся в литературе данным (Овен, 1959) о нахождении икринок второго чрезвычайно редко встречающегося в Черном море вида.
Анализ условий размножения и строения икринок Clupea pilchardus позволяет считать сомнительными данные (Водяницкий, 1930а) о нахождении в Черном море икринок этого вида. В то же время в список рыб с демерсальной икрой, достоверно размножающихся в Черном море, включен Tripterygion tripteronotus, личинки и мальки которого в большом количестве обнаружены в зарослях цистозиры (А. Д. Гордина, неопубликованные данные). Светлый горбыль (Umbrina cirrosa) отнесен нами к группе рыб с пелагической икрой. Исходя из строения овариальных икринок, более вероятно предположить, что икринки у светлого горбыля, как и у темного, — пелагические. В ранее опубликованных работах (Дехник, 1969; Дехник, Дука и др., 1970) соответствующие уточнения не были внесены.
Таким образом, из 117 собственно морских рыб, обитающих в Черном и Азовском морях (Расс, 1965), 63 вида и подвида проходят в своем развитии две планктонные фазы развития — фазу икринки и фазу личинки. Из этого количества в Черном море достоверно размножаются 28 видов и подвидов. Отмечены единичные случаи нахождения икринок, личинок или текучих особей двух видов (Zeus faber pungio, Trigla lucerna). Два подвида — азовская хамса (Engraulis encrasicholus maeoticus) и азовская камбала-калкан (Scophthalmus maeoticus torosus) — размножаются только в Азовском море.
Одну планктонную фазу — фазу личинки — проходят 40 видов и подвидов. В составе черноморской ихтиофауны насчитывается 28 видов и подвидов с донной икрой, пелагические личинки которых закономерно или спорадически встречаются в планктоне.
ЭТАПЫ И СТАДИИ НАЧАЛЬНОГО ОНТОГЕНЕЗА РЫБ
В эмбриональном и постэмбриональном развитии рыб различают четыре периода (фазы), отличающиеся морфологическими особенностями и всей биологией организма: зародышевый, предличиночный, личиночный и мальковый (Расс, 1946, 1948). Каждый из периодов имеет ряд этапов (стадий), характеризующихся определенными биологическими качествами (Расс, 1946; Матвеев, 1956).
Этапность эмбрионального развития рыб, особенно морских видов, остается до настоящего времени недостаточно изученной. Т. С. Расс (1946) выделяет четыре стадии развития икринок: 1) дробления и обрастания; 2) зародышевой полоски; 3) неоформившегося эмбриона и 4) оформившегося эмбриона. Б. С. Матвеев (1956) сделал попытку «наметить общие процессы в индивидуальном развитии различных животных, объединяющиеся специфическими приспособлениями к условиям существования». В эмбриональном развитии рыб (на примере осетровых) он различает шесть этапов, разграничивающихся морфологическими признаками: 1) формирование зиготы (от проникновения сперматозоида в яйцеклетку до начала дробления); 2) дробление — преобразование оплодотворенной яйцеклетки в многоклеточный организм; 3) гаструляция — процесс первичного морфогенеза зародышевых органов; 4) нейруляция — процесс образования трубчатой нервной системы, а также процесс сегментации тела; 5) обособление зародыша на икринке; 6) подвижность эмбриона (от начала пульсации сердца до выклева).
Этапы эмбрионального развития, выявленные нами у черноморской хамсы (Дехник, 1959) и прослеженные позднее на других рыбах (Дехник, 1961), в основном совпадают со схемой эмбриогенеза Б. С. Матвеева. В отличие от предложенной им схемы, процесс формирования зиготы мы объединяем с процессом дробления в I этап, а во II этап выделяем качественно новый процесс — процесс топографического перемещения бластомеров к поверхности с образованием бластоцеля — эпителиальную бластулу. Под термином «этап», как это введено В. В. Васнецовым (1948, 1953), следует понимать интервалы развития, имеющие определенные биологические качества. Термин «стадия» употребляется в понимании С. Г. Крыжановского (1949, 1953, 1956) для обозначения каждого данного момента развития.
В эмбриональном периоде развития подавляющего большинства морских рыб с пелагической икрой четко прослеживаются шесть последовательных этапов. Каждый этап характеризуется определенными качественными процессами и морфологическими особенностями. Развитие в пределах этапов и на переходах от одного этапа к другому характеризуется постепенностью (Крыжановский, 1953). Процессы роста (количественные изменения) тесно переплетаются с процессами дифференцировки (качественными изменениями).
В общей форме этапы эмбрионального развития морских рыб с пелагической икрой характеризуются следующими признаками:
I этап — дробление. Начинается от оплодотворения и заканчивается образованием многоклеточного зародышевого диска, или бластомерной бластулы.
II этап — эпителиальная бластула. Характеризуется смещением бластомеров в поверхностный слой и образованием многослойной пластинки, прикрывающей бластоцель.
III этап — обрастание желтка бластодиском и гаструляция. Начинается увеличением диаметра бластодиска и почти одновременным подворачиванием его краев. По мере развития формируется зародышевая полоска, закладываются зародышевые органы. Этап заканчивается замыканием бластопора.
IV этап — зародышевая полоска. Обособляется голова, образуется нервный тяж, намечается хорда, мозговые пузыри, глазные бокалы, слуховые капсулы; начинается сегментация тела.
V этап — рост хвостового отдела. Начинается отделением хвоста от желтка. На этом этапе образуется сердце, зачатки кишечника, печени. Намечается плавниковая кайма. С ростом хвостового отдела происходит дифференцировка хвостовых миотомов. Этап продолжается до начала пульсации сердца.
VI этап — подвижный эмбрион. Почти одновременно с пульсацией сердца начинаются слабые движения эмбриона, который время от времени вздрагивает, подергивается. По мере роста эмбриона и формирования личиночных органов движения становятся более энергичными. Перед выклевом эмбрион вместе с желтком свободно поворачивается в яйцевой оболочке; частота пульсации сердца возрастает.
Таким образом, принимаемые нами I и II этапы соответствуют I стадии Т. С. Расса, III и IV — II стадии, V и VI — III и IV стадиям (Расс, 1946, 1948).
В эмбриональном развитии рыб четко прослеживаются видовые особенности. Межвидовые различия касаются времени появления зачатков тех или иных органов, формы тела эмбриона, развития и характера пигментации. Существенные различия проявляются в длительности VI этапа, чем главным образом определяется степень развития эмбрионов перед выклевом и выклюнувшихся личинок.
В развитии демерсальных яиц рыб семейств Labridae и Pomacentridae выявлены те же 6 этапов (Салехова, 1971).
Выклев эмбриона сопровождается существенными изменениями экологии. Эмбрион освобождается от оболочки, до этого времени ограничивающей воздействие на него различных абиотических и биотических факторов среды. Он приобретает сразу же после выклева активную подвижность (разные способы и формы движения у разных видов) и как подвижный организм делается более привлекательным объектом для многочисленных планктонных хищников. Задолго до рассасывания желточного мешка (на 2—4-й день после выклева) начинается переход к внешнему питанию (этап смешанного питания). Таким образом, в этот период жизни проявляются сложные взаимоотношения, в которых выклюнувшийся эмбрион (личинка) является, с одной стороны, жертвой, с другой — хищником, активно добывающим пищу. Следовательно, после выклева наступает качественно новый период в жизни организма (личиночный), который резко отличается от предшествующего эмбрионального периода. Исходя из изложенного, на наш взгляд, не следует вводить для этапа желточного питания термин «выклюнувшийся эмбрион» (Крыжановкий, 1949, 1953, 1956), а более правильно, как это делают Т. С. Расс (1946), Б. С. Матвеев (1956), Т. А. Перцева-Остроумова (1961), ограничивать период эмбрионального развития моментом выклева эмбриона.
В личиночном периоде онтогенеза подавляющего большинства морских рыб мы различаем четыре этапа.
I этап — желточное питание. Личинки слабо развиты. Голова плотно прижата к большому желточному мешку. Грудные плавники зачаточные или отсутствуют. Глаза не пигментированы. Нерасчлененная плавниковая складка окаймляет тело. Личинки длительное время находятся в покое. Они занимают в воде вертикальное, косое (головой вниз или вверх) или горизонтальное (брюшной стороной вверх) положение. Движения в форме скачков в горизонтальной плоскости, вращательные вокруг оси тела, винтообразные в вертикальном направлении от дна к поверхности. Этап продолжается до образования ротового отверстия и появления пигмента в глазах, когда начинает проявляться поисковый инстинкт и в кишечниках появляются первые пищевые организмы.
II этап — смешанное питание. Желточный мешок заметно резорбирован. Образовалось ротовое отверстие; рот приобрел подвижность. Развиваются челюстной и жаберный аппараты. В глазах появился черный пигмент. Плавательный пузырь заполняется воздухом. Грудные плавники значительно увеличились в размере и приобрели подвижность, благодаря чему личинки начали двигаться в разных направлениях. Увеличивается скорость и продолжительность движения; значительно сократились периоды покоя. Появляется поисковый инстинкт. Личинки наряду с использованием желтка питаются внешней пищей, в основном мелкими малоподвижными формами зоопланктона (Tintinnidae, науплиусы Copepoda, копеподитные стадии Oithona minuta) и фитопланктоном (в основном Prorocentrum micans, Exuviaella cordata, Coscinodiscus sp.).
III этап — внешнее личиночное питание. Желточный мешок полностью резорбирован. Заканчивается формирование челюстного и жаберного аппаратов. Формируются лучи в непарных и грудных плавниках. Появляются зачатки брюшных плавников. Личинки приобретают способность к длительным целенаправленным движениям. При помощи подвижных хорошо развитых грудных плавников и сильных изгибов хвостового отдела личинки могут достаточно быстро и точно поворачиваться, производить стремительные скачки, ускользая от хищников или охотясь за пищей. Четко выражены хватательные движения рта. В пищевом комке появляются крупные зоопланктеры. Этап заканчивается формированием непарных плавников.
IV этап — переход к образу жизни взрослых форм. Этот этап и последующие периоды онтогенеза нами не изучались.
Описанная схема этапности личиночного периода жизни прослеживается для I группы выклюнувшихся личинок. У личинок многих видов, выклюнувшихся из донной икры, и у некоторых личинок, выклюнувшихся из пелагической икры, как это рассмотрено ниже, выявлены отклонения от приведенной схемы развития, в частности, может выпадать I этап или (у некоторых видов) I и II этапы личиночного периода.
Таким образом, предлагаемые нами I и II этапы соответствуют фазе предличинки согласно схеме, предложенной Т. С. Рассом (1946, 1948), или фазе выклюнувшегося эмбриона, по данным С. Г. Крыжановского (1949, 1953, 1956), III и IV этапы соответствуют фазе личинки по Т. С. Рассу.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПЕЛАГИЧЕСКИХ ИКРИНОК РЫБ ЧЕРНОГО МОРЯ
1 (6). Икринки овальной или слабо овальной формы.
2 (3). Икринки хорошо выраженной овальной формы; желток сегментирован; обычны в планктоне с конца апреля — начала мая до конца сентября — начала октября. Engraulis encrasicholus ponticus (рис. 4)
3 (2). Икринки слабо выраженной овальной формы; желток гомогенный.
4 (5). Тело эмбриона массивное; пигмента на теле нет; встречаются в планктоне с мая до конца сентября. Scorpaena porcus (рис. 55)
5 (4). Тело эмбриона тонкое, стройное; к моменту образования хвостовой почки на теле появляются меланофоры; встречаются в планктоне с июня до конца сентября. Ophidion rochei (рис. 45)
6 (1). Икринки сферические.
7 (18). Икринки без жировых капель.
8 (9). Икринки крупные диаметром более 1,5 мм; желток гомогенный; перивителлиновое пространство узкое; обычны в планктоне с конца мая до сентября. Uranoscopus scaber (рис. 40)
9 (8). Икринки диаметром менее 1,5 мм.
10 (13). Желток сегментирован.
11 (12). Желток крупно сегментирован; икринки средней величины, диаметром 0,90—1,35 мм; встречаются на всем пространстве Черного моря преимущественно зимой (декабрь — февраль), летом единичные икринки могут быть встречены ниже слоя температурного скачка. Sprattus sprattus phalericus (рис. 1)
12 (11). Мелко сегментирован поверхностный слой желтка; икринки мелкие, диаметром 0,61—0,81 мм; обычны в планктоне у берегов с июня по сентябрь. Callionymys sp. (рис. 47)
13 (10). Желток гомогенный.
14 (15). Икринки размером менее 1 мм; оболочка очень тонкая, прозрачная с розовым или фиолетовым отливом; встречаются в планктоне с середины мая до конца августа, наиболее часто — в июне. Ctenolabrus rupestris (рис. 33)
15 (14). Икринки размером более 1 мм; оболочка не имеет розового или фиолетового отлива; встречаются в планктоне преимущественно зимой, летом — ниже слоя температурного скачка.
16 (17). Диаметр икринок 1,05—1,35 мм; тело эмбриона прогонистое; диффузный каротиноидный пигмент на теле появляется в конце IV этапа; в это же время появляются меланофоры и ксантофоры на желточном мешке в области хвостовой почки; икринки встречаются преимущественно с января по апрель. Platichthys flesus luscus (рис. 61)
17(16). Диаметр икринок 1,10—1,33 мм; тело эмбриона массивное; каротиноидный пигмент появляется незадолго до выклева; на желточном мешке пигмента нет; икринки встречаются преимущественно с декабря по март. Odontogadus merlangus euxinus (рис. 9)
18 (7). Икринки с одной или несколькими жировыми каплями.
19 (22). Икринки с несколькими или многими жировыми каплями.
20 (21). Жировых капель 3—6; диаметр икринок 1,40—1,65 мм; желток гомогенный; икринки встречаются в планктоне в мае — июле, преимущественно в открытом море. Sarda sarda (рис. 50)
21 (20). Жировых капель много (несколько десятков); диаметр икринок 1,05—1,35 мм; желток сегментирован; икринки встречаются в планктоне с июня по сентябрь у берегов. Solea lascaris nasuta (рис. 63)
22 (19). Икринки с одной жировой каплей.
23 (30). Икринки крупные, их диаметр более 1,6 мм.
24 (27). Желток гомогенный.
25 (26). Диаметр икринок 1,96—2,00 мм, жировой капли — 0,35—0,40 мм. Zeus faber pungio (рис. 11, а, б)
В Черном море икринки не найдены; в Средиземном море нерест с июня по сентябрь.
26 (25). Диаметр икринок 2,13—2,36 мм, жировой капли — 0,53—0,57 мм. Lophius piscatorius
В Черном море нерест не отмечен; в Средиземном море нерест происходит зимой.
27 (24). Желток сегментирован.
28 (29). Диаметр икринок 1,6—1,8 мм, жировой капли — 0,4 мм; перивителлиновое пространство узкое. Xiphias gladius (рис. 53, а, б)
В Черном море икринки не найдены; в Средиземном море нерест с конца июня до августа.
29 (28). Диаметр икринок 1,5—1,7 мм, жировой капли — 0,16 мм; перивителлиновое пространство широкое. Sardina pilchardus (рис. 3, а)
В Черном море икринки не найдены; в Средиземном море разгар нереста зимой.
30 (23). Икринки среднего размера и мелкие, диаметром менее 1,6 мм.
31 (46). Икринки среднего размера диаметром от 1,0—1,1 до 1,6 мм.
32 (35). Желток сегментирован.
33 (34). Диаметр икринок 1,0—1,15 мм, жировой капли — 0,22—0,30 мм; перивителлиновое пространство узкое. Sphyraena sphyraena
В Черном море икринки не найдены; в Средиземном море нерест с июня по сентябрь.
34 (33). Диаметр икринок 1,20—1,40 мм, жировой капли — в среднем 0,121 мм; перивителлиновое пространство широкое. Sardinella aurita (рис. 3, в)
В Черном море не размножается; в Средиземном море нерест в основном с июля по сентябрь.
35 (32). Желток гомогенный.
36 (39). Икринки диаметром более 1,2—1,3 мм; жировая капля очень крупная, диаметром более 0,35 мм.
37 (38). Диаметр икринок 1,33—1,47 мм, жировой капли — 0,35—0,38 мм;
в Черном море икринки встречаются редко в июне — августе. Trigla lucerna (рис. 57)
38 (37). Диаметр икринок 1,32 мм, жировой капли — 0,44 мм. Mugil labrosus
В Черном море икринки не найдены; в Средиземном море период нереста достоверно не установлен.
39 (36). Диаметр икринок менее 1,3 мм, жировой капли — менее 0,35 мм.
40 (44). На теле эмбриона есть сгруппированные пигментные клетки (пятна), которые в отраженном свете имеют ярко-желтую, в проходящем — черную окраску.
41 (42). Диаметр икринок 1,07—1,19 мм, жировой капли — 0,29—0,36 мм. Черный пигмент преимущественно в туловищном отделе; скопления желтого пигмента (пятна) расположены за глазами, на нижней стороне туловища и в области жировой капли. Scomber scombrus (рис. 49)
В Черном море икринки не найдены; в Мраморном море нерест — с марта по май, в Средиземном — с января по апрель.
42 (41). Диаметр икринок 0,94—1,15 мм, жировой капли — 0,19—0,25 мм. После замыкания бластопора на теле эмбриона и на жировой капле появляются меланофоры; на VI этапе образуются характерные пигментные пятна за глазами и в хвостовом отделе на брюшной стороне; икринки обычны в планктоне с конца мая до середины сентября. Trachinus draco (рис. 38)
43 (40). Меланофоры и ксантофоры покрывают все тело эмбриона или часть его; сгруппированного в виде пятен пигмента нет.
44 (45). Диаметр икринок 1,15—1,25 мм, жировой капли — 0,23—0,25 мм. Меланофоры и ксантофоры густо расположены в туловищном отделе; в хвостовом отделе — редкие точечные меланофоры, карогиноидного пигмента нет; икринки встречаются в планктоне с июня до середины сентября. Sciaena umbra (рис. 25)
45 (44). Диаметр икринок 1,10—1,33 мм, жировой капли — 0,17—0,23 мм. По всему телу эмбриона густо расположены меланофоры и эритрофоры; икринки встречаются в планктоне в апреле — мае. Scophthalmus maeoticus maeoticus (рис. 58)
46 (31). Икринки мелкие, диаметром от 0,6 до 1,0—1,1 мм.
47 (56). Жировая капля крупная, диаметром более 0,25 мм.
48 (51). Диаметр икринок менее 0,80 (0,88) мм.
49 (50). Диаметр икринок 0,71—0,78 мм, жировой капли — 0,26—0,31 мм. На теле эмбриона рано появляются крупные ветвистые меланофоры; икринки встречаются с конца мая — начала июня до конца августа — начала сентября (иногда в октябре). Mugil cephalus (рис. 12)
50 (49). Диаметр икринок 0,76—0,80 (0,88) мм; жировой капли — 0,31—0,40 мм; тело эмбриона вначале пигментировано точечными редко рассеянными меланофорами. По мере развития пигментация усиливается, меланофоры становятся ветвистыми; икринки встречаются с июня по сентябрь. Mugil saliens (рис. 14)
51 (48). Диаметр икринок более 0,80 мм.
52 (53). Диаметр икринок 0,87—1,05 мм; жировая капля диаметром 0,31—0,37 мм с неровными очертаниями; ветвистые пигментные клетки распределены по всему телу эмбриона; четких пигментных рядов нет; икринки встречаются в планктоне с конца июля — начала августа до октября. Mugil auratus (рис. 13)
53 (52). Жировая капля хорошо очерченная с гладкой поверхностью; на спинной стороне тела ряд из крупных, редко расположенных меланофоров; ответвления меланофоров заходят на плавниковую кайму.
54 (55). Диаметр икринок 0,80—1,15 мм, жировой капли — 0,27—0,35 мм, желток крупно сегментирован у анимального полюса; на брюшной стороне хвостового отдела 1—2 меланофора; икринки закономерно встречаются в планктоне в июле и августе. Pomatomus saltatrix (рис. 20)
55 (54). Диаметр икринок 0,95—1,03 мм, жировой капли — 0,25—0,35 мм; желток гомогенный; на брюшной стороне хвостового отдела — пигментный ряд. Thunnus thynnus (рис. 52)
Размножение в Черном море достоверно не установлено; в Средиземном море нерест с апреля по июль.
56 (47). Жировая капля диаметром менее 0,25 (0,29) мм.
57 (60). Желток полностью или частично сегментирован.
58 (59). Диаметр икринок 0,73—1,00 мм; желток полностью сегментирован; жировая капля с неровными очертаниями, шероховатая, размером 0,19—0,25 (0,29) мм; пигмент на зародыше черный и каротиноидный; икринки встречаются в планктоне с конца мая до сентября. Trachurus mediterraneus ponticus (рис. 22)
Диаметр икринок 0,76—0,78 мм, жировой капли — 0,19—0,23 мм; в Черном море икринки не найдены; в Средиземном — нерест в течение всего года, разгар — летом. Trachurus trachurus trachurus (рис. 24, а, б)
59 (58). Диаметр икринок (0,61) 0,70—0,88 мм, сегментирован только поверхностный слой желтка; жировая капля обычно четко очерчена, блестящая, с гладкой оболочкой, размером 0,16—0,25 мм; пигмент только черный, икринки встречаются в планктоне с конца мая до сентября. Mullus barbatus ponticus (рис. 30)
60 (57). Желток гомогенный.
61 (64). Икринки очень мелкие, диаметром 0,60—0,70 мм.
62 (63). Диаметр икринок 0,59—0,70 мм, жировой капли — 0,10—0,13 мм; встречаются в планктоне с июня по август. Arnoglossus kessleri (рис. 60)
63 (62). Диаметр икринок 0,60—0,70 мм, жировой капли — 0,16—0,18 мм; в Черном море не найдены; в Средиземном — нерест с апреля по июль. Coris julis (рис. 35, а)
64 (61). Диаметр икринок более 0,75 мм.
65 (66). Диаметр икринок более 0,90 мм; жировая капля очень мелкая, ее диаметр не более 0,15 мм; икринки встречаются в планктоне с конца июня до середины сентября. Serranus scriba (рис. 17)
Диаметр икринок 0,92—0,97 мм, жировой капли — 0,14—0,15 мм; в Черном море икринки не найдены, в Средиземном — нерест с мая по август. Serranus cabrilla (рис. 19, а)
66 (65). Диаметр икринок от 0,75 до 0,90 мм — 1,0 мм, жировой капли — более (0,13) 0,15 мм.
67 (70). На теле зародыша имеется каротиноидный пигмент; икринки встречаются в планктоне летом.
68 (69). Диаметр икринок 0,75—0,97 мм, жировой капли — 0,15—0,20 мм; зародыш имеет характерную пигментацию: парные скопления пигмента (в проходящем свете — черного, в отраженном — желтого) впереди и позади глаз, в области слуховых капсул, у ануса и в середине хвостового отдела на брюшной стороне; икринки встречаются в планктоне с июня по сентябрь. Diplodus annularis (рис. 26)
Икринки диаметром 0,91—1,01 мм, жировой капли — 0,15—0,20 мм; в Черном море не найдены; и Средиземном — нерест с апреля по июнь. Diplodus sargus (рис. 24, д)
69 (68). Икринки в среднем несколько крупнее, их диаметр 0,87—1,0 мм, жировой капли — 0,17—0,22 мм; пигментация сходная, встречаются в планктоне с июня по сентябрь. Boops boops (рис. 28)
70 (67). На теле зародыша нет каротиноидного пигмента; икринки встречаются в планктоне с сентября по май, преимущественно в октябре —
декабре. Gaidropsarus mediterraneus (рис. 7.)
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Лучеперые рыбы (Actinopterygii)
впервые появились также в девонский период.
Парные плавники у этих рыб обычно не
имеют покрытой чешуей лопасти
основания, а поддерживаются многолучевым
скелетом без центральной оси для базальных
опорных элементов. К подклассу
лучеперых рыб относятся древние ганоидные
рыбы, сохранившие примитивные черты
строения внутреннего скелета и скелета
непарных плавников, чешуйного покрова
и пищеварительной системы, и костистые
рыбы, более прогрессивные по строению
и специализации, имеющие лучше
окостеневший скелет.
ПОДКЛАСС ЛУЧЕПЕРЫЕ РЫБЫ (ACTINOPTERYGII)
Лучеперые рыбы составляют свыше 95%
ныне живущих рыбообразных и рыб.
Опору осевого скелета у них образует, как
правило, костный позвоночник, только
у немногих сохраняется хорда или ее
остатки. Основания парных плавников не
носят характера вдающейся в плавник
лопасти, покрытой чешуей, или такая
лопасть невелика и скелет ее не имеет
опорной центральной оси, а образован рядом
элементов — радиалий. Чешуя у
большинства представляет собой тонкие костные
пластинки и только у немногих более
примитивных групп покрыта эмалевидным
веществом — ганоином. Анальное и
мочеполовое отверстия, как правило, не
объединены в клоаку и отодвинуты от
тазового пояса (за исключением лишь
примитивных хрящевых ганоидных рыб). Очень
характерно строение головного мозга, в
котором у лучеперых рыб, в отличие от
лопастеперых и от всех других
позвоночных, передний мозг имеет тонкую
эпителиальную крышу, лишенную нервного
вещества.
Лучеперые рыбы появились примерно
за 350 миллионов лет до нашей эры и
были сперва представлены преимущественно
более примитивными группами толсточешуйных ганоидных рыб. В течение 100—180
миллионов лет, в пермский и
триасовый периоды, эти группы были
многочисленны и разнообразны. Затем их
численность и распространение сократились,
началось вымирание, и до нашего времени
сохранилось всего около 50 видов этих
некогда многочисленных более архаичных
групп лучеперых рыб. На смену им
начали развиваться собственно костистые
рыбы, появившиеся около 200 миллионов лет
назад, в среднем триасовом периоде. Эта
группа развивалась в течение первых
50 миллионов лет довольно медленно.
Затем, в последующие 50—70 миллионов лет,
эволюция костистых рыб чрезвычайно
ускорилась, и к концу этого времени они
начинают доминировать над ганоидными.
В настоящее время костистые рыбы
достигли замечательного разнообразия и
многочисленности — свыше 19000 видов,
группируемых в большое число отрядов.
Система подкласса лучеперых рыб все
еще недостаточно разработана.
Относительно простые схемы, использовавшиеся до
1940 г., неприемлемы в настоящее время
после обстоятельных морфолого-таксономических и палеонтологических
исследований последних десятилетий.
Работа над новой системой еще не
закончена. В тексте этой книги говорится о
36 отрядах, группируемых в целях
удобства рассмотрения в 10 надотрядов.
Принимается нижеследующая система:
Надотряд I. Ганоидные (Ganoidomorpha), с 4
отрядами: 1) Осетрообразные, 2) Многоперообразные, 3) Амиеобразные, 4) Панцирникообразные.
Надотряд II. Клюпеоидные (Clupeomorpha),
с 6 отрядами: 5) Тарпонообразные, 6) Гоноринхообразные, 7) Сельдеобразные,
8) Лососеобразные, 9) Миктофообразные, 10) Китовидкообразные.
Надотряд Ш. Араваноидные (Osteoglossomorpha),
с 2 отрядами: 11) Араванообразные, 12) Клюворылообразные.
Надотряд IV. Ангвиллоидные (Anguillomorpha),
с 3 отрядами: 13) Угреобразные, 14) Мешкоротообразные, 15) Спиношипообразные.
Надотряд V. Циприноидные (Cyprinomorpha),
с 2 отрядами: 16) Карпообразные, 17) Сомообразные.
Надотряд VI. Атериноидные (Atherinomorpha),
с 3 отрядами: 18) Карпозубообразные,
19) Сарганообразные, 20) Атеринообразные.
Надотряд VII. Параперкоидные (Parapercomorpha), с 2 отрядами: 21) Перкопсообразные,
22) Трескообразные.
Надотряд VIII. Бериксоидные (Beryxomorpha),
с 3 отрядами: 23) Бериксообразные, 24) Солнечникообразные, 25) Опахообразные.
Надотряд IX. Перкоидные (Percomorpha), с
7 отрядами: 26) Колюшкообразные, 27)
Кефалеобразные, 28) Слитножаберникообразные,
29) Окунеобразные, 30) Скорпенообразные,
31) Камбалообразные, 32) Иглобрюхообразные.
Надотряд X. Батрахоидные (Batrachomorpha),
с 4 отрядами: 33) Пегасообразные, 34) Жабообразные, 35) Присоскобрюхообразные,
36) Удилыцикообразные.
Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Часть 2. (Acipenseriformes — Polynemiformes). Г.У.Линдберг, М.И.Легеза 1965 г.
VI. Класс TELEOSTOMI — ВЫСШИЕ РЫБЫ
Внутренний скелет более или менее окостеневший. Кожные кости есть на черепе, на челюстях и на плечевом поясе. Наружное жаберное отверстие одно, прикрытое жаберной крышкой. Palatoquadratum не слито с черепом. (Берг, 1948 : 12, 1949 : 1317). 2 подкласса: Crossopterygii и Actinopterygii, из которых первый в Японском море и в сопредельных водах не представлен.
Подкласс ACTINOPTERYGII
Скелет парных плавников не бисериального типа. Нет внутренних ноздрей. Внутренние ноздри есть у некоторых Uranoscopidae (Astroscopus) (Берг, 1940 : 159, сноска 2). Чешуя не космоидного типа. (Берг, 1948 : 57).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ОТРЯДОВ МОРСКИХ РЫБ ПОДКЛАССА ACTINOPTERYGII КЛАССА TELEOSTOMI, ВСТРЕЧАЮЩИХСЯ В ВОДАХ ЗАПАДНОЙ ЧАСТИ ТИХОГО ОКЕАНА
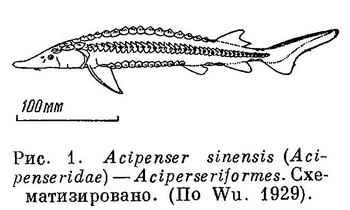
1 (2). Тело покрыто 5 рядами крупных костяных жучек ромбической формы (рис. 1). Тело голое у сем. Polyodontidae, представленного в пресных водах Китая 1 видом — Psephurus gladius (Martens), распространенным в Янцзыцзяне (рис. 2). XII. Acipenseriformes.
2 (1). Тело не покрыто 5 рядами костяных жучек.
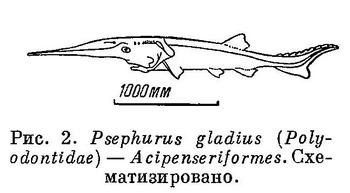
3 (4). Череп асимметричный; оба глаза на одной или правой или левой стороне головы, которая является верхней стороной тела (рис. 3). Спинной и анальный плавники с большим числом лучей. Тело сильно сжатое с боков, высокое, овальной формы. Рыбы лежат на дне на боку. XXXII. Pleuronectiformes.
4 (3). Череп симметричный; глаза расположены по обе стороны головы. Рыбы если лежат на дне, то не на боку, а на брюхе.
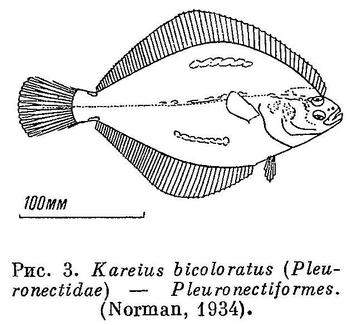
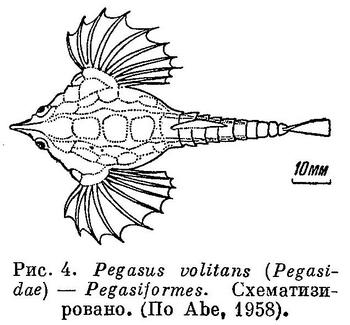
5 (6). Тело сильно приплюснуто сверху вниз и заключено в панцирь из костных пластинок. Грудные плавники с горизонтальным основанием, очень большие, с 10—18 неветвистыми лучами, твердыми в виде колючек у основания и мягкими членистыми на конце; расположены плавники горизонтально, что подчеркивает сильную приплюснутость тела (рис. 4). Спинной плавник 1, короткий; в нем, как и в анальном, по 5 мягких неветвистых лучей. Брюшные плавники заметно позади основания грудных на брюхе; в них по 1 колючему и 1—3 ветвистым лучам. В хвостовом 8 лучей. Рыло удлиненное, часто шилообразное, зазубренное. Рот нижний, беззубый. Подглазничные кости соединены с предкрышкой. Плавательного пузыря нет. XXXVII. Pegasiformes.
6 (5). Тело, голова и плавники не как на рис. 4. У несколько сходных по форме рыб сем. Agonidae грудные плавники с вертикальным или косым, но не горизонтальным основанием.
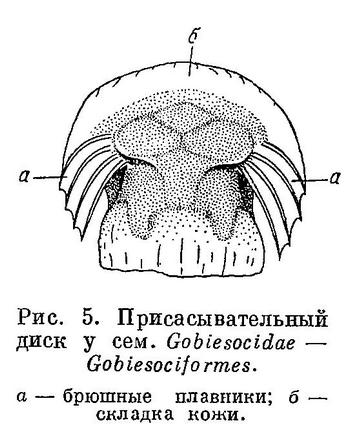
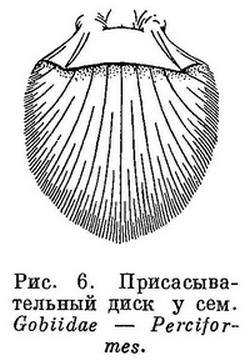
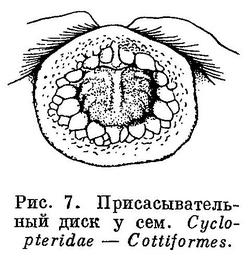
7 (8). На груди имеется присасывательный диск полукруглой формы (рис. 5), образованный широко отставленными друг от друга брюшными плавниками и складкой кожи, поддерживаемой спереди посредством cleithra, сзади — посредством postcleithra; в брюшных плавниках в каждом 1 скрытый колючий и 4 неветвистых луча. Не путать с брюшным диском у Gobioidei и Cyclopteridae из отрядов Perciformes и Cottiformes (рис. 6, 7). Один спинной плавник, расположенный в задней части тела над анальным; в плавниках нет колючих лучей (рис. 8). Кожа голая. Плавательного пузыря нет. XXXV. Gobiesociformes.
8 (7). На груди нет такого присасывательного диска.
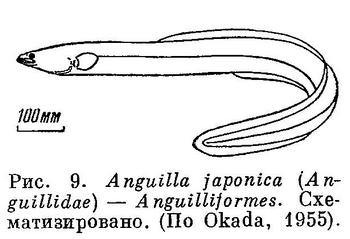
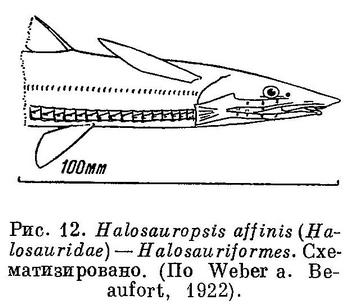
9 (10). Тело угреобразное (рис. 9). Брюшных плавников нет. Плавники без колючек. Спинной и анальный плавники очень длинные и, как правило, соприкасаются с хвостовым или сливаются с ним; иногда хвостовой плавник совершенно отсутствует и конец хвоста голый. Нет отдельных предчелюстных костей; они слиты с мезэтмоидом (а нередко также и с сошником); верхний край рта окаймлен как этими костями, так и верхнечелюстными. (У рыб с угреобразной формой тела из отряда Symbranchiformes (рис. 10), широко распространенных в пресных и солоноватых водах тропического пояса, предчелюстные не слиты с мезэтмоидом и почти или совсем отодвигают верхнечелюстные от верхнего края рта. Жаберные отверстия соединены в 1 непарную продольную щель на горле. Плавательного пузыря нет. У взрослых нет грудных и обычно брюшных плавников, но у личинок 15 мм длины грудные еще имеются; нет лучей в длинных спинном и анальном плавниках, сливающихся с хвотовым. У сем. Alabetidae, распространенного в водах Австралии и Тасмании, имеются расположенные на горле брюшные плавники из 2 лучей, а также лучи в вертикальных плавниках.) Позвонков много, до 260. Плавательный пузырь, если он есть, соединяется с кишечником при помощи протока (открытопузырные). (У рыб с угреобразной формой тела из отрядов Notacanthiformes (рис. 11) и Наlosauriformes (рис. 12) плавательный пузырь не имеет протока (закрытопузырные). Брюшные плавники (из 8—10 лучей) имеются, расположены на брюхе. У Notacanthiformes в отличие от Halosauriformes в анальном плавнике передние лучи, а в брюшных плавниках наружные лучи колючие; впереди спинного плавника имеется ряд изолированных колючих лучей или спинной плавник представлен только одними колючками; фотофор не бывает; предкрышка развита нормально, не зачаточная. Океанические глубоководные рыбы, известные и у берегов Японии. Для южных берегов п-ова Корея, из Мокпхо указывается нахождение Halosaurus affinis Günther из сем. Halosauridae (Mori, 1952 : 67), но мы не вносим этот вид в состав фауны рыб Желтого моря, так как эта глубоководная рыба была поймана несомненно на значительных глубинах Восточно-Китайского моря.) Жаберные щели узкие. Длинные лучи жаберной перепонки поддерживают кожу, прикрывающую большую жаберную полость. XVII. Anguilliformes.

10 (9). Тело не угреобразное, а если напоминает угреобразное, то имеются или брюшные плавники, или колючие лучи в плавниках, или обособленные предчелюстные кости, или неувеличенная жаберная полость.
11 (12). Челюсти, а также гиомандибулярная и квадратная кости очень длинные, далеко простираются назад и в несколько раз превышают длину черепной коробки (рис. 13). Рот огромный, глотка громадная, растяжимая. (Исключительно большой рот по сравнению с другими семействами отряда Perciformes имеют представители сем. Chiasmodontidae, но он все же ничтожно мал по сравнению со ртом и глоткой у Saccopharyngiformes.) Крышечных костей нет. Брюшных и хвостового плавников нет, или последний зачаточный. В плавниках нет колючих лучей. Жаберные отверстия в виде овальных щелей на брюхе. Имеются светящиеся органы в виде борозды, расположенной вдоль основания спинного плавника. Плавательного пузыря, нет. Глубоководные рыбы (рис. 14) [Saccopharyngiformes.]
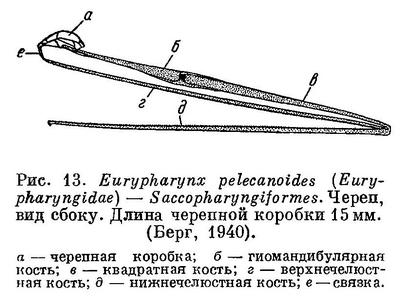
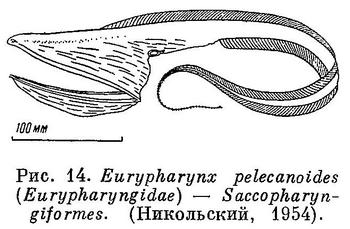
12 (11). Челюсти нормальной длины, а если и увеличены, то выдаются вперед, а не назад. Глотка не громадная и не растяжимая.
13 (14). Первый спинной плавник видоизменен в присасывательный диск, расположенный на голове (рис. 15). XXXIII. Echeneiformes.
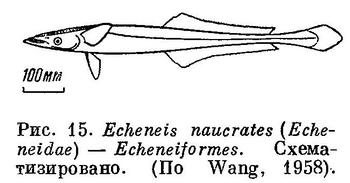
14 (13). Первый спинной плавник не видоизменен в присасывательный диск.
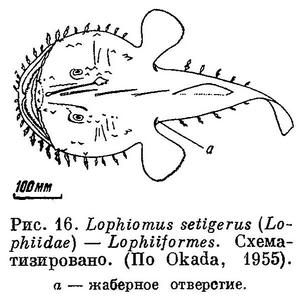
15 (16). Жаберные отверстия расположены под или позади грудных плавников, но не впереди них, малы, овальной формы. Брюшные плавники, если имеются, на горле, с 1 колючим и 5 ветвистыми лучами. В первом спинном плавнике, если имеется, первый луч превращен в удочку (illicium; рис. 16). XXXVI. Lophiiformes.
16 (15). Жаберные отверстия расположены впереди грудных плавников.
17 (18). Рыло сильно удлинено и имеет вид трубки; на конце которой расположен маленький рот. Квадратная и предкрышечная кости сильно удлинены (рис. 17). Брюшные плавники, если имеются, расположены на брюхе или во всяком случае отчетливо за грудными, не имеют колючек, а лишь 3—7 мягких лучей. 1 спинной плавник, также без колючек, а если 2, то первый с колючими лучами. XXII. Syngnathiformes.
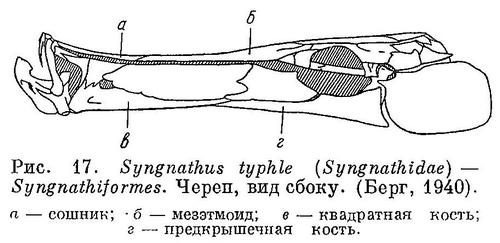
18 (17). Рыло не имеет вида трубки; если же похоже на трубку, то брюшные плавники расположены под или недалеко за грудными и имеют, как правило, сильную колючку.
19 (20). Грудной плавник (рис. 18) имеет 2 основания: верхняя часть плавника прикреплена связками, к 2 верхним радиалиям; нижняя, состоящая из нескольких свободных нитей — к четвертому (нижнему) радиалию; промежуточный третий радиалий не несет плавниковых лучей. (У Rhodichthys regina Collett, относимого в настоящее время к сем. Liparidae (Cottiformes), нижняя лопасть грудного плавника также совершенно отделена от верхней, но в отличие от Polynemiformes у этой рыбы хвостовой плавник сливается со спинным и анальным, а вместо нормальных брюшных плавников, расположенных под грудными или позади их основания, имеются на горле усиковидные лучи нижней лопасти грудного плавника, которые ошибочно могут быть приняты за брюшные плавники (Андрияшев,. 1954 : 467, рис. 273).) Брюшные плавники под грудными, несколько позади их основания. XXVIII. Polynemiformes.
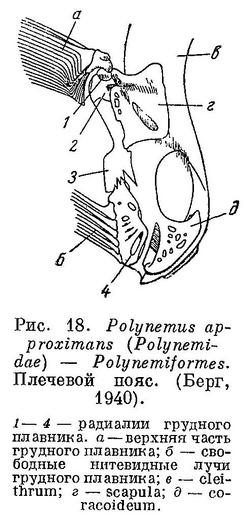
20 (19). Грудной плавник с 1 основанием; все радиалии несут лучи; нижние лучи иногда бывают значительно изменены по сравнению с верхними, но значительного промежутка между основаниями 2 групп лучей не бывает.
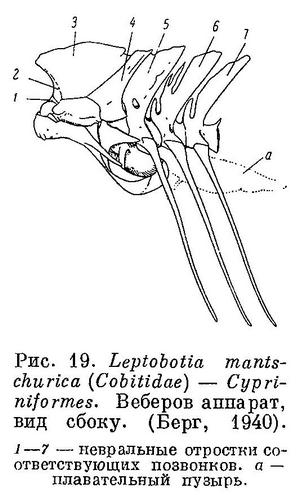
21 (22). Веберов аппарат имеется (рис. 19). (О наличии Веберова аппарата можно судить по изменению передних позвонков
позвоночника, что хорошо видно на рисунках в работе Такахаси (Y. Takahashi, 1962, figs. 20, 20'), в которой приведено описание позвоночников 121 вида костистых рыб.) Брюшные плавники, если они есть, за грудными на брюхе. Плавники без колючек (рис. 20), если же в спинном, анальном и грудном плавниках, или же в некоторых из них имеются колючие лучи, то их обычно не более 1 (в спинном иногда бывает 2 колючки). Иногда бывает жировой плавник. Плавательный пузырь, как правило, соединяется протоком с кишечником (открытопузырные). (В Японском море известно только 3 вида, представители 3 семейств.) XVI. Cypriniformes.
22 (21). Веберова аппарата нет.
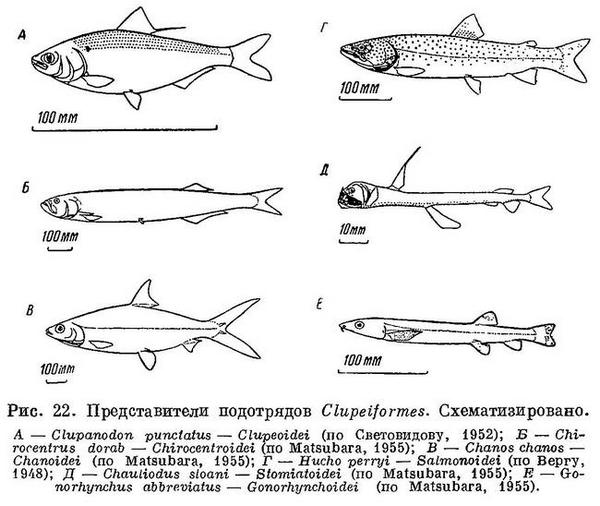
23 (24). Мезокоракоид (рис. 21), как правило, имеется (нет у подотряда Esocoidei — щуковидных, встречающихся в пресных водах, и у семейства Microstomidae из подотряда Salmonoidei — лососевидных). Ротовая щель сверху окаймлена, как правило, предчелюстными и верхнечелюстными костями (только одними предчелюстными у семейства Albulidae и Chanidae). Чешуя, как правило, циклоидная. Плавательный пузырь обычно с протоком (открытопузырные). (У Argentina (Clupeoidei) и у Opisthoproctus (Opisthoproctoidei) плавательный пузырь не соединен с кишечником (Берг, 1940 : 241, 248).) В спинном плавнике нет колючих лучей. Бывает жировой плавник. Брюшные плавники, если имеются, расположены на брюхе далеко за грудными, а если недалеко за ними, то нет жирового плавника и нет фотофор. В Японском море встречаются представители 6 подотрядов (рис. 22). XIII. Clupeiformes.
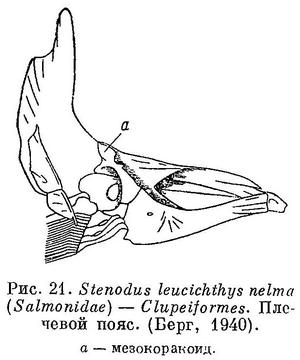
24 (23). Мезокоракоида нет.
25 (26). Жировой плавник обычно имеется. Сходны с подотрядом Stomiatoidei (Clupeiformes), отличаясь от него, кроме отсутствия мезокоракоида, тем, что рот окаймлен одними предчелюстными. Брюшные плавники с 6—13 лучами; расположены они под грудными плавниками (рис. 23), а если позади их, то недалеко, и, кроме того, имеются жировой плавник и фотофоры (рис. 24). Тазовые кости не соединены с ключицей. Плавательного пузыря нет, или он с протоком (открытопузырные). Радиалии грудного плавника окостеневшие. Хвостовой плавник хорошо развит, не сливается с анальным. Бывают светящиеся органы. XIV. Scopeliformes. или XIV. Scopeliformes.

26 (25). Жирового плавника нет
27 (28). Хвостовой плавник редуцирован и сливается, но только с одним длинным анальным плавником (Сходный тип хвостовой части тела имеется у Siluridae (Cypriniformes), у Notopteridae (Clupeiformes) и у Notacanthidae (Notacanthiformes), но у всех у них брюшные плавники находятся на брюхе, а не впереди грудных.); спинной плавник с очень коротким основанием, высокий, расположен почти над основанием грудного плавника. Брюшные плавники впереди грудных и состоят каждый из 1—2 лучей (рис. 25). Близок к Scopeliformes, но радиалии грудного плавника представлены 1 хрящевой пластинкой. XV. Ateleopiformes.
28 (27). Хвостовой плавник, если имеется, хорошо выражен, а если слит, то в равной мере как с анальным, так и с хвостовым плавниками.
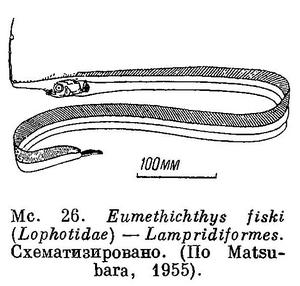
29 (44). Брюшные плавники имеются и прикреплены нормально на брюхе, или недалеко за грудными, или, наконец, под грудными, но заметно позади вертикали заднего края основания грудных плавников; иногда они бывают очень малы и широко разобщены друг от друга. Как исключение брюшные плавники расположены под основанием грудных у сем. Veliferidae (рис. 30). Брюшных плавников, как исключение, нет у рыб рода Eumethichthys сем. Lophotidae (Lampridiformes), но для них характерен выступающий вперед, иногда очень далеко, лоб; на переднем, конце лба начинается длинный спинной плавник, доходящий до хвостового плавника; анальный плавник с коротким основанием очень маленький, расположен недалеко от заднего конца лентообразного тела (рис. 26) или отсутствует. Брюшные плавники отсутствуют также у Trachypterus ishikawai из сем. Trachypteridae (Lampridiformes), но они могут быть мизерными и поэтому незаметными; тело у них также лентообразное (рис. 27).


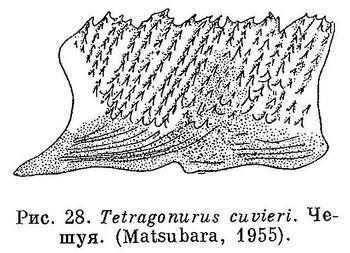
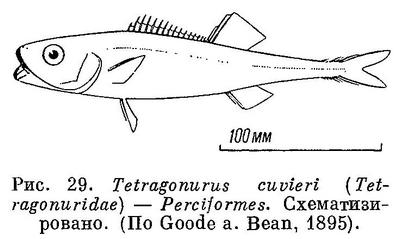
30 (31). Имеется своеобразная ромбическая чешуя с килями; расположена она косыми поперечными рядами. Чешуи каждого ряда соединены друг с другом (рис. 28). Передняя часть спинного плавника колючая (рис. 29). XXIX. Perciformes (частью: Tetragonuridae).
31 (30). Нет такой чешуи.
32 (37). Колючих лучей нет ни в одном плавнике.
33 (34). Тело высокое, сжатое с боков. В брюшных плавниках
8—9 или 15—17 лучей. Если в брюшных плавниках 8—9 мягких лучей, то лучи спинного и анального плавников спереди очень длинные, а кзади постепенно укорачивающиеся, не имеют вида типичных колючих лучей, хотя они неветвистые и даже твердые; только первые 1—2 сильно укороченных луча спинного и анального плавников могут быть приняты за колючие лучи. Хвостовой стебель короткий, довольно высокий, не круглый в сечении. Хвостовой плавник с выемкой. Тело высокое, сжатое с боков; скелет хорошо окостеневший. Имеется орбитосфеноид. XXIII. Lampridiformes (частью: Veliferidae (рис. 30) и Lampridae (рис. 31)).

34 (33). Тело низкое. В брюшных плавниках 6—7 лучей.
35 (36). Боковая линия имеется, расположена вдоль нижнего края тела. Хвостовой плавник глубоко выемчатый. Нижнеглоточные кости полностью слиты. В брюшных плавниках 6 ветвистых лучей (рис. 37 и 38). У рода Dermogenys Van Hasselt, 1823 (Hemirhamphidae, Beloniformes из вод Филиппинских островов и южнее) в анальном плавнике имеется 3 слабых, колючих луча несколько отделенных от плавника (Fowler, 1933, 1934 : 326, fig. 80). XVIII. Beloniformes.
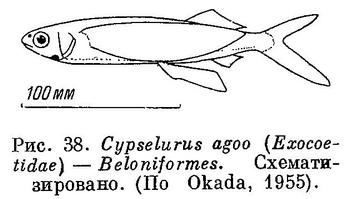
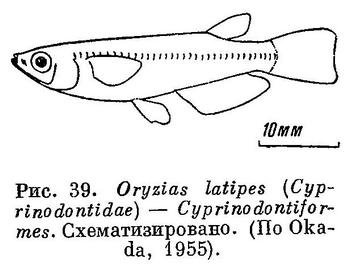
36 (35). Боковой линии нет. Если имеются сейсмосензорные поры, то они не располагаются вдоль нижнего края тела. Хвостовой плавник закруглен или обрезан, но глубоко не вырезан. Нижнеглоточные
кости не слиты. (Рис. 39). XXIV. Cyprinodontiformes.
37 (32). Колючие лучи имеются хотя бы в одном из плавников.
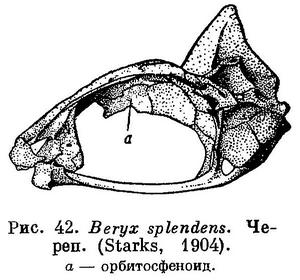
38 (39). Брюшные плавники без колючих лучей. Колючие лучи имеются в первом спинном и анальном плавниках. В брюшных плавниках 7—8 мягких лучей. Есть гиоидные усики. (У рыб сем. Mullidae (Perciformes) также имеются усики, но не гиоидные; они расположены сразу же за симфизисом нижней челюсти (рис. 33); кроме того, у Mullidae спинной плавник с 2 основаниями и нет орбитосфеноида. У рыб сем. Ophidiidae за гиоидные усики могут быть ошибочно приняты брюшные плавники. У этих рыб нет колючих лучей) Есть орбитосфеноид (рис. 32). XXV. Beryciformes (частью: Polymixiidae).
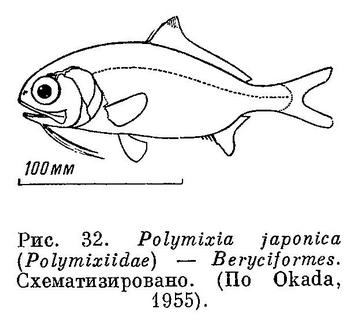
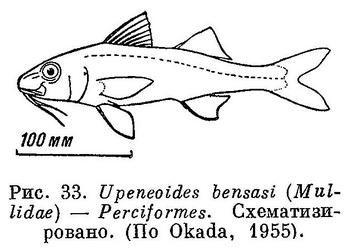
39 (38). Брюшные плавники с колючими лучами.
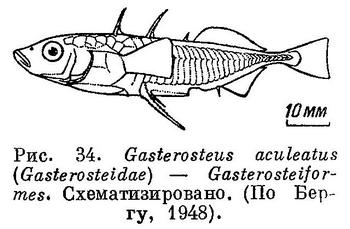

40 (41). В брюшных плавниках, расположенных очень недалеко за основанием грудных, имеется обычно очень сильный, колючий луч и 0—4 мягких луча (рис. 34). (Сильная колючка в брюшных плавниках с числом ветвистых лучей менее 5 имеется у рыб сем. Monocentridae (Beryciformes), у которых все тело одето в броню из крупных костных пластинок и у которых есть орбитосфеноид (рис. 35). Сильная колючка имеется и у рыб сем. Triacanthodidae и Triacanthidae (Tetrodontiformes), у которых тазовые кости прикреплены прямо к ключице и имеется очень маленький рот и сросшиеся или соединенные верхнечелюстные и предчелюстные кости (рис. 36).) Впереди спинного плавника не менее 2 свободных колючек, не соединенных между собой перепонкой. Тело веретенообразное, голое или с рядом пластинок вдоль боков тела. Тазовые кости не причленены прямо к ключице. Орбитосфеноида нет.XXI. Gasterosteiformes.


41 (40). В брюшных плавниках колючий луч не такой сильный, как у Gasterosteiformes, и имеется во всяком случае не менее 5 ветвистых лучей.
42 (43). Брюшные плавники расположены на брюхе или под грудными, но заметно позади их основания (рис. 40). (Брюшные плавники, расположенные на брюхе, имеются, как исключение, в отряде Zeiformes у Parazen pacificus (Zeidae), но у этого вида спинные плавники соприкасаются основаниями, а брюшные плавники не имеют колючего луча (рис. 49).) Тазовые кости соединены с грудным поясом (с cleithra или с postcleithra) посредством связки. Спинные плавники далеко отставлены один от другого. XXVII. Mugiliformes.
43 (42). Брюшные плавники расположены не на брюхе, а под грудными, но не очень далеко позади их основания. Тазовые кости соединены с грудным поясом путем непосредственного прикрепления к ключице (cleithrum). Спинные плавники соприкасаются основаниями, а если разобщены, то у рыб или 1 пара ноздрей (Notothenioidae) или имеются усики на симфизисе (Mullidae). XXIX. Perciformes (частью: Percoidei).
44 (29). Нет брюшных плавников, а если они имеются, то прикреплены они под основаниями грудных плавников, или впереди них на горле, или даже на подбородке.
45 (52). Спинной и анальный плавники без колючих лучей и у рыб из рассматриваемого района не слиты с хвостовым; если же в спинном бывает 1 колючка, то хвостовая часть тела сильно удлинена, а хвостовой плавник мал и трудно различим. Тазовые кости прикреплены к ключице (cleithrum) с помощью связок. Рыбы без колючих лучей в плавниках встречаются и в других отрядах, но у них тазовые кости прикреплены прямо к ключице.
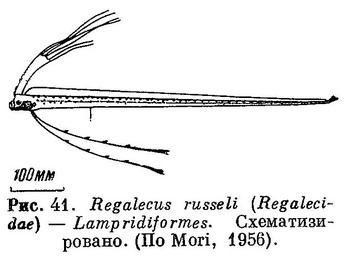
46 (47). Тело сильно удлиненное, часто лентообразное, иногда с изогнутым кверху хвостовым плавником, нередко с удлиненными лучами спинного плавника на голове. Брюшные плавники отсутствуют, а если имеются, то без колючего луча (рис. 41). Есть орбитосфеноид (рис. 42) XXIII. Lampridiformes (частью: Regalecidae).
47 (46). Тело не сильно удлинено, а если удлинено, то нет орбитосфеноида.
48 (51). Колючего луча в брюшных плавниках нет.
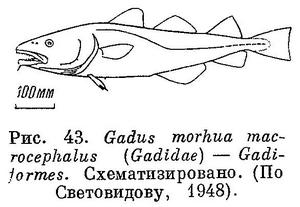
49 (50). Брюшные плавники расположены впереди грудных. Хвостовой плавник обособленный, симметричный (рис. 43). Хвостовой плавник слит со спинным и анальным у сем. Muraenolepidae, распространенного в водах южного полушария. XIX. Gadiformes.
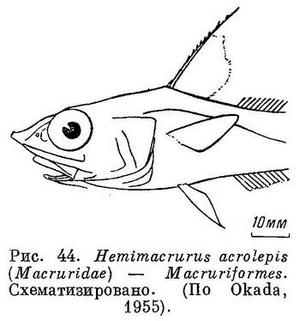
50 (49). Брюшные плавники расположены под грудными, а если не сколько впереди, то хвостовой плавник необособлен. (У рода Lyconodus, Gilchrist, 1922 (Macruridae, Macruriformes) брюшные плавники расположены заметно позади основания грудных.) Хвостового стебля нет. Хвостовой плавник слабо развит, сливается со спинным и анальным плавниками, имеющими длинные основания. В спинном плавнике иногда имеется 1 колючка (рис. 44). XX. Macruriformes (частью: Macruridae).
51 (48). Колючий луч в брюшных плавниках имеется; он пластинчатовидный. Кроме него имеется 5 ветвистых лучей. Нет боковой линии.
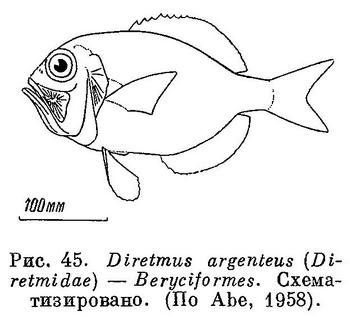
Есть орбитосфеноид (рис. 45). XXV. Beryciformes (частью: Diretmidae).
52 (45). Спинной и анальный плавники с колючими лучами.
53 (56). Тазовые кости не прямо прикреплены к ключице (cleithrum), а с помощью связки.
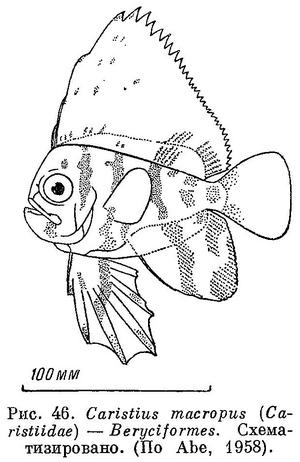
54 (55). В спинном и анальном плавниках не более чем по 2 колючих луча. Спинной плавник 1; начинается он над глазами, очень высокий, больше высоты тела. Брюшные плавники (I 5) также с длинными лучами, превосходящими длину головы (рис. 46). Есть орбитосфеноид. XXV. Beryciformes (частью: Caristiidae).
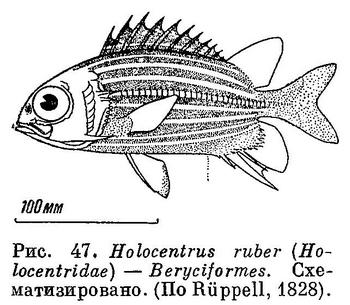
55 (54). В спинном и анальном плавниках значительно больше чем по 2 колючих луча: в спинном 7—13 (обычно 10—13) сильных колючих лучей, в анальном 4 сильных колючих луча. Спинных плавников 2: они невысокие, соприкасаются, хотя хорошо различаются; основание первого значительно больше основания второго спинного плавника (рис. 47). Есть орбитосфеноид. XXV. Beryciformes (частью: Holocentridae).
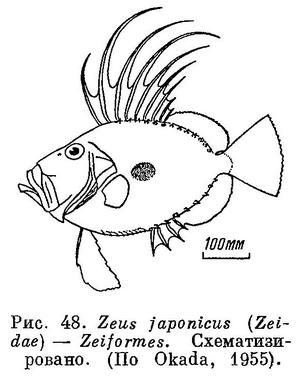
56 (53). Тазовые кости прямо прикреплены к ключице (cleithrum), без помощи связок.
57 (58). Анальный плавник в передней части обычно имеет 1—4 сильных колючих луча, которые отставлены несколько вперед и выделяются в виде самостоятельного плавничка (рис. 48); реже не выделяется (рис. 49). (У рода Parazen Kamohara, 1935 ни в брюшном (в котором 7 ветвистых лучей), ни в анальном плавниках нет сильных колючих лучей и брюшные плавники расположены на брюхе (рис. 49).) Брюшные, как правило, под грудными или впереди них; 1 колючий и 6—9 ветвистых лучей. Хвостовой I+10—13+I. Задневисочная не вильчатая. XXVI. Zeiformes.
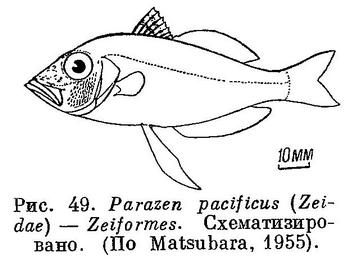
58 (57). Анальный плавник не имеет в передней части самостоятельной, отделенной колючей части.
59 (60). Очень сильная колючка в брюшных плавниках и 3 ветвистых луча. Тело одето в броню. Имеется орбитосфеноид. Рот и жаберные отверстия большие (рис. 35). XXV. Beryciformes (частью: Monocentridae).
60 (59). Нет такой колючки или, если есть, тело не одето в броню.
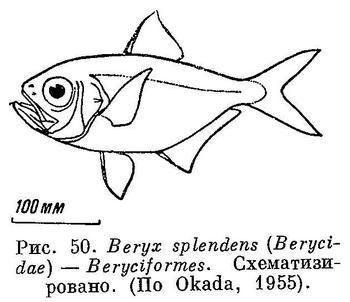
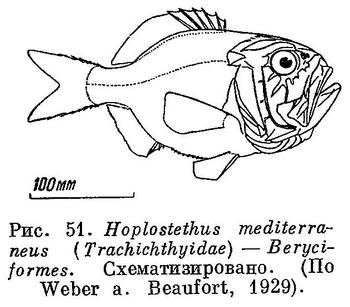
61 (62). В брюшных плавниках 1 колючий луч и более 6 (7—13) ветвистых лучей, а если 6 ветвистых лучей, то в анальном 1—2 колючих луча и брюшко обычно со срединным рядом килеватых или зазубренных чешуй. Имеется орбитосфеноид. XXV. Beryciformes (частью: Berycidae (рис. 50) и Trachichthyidae (рис. 51).
62 (61). В брюшных плавниках, если они имеются, 1 колючий и не более 6 ветвистых лучей.
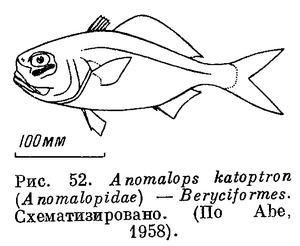
63 (64). Имеется светящийся орган под глазом (рис. 52). Спинной плавник с 2 основаниями. В первом спинном 4—5 колючих лучей; в анальном плавнике 2 колючих луча. Имеется орбитосфеноид. XXV. Beryciformes (частью: Anomalopidae).
64 (63). Нет светящегося органа под глазом.
65 (68). Подглазничная опора имеется — подглазничные кости своим задним отростком прочно соединены с предкрышкой; она хорошо прощупывается под кожей щеки (рис. 53).
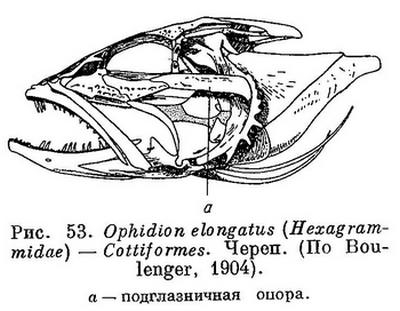
66 (67). Носовые кости не слиты вместе. Позади верхнезатылочной и теменных костей нет 2 пар tabularia. С предкрышкой соединена вторая подглазничная. XXX. Cottiformes.
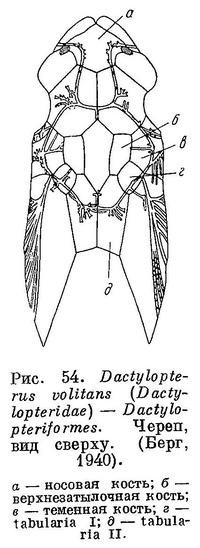
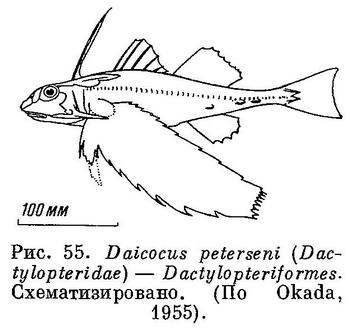
67 (66). Носовые кости слиты вместе, образуя 1 непарную кость. Позади верхнезатылочной и теменных костей имеется 2 пары tabularia, из которых задняя пара очень крупная (рис. 54). С предкрышкой соединена первая подглазничная. Грудные плавники очень большие (рис. 55). XXXI. Dactylopteriformes.
68 (65). Подглазничной опоры нет, подглазничные кости не соединены с предкрышкой.
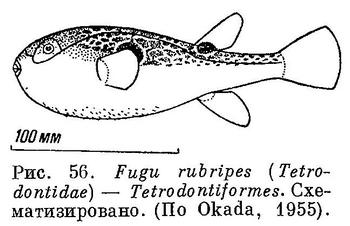
69 (70). Рот очень мал; верхнечелюстные кости плотно соединены с предчелюстными, иногда даже слиты с ними. Жаберные отверстия небольшие (рис. 56). Задневисочная, если имеется, не вильчатая, соединена швом с pteroticum. XXXIV. Tetrodontiformes.
70 (69). Рот не такой маленький; верхнечелюстные не слиты и не соединены с предчелюстными. Жаберные отверстия обычно большие. Задневисочная обычно вильчатая. XXIX. Perciformes.
Рыбы северных морей СССР. А.П.Андрияшев 1954 г.
Класс TELEOSTOMI — ВЫСШИЕ РЫБЫ
Берг, Система рыб, 1940 : 149.
Есть настоящие челюсти. Полукружных каналов 3. Парные плавники (если не утрачены вторично) есть. Внутренний скелет более или менее окостеневший. Кожные кости есть на черепе, челюстях и плечевом поясе Наружное жаберное отверстие одно, прикрыто жаберной крышкой которая, как правило, поддерживается специальным скелетом. Palatoquadratum не слито с черепом, который обычно гиостилического типа. Нет клоаки. Никогда не бывает птеригоподиев. Известны с девона.
В пределах северных морей СССР представители 25 отрядов подкласса Actinopterygii.
Рыбы пресных вод СССР и сопредельных стран. Л.С.Берг 1948 г.
Подкласс ACTINOPTERYGII
Скелет парных плавников не бисериального типа. Нет внутренних, ноздрей. Чешуя не космоидного типа.