Merlangius merlangus (Linnaeus, 1758)
Мерланг, атлантическо-средиземноморский мерланг, исландский мерланг, норвежский мерланг, черноморский мерланг, мерланка (Rus),
Whiting
(Eng)
Синонимы:
Gadus euxinus Nordmann, 1840
Gadus merlangus Linnaeus, 1758
Merlangus linnei Malm, 1877
Merlangus vulgaris Fleming, 1828
Odontogadus merlangus (Linnaeus, 1758) Атлантическо-средиземноморский мерланг (Rus),
Odontogadus merlangus euxinus (Nordmann, 1840) Черноморский мерланг, мерланка (Rus),
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
Мерланг черноморский — Odontogadus merlangus euxinus (Nordmann, 1840)
Два анальных плавника сближены, почти слиты основаниями. Боковая линия слабо изогнута. Усика на подбородке нет, или он очень маленький. D1 14—17. D2 16—19. D3 18—22. A1 28—32. A2 19—22. Окраска тела ровная серо-оливковая, более темная на спине, более светлая к брюшку. Длина тела в среднем 28 см. Обитает в прибрежной зоне Черного моря.
Ихтиопланктон Черного моря. Т.В.Дехник 1973 г.
ЧЕРНОМОРСКИЙ МЕРЛАНГ — ODONTOGADUS MERLANGUS EUXINUS (NORDMANN)
Условия размножения. Данные по размножению мерланга детально проанализированы и обобщены В. Д. Бурдак (1964) в монографии по биологии этого вида.
Сопоставление результатов наблюдений различных исследователей (Водяницкий, 1930а, 1936; Пчелина, 1936, 1940; Косякина, 1938; Виноградов, 1948, 1949) и анализ собственных материалов позволили В. Д. Бурдак прийти к заключению, что в Черном море мерланг нерестится круглый год. Круглогодичность нереста мерланга В. Д. Бурдак объясняет тем, что в Черном море пригодные для размножения температурные условия он находит в течение всего года. Зимой он нерестится во всем верхнем 80-метровом слое, температура которого составляет в это время 7—8°. Летом нерест происходит в постоянно охлажденных слоях, где температура воды колеблется от 6 до 8° (Виноградов, 1948; Водяницкий и Казанова, 1954; Бурдак, 1955, 1964).
Массовый нерест мерланга в Черном море происходит в холодное время года, преимущественно с декабря по март. Икринки и личинки встречаются в планктоне при температуре воды 4,0—16,0° (Зайцев, 1959а; Георгиев и др., 1960). Нерестится мерланг на всем пространстве Черного моря в пределах континентального плато при солености как обычной для Черного моря — 17—18‰ (Бурдак, 1964), так и при пониженной (14,75‰) — в районе Одесского залива (Зайцев, 1959а).
В пределах своего ареала мерланг нерестится повсюду, где он встречается (Ehrenbaum, 1905—1909). В Северном море нерестовый период также растянут. Нерест начинается в конце января, иногда в конце декабря, и продолжается до июля. Разгар нереста в южной части Северного моря наблюдается в марте и апреле (Ehrenbaum, 1905—1909; Aurich, 1942). В Средиземном море размножение этого вида не установлено (D'Ancona, 1933). Сроки нереста смещаются от летних месяцев в северных районах ареала к зимним — в южных. Эти изменения хорошо показаны В. Д. Бурдак (1964), которая обобщила данные различных исследователей. На основании этих данных В. Д. Бурдак пришла к заключению, что в разных районах ареала в период массового нереста средняя температура воды изменяется в очень небольших пределах — от 6 до 9—10°. По наблюдениям Аурих (1942), оптимальная температура для нереста мерланга лежит между 5—8°, по Дама (Damas, 1909) — между 6—10°, согласно Шмидта (1909), — изменяется от 5 до 10° и по Расселу (Russel, 1935) — от 9,0 до 9,5°.
Соленость в пределах нерестового ареала этого вида колеблется довольно значительно. Аурих (1942) отмечает, что распределение икринок мерланга в южной части Северного моря ограничено в основном изогалиной 34,75‰ и что во время нереста мерланг избегает вод с соленостью меньше 32‰. В Черном море, как показано выше, развивающиеся икринки и личинки мерланга встречаются при солености 14,75—18,0‰.
Нерест мерланга повсеместно происходит в пределах континентального плато, на глубине не более 150—200 м, преимущесгвенно от 25 до 100 м (Ehrenbaum, 1905—1909; Damas, 1909; Schmidt, 1909; Knudsen, 1950). Наиболее благоприятной глубиной для нереста Аурих (1942) считает 20—70 м.
Строение икринок. Икринки мерланга пелагические, сферической формы, без жировой капли. Желток гомогенный, оболочка гладкая, совершенно прозрачная, перивителлиновое пространство очень узкое (рис. 9).
Размеры живых развивающихся икринок по данным различных авторов колеблются от 1,10 до 1,33 мм (Водяницкий и Казанова, 1954; Зайцев, 1959а; Георгиев и др., 1960; наши данные). Средний размер икринок по Ю. П. Зайцеву 1,20 мм, по нашим измерениям — 1,27 мм. Размеры икринок мерланга в Северном море близки к приведенным для Черного моря — 0,97—1,32 мм. Их средний размер уменьшается от февраля к июню от 1,213 до 1,043 мм (Ehrenbaum, 1905—1909). По форме, величине и строению икринки мерланга очень сходны с икринками глоссы (Platichthys flesus luscus), встречающимися в планктоне, как и икринки мерланга, преимущественно в холодное время года. Отличительные признаки, проявляющиеся по мере развития эмбрионов обоих видов, приведены ниже при описании развития глоссы.
Эмбриональное развитие. Опыты по развитию икры и личинок мерланга были поставлены в январе и феврале 1964 г. Для опытов были использованы икринки, собранные в аквариуме, где мерланг регулярно нерестился. Обычно ежедневно или через сутки в аквариуме появлялись свежевыметанные икринки мерланга. В 8—9 ч утра икринки находились на начальных стадиях развития — с обозначающимся протоплазматическим куполом или с намечающейся первой бороздой дробления. Образование протоплазматического бугорка на анимальном полюсе происходит примерно через 1,5—2 ч после оплодотворения. Следовательно, в условиях аквариума мерланг нерестится в утренние часы. Плазменный бугорок занимает небольшую часть поверхности желтка, высота его составляет около 10% диаметра желтка.
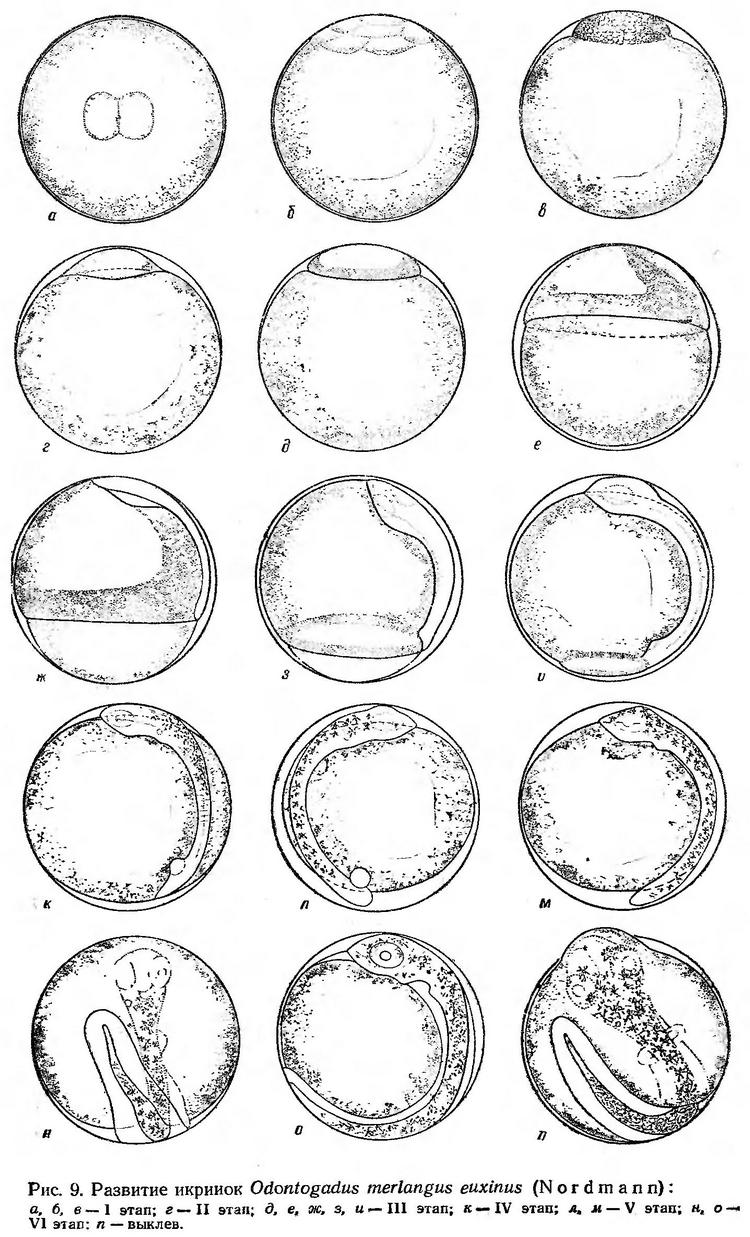
Процесс дробления идет медленно. В течение 3 ч при температуре 7,2° образуется 8 бластомеров. Через 5 ч после появления первой борозды насчитывается 32 бластомера. При температуре 7,2—8,0° процесс дробления продолжался около 27 ч (рис. 9, а, б, табл. 11). Высота бластодиска на стадии мелкоклеточной морулы равна 0,16—0,17 мм, длина основания 0,60—0,62 мм (рис. 9, в). В последующие 19—20 ч происходит смещение бластомеров в поверхностный слой, образуется бластоцель. Зародышевый диск чуть приподнимается, края его несколько приспущены. Высота зародышевого диска 0,17—0,20 мм, длина основания 0,67—0,70 мм (рис. 9, г). Вслед за обрастанием начинается подворачивание краев бластодиска и через 3—4 ч образуется зародышевое утолщение (рис. 9, д). Через 9—10 ч после начала обрастания бластодиск охватывает примерно 1/3 поверхности желтка (рис. 9, е). По мере сползания бластодиска формируется зародышевая полоска. Когда края бластодиска достигают экватора, начинается дифференцировка зародышевых органов — закладывается нервная трубка, намечаются зачатки глаз (рис. 9, ж, з). Ко времени замыкания желточной пробки обособляются глазные бокалы, образуются мозговые доли, появляется 7—8 туловищных сегментов. Головной конец зародыша достигает анимального полюса (рис. 9, и). Процесс обрастания и гаструляции при температуре 7,2° продолжался 35—37 ч (рис. 9, д, и).
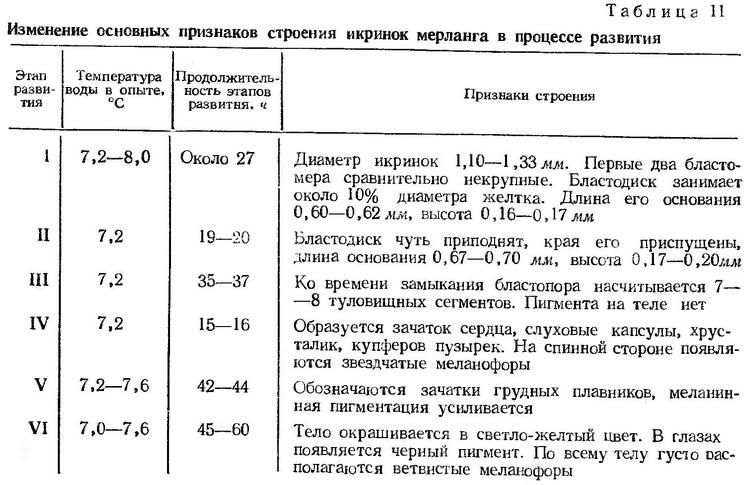
К моменту закрытия бластопора (на IV этапе развития) на спинной стороне тела появляются первые меланофоры. Намечаются зачаток сердца, слуховые капсулы, хрусталики, кишечник, образуется купферов пузырек (рис. 9, к). Звездчатые меланофоры расположены на голове и на спинной стороне в передней части туловища. Через 15—16 ч после замыкания бластопора появляется хвостовая почка (рис. 9, л). Черный пигмент рассеян по всему телу, купферов пузырек отшнуровывается и затем исчезает. Вместе с ростом хвостового отдела происходит нарастание в длину нервней трубки, формируются хвостовые миотомы. На V этапе четко обозначаются мозговые доли, слуховые капсулы, хрусталики, появляются зачатки грудных плавников. Когда эмбрион охватывает около 2/3 поверхности желтка, начинается биение сердца. Первоначально пульсация очень слабая — 32—34 удара в минуту. При температуре воды 7,2 — 7,6° V этап продолжался 42 — 44 ч (рис. 9, л, м).
Одновременно с пульсацией сердца начинаются первые слабые мышечные подергивания эмбриона. По мере удлинения хвостового отдела движения становятся более энергичными. Частота биения сердца возрастает до 65—70 ударов в минуту. На VI этапе значительно увеличиваются глаза, мозговые доли, зачатки грудных плавников. Широкая плавниковая кайма начинается на спинной стороне на уровне заднего края глаза, опоясывает хвост и заканчивается у заднего края желточного мешка. В заглазннчной области появляется два крупных черных пигментных пятна. Голова и передняя часть тела эмбриона окрашиваются в бледно-желтый цвет, все тело интенсивно пигментируется ветвистыми меланофорами (рис. 9, н). В пигментации эмбрионов наблюдаются существенные различия. Некоторые эмбрионы перед выклевом приобретают интенсивно черную окраску благодаря крупным паутинообразным меланофорам, густо расположенным на светло-желтом фоне на спинной стороне и на боковых поверхностях. Другие эмбрионы из одного и того же опыта пигментированы значительно слабее. На верхней и боковых поверхностях расположены сравнительно мелкие звездчатые меланофоры, так что четко проступает фоновая желтая окраска. Указанные различия проявляются в дальнейшем и у личинок.
Незадолго перед выклевом начинается пигментация глаз (рис. 9, о). Пульсация сердца учащается до 90—94 сокращений в минуту. Хвост эмбриона почти смыкается с головой. Голова упирается в оболочку и постепенно ее выпячивает (рис. 9, п). Под воздействием фермента вылупления оболочка утончается и разрывается. Через разрыв выталкивается голова, а затем с помощью сильных мускульных движений освобождаются от оболочки туловище и хвост. Эмбриональный период развития мерланга при температуре 7,2—8,0° продолжался около 8 суток (табл. 11).
Постэмбриональное развитие. Живые, только что выклюнувшиеся личинки имеют длину тела 2,7—3,2 мм (Водяницкий и Казанова, 1954; Зайцев, 1959а; наши данные). Желточный мешок большой, эллипсоидальной формы. Голова плотно прижата к нему. Анальное отверстие расположено сразу за желточным мешком и открывается сбоку плавниковой складки. Антеанальное расстояние составляет около 50% всей длины тела. Тело личинки окрашено в светло-желтый цвет и покрыто тонкими паутинообразными меланофорами. Голова и передняя часть туловища обычно окрашены более интенсивно. Желтый пигмент распространяется на плавниковую кайму и на желточный мешок. В глазах имеется сероватый пигмент (рис. 10, а). Личинки плавают в горизонтальном положении брюшной стороной вверх. Очень пассивны. Лишь изредка совершают короткие перемещения и снова переходят в состояние покоя. Концентрируются преимущественно в приповерхностном слое воды.
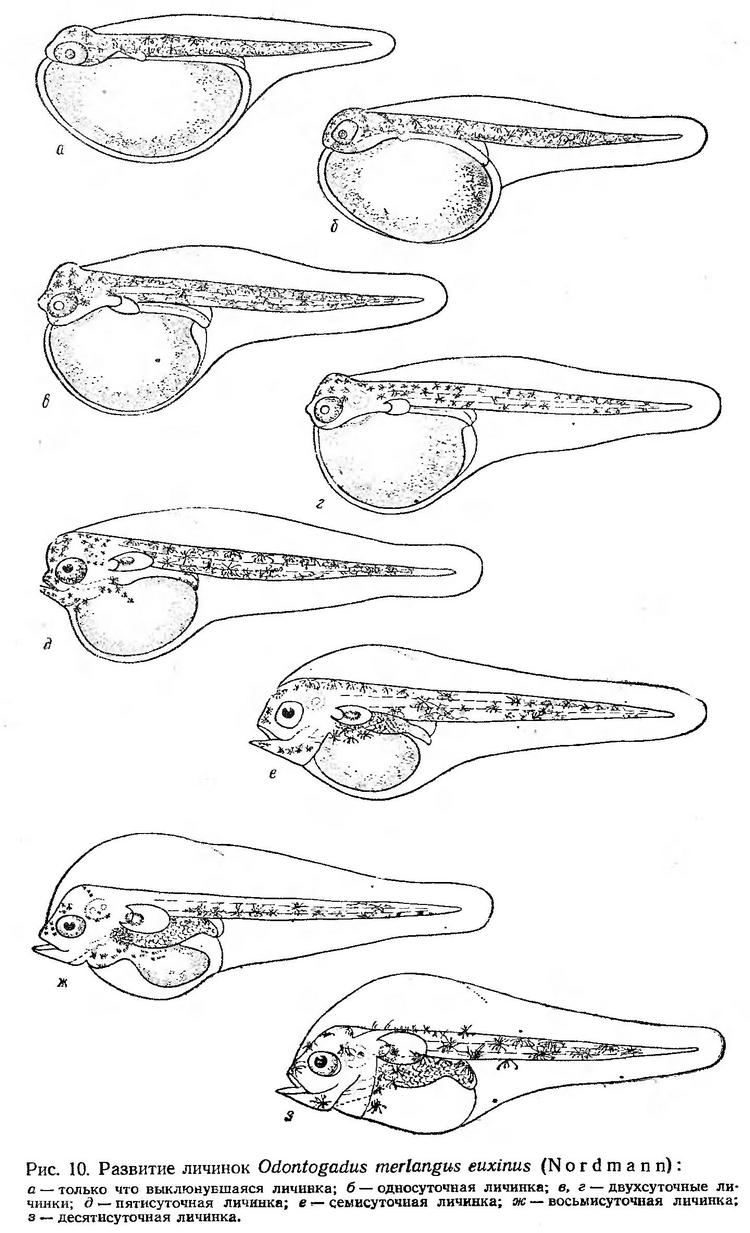
Через сутки длина личинок увеличивается в среднем до 3,4 мм. Антеанальное расстояние относительно длины тела сокращается за счет удлинения хвостового отдела. Желтый пигмент распространяется на желточный мешок. По мере развития тело делается более прогонистым, относительная длина антеанального расстояния уменьшается (рис. 10, б). У личинок в возрасте двух суток передняя часть головы освобождается от желтка, намечается ротовая ямка. Желточный мешок начинает рассасываться и по мере его уменьшения передняя часть спинной плавниковой каймы заполняется жидкостью, так что образуется большая вздутая полость.
У двухсуточных личинок усиливается пигментация глаз. Желтый пигмент распространяется на плавниковую кайму. Все тело личинок покрыто паутинообразными (рис. 10, в) или звездчатыми компактными (рис. 10, г) меланофорами, располагающимися преимущественно на спинной стороне и боковых поверхностях. Личинки становятся более активными, стремительно убегают при приближении какого-либо предмета (препаровальной иглы или пипетки).
У трехсуточных личинок хорошо обозначена ротовая ямка, появляется плавательный пузырь, усиливается желтый пигмент, он покрывает все тело; желточный мешок распространяется на плавниковую кайму.
В возрасте пяти суток (рис. 10, д) личинки достигают наибольшей длины и имеют самое короткое антеанальное расстояние. На более поздних этапах развития происходит укорачивание длины тела, пропорции меняются снова в сторону увеличения антеанального расстояния и возрастания отношения высоты тела к длине. Спинная полость продолжает заполняться водой. У пятисуточных личинок глаза интенсивно пигментированы, хотя предметное видение, очевидно, не выражено. Личинки принимают нормальное горизонтальное положение (плавают спиной вверх), отмечается слабое движение рта. Они еще мало подвижны — периоды покоя значительно больше времени разовых перемещений. Изменяется характер пигментации. Желтый пигмент в значительной степени редуцируется; меланофоры перемещаются на вентральную сторону. Это перемещение связано с изменением положения тела. Как и у многих окунеобразных, пигментные клетки расположены на стороне тела, обращенной к дну, а не к поверхности (Крыжановский, 1953). Такое расположение пигментных клеток С. Г. Крыжановский (1959) рассматривает как защитное приспособление. Прозрачные личинки с темным нижним контуром мало заметны сверху на фоне дна.
У семисуточных личинок (рис. 10, е) большие вертикально поставленные грудные плавники приобретают подвижность. Личинки начинают плавать в разных направлениях. Желточный мешок еще сравнительно большой. Челюсти оформлены, движение рта становится более ритмичным.
Восьмисуточные личинки мерланга (рис. 10, ж) еще имеют сравнительно большой остаток желточного мешка. Желтый пигмент исчезает. Движения личинок стремительные, несмотря на наличие большой оводненной полости.
В возрасте 10 суток у личинок сохраняется небольшой остаток желточного мешка (рис. 10, з). Плавательный пузырь заполняется воздухом. Личинки активно двигаются в разных направлениях, держатся преимущественно у дна аквариума. В возрасте 11—12 суток личинки в условиях аквариума погибают.
Распределение и численность икринок и личинок. Икринки и личинки мерланга встречаются вдоль всего северного, восточного и западного побережья Черного моря (Водяницкий, 1930а; Косякина, 1938; Зайцев, 1959а; Георгиев и др., 1960; Бурдак, 1964). Сведений о размножении этого вида у побережья Турции нет.
По наблюдениям В. А. Водяницкого (1930а), в Новороссийской бухте икринки мерланга ловились в самые холодные месяцы — с января по апрель — только в поверхностных слоях воды в небольшом количестве. Е. Г. Косякина (1938) отмечает нахождение икринок этого вида обычно с ноября по май в поверхностных слоях, а также в сентябре на глубине 140 м.
Мальки длиной 9,2—28 мм, по данным 3. М. Пчелиной (1940), встречались в Новороссийской бухте единичными экземплярами в марте и апреле как в поверхностных слоях, так и глубже лежащих. Автор отмечает также нахождение малька мерланга 11 мм длиной в открытом море на глубине 30 м в конце июня. На нахождение мальков мерланга в открытом море в феврале указывает В. Н. Никитин (1929). У болгарского побережья икринки и личинки мерланга, по данным Ж. М. Георгиева и др. (1960), встречаются в открытом море (в 10—15 милях от берега) всегда единичными экземплярами. В указанных работах, к сожалению, не приводятся данные о численности икринок и личинок этого вида в планктоне. Не содержится также подобных данных в обзорной работе В. Д. Бурдак (1964), хотя дана карта распределения их по всему северному и восточному побережью Черного моря.
По наблюдениям Ю. П. Зайцева (1959а), в Одесском заливе наибольшее отмеченное количество икринок мерланга составило 1 экз/м2 поверхности моря. Наши данные также свидетельствуют о малой численности икринок мерланга в плактоне. В холодное время года (январь, 1959 г.) численность икринок колебалась от 2 до 14 экз/м2 (в среднем 6 экз.), в летнее время (июль 1957 г.) — от 2 до 6 экз/м2 (в среднем 4 экз.). Количество личинок в оба периода наблюдений не превышало 6 экз/м2.
Приведенные данные указывают на сравнительно низкую интенсивность нереста мерланга в Черном море. Это подтверждается сопоставлением численности икры и личинок мерланга в Черном море с соответствующими данными в центре его ареала — в Северном море. Согласно наблюдениям Ауриха (1942), в разгар нереста в южной части Северного моря (март 1926 и 1937 гг.) численность икринок в планктоне достигала 330—375 экз/м2, а численность личинок — 75 экз/м2. В проливе Скагеррак автором была отмечена очень высокая численность икринок — 702 экз/м2.
Сравнительно небольшое количество икринок и личинок мерланга, отмеченное в планктоне Черного моря, на первый взгляд, не соответствует выводам В. Д. Бурдак (1964) о многочисленности популяций этого вида в Черном море. Однако особенности биологии размножения мерланга (круглогодичный нерест, многопорционный характер икрометания, неодновременность созревания нерестовых популяций) свидетельствуют о непрерывном поступлении икринок и личинок в планктон и, следовательно, о высокой их среднегодовой продукции.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Мерланг (Odontogadus merlangus)
распространен у атлантических и
средиземноморских берегов Европы, с особым
подвидом в Черном море. Атлантическо-средиземноморский мерланг достигает
обычно длины 30—50 см, но бывает и до
55—68 см. Спина и верх головы у него
буровато-коричневые, бока с
пятнышками, сетчатым или волнистым узором.
Первый анальный плавник длинный (30—35 лучей), хвостовой плавник без выемки.
Усика на подбородке нет или он очень
маленький.
У берегов Европы мерланг доходит на
север до Исландии и юго-западной части
Баренцева моря. Держится на небольшой
глубине, заходит в предустьевые
пространства. Нерестует на глубине 20—80 м
при температуре воды 5—10° С. Личинки
и мальки долго держатся под колоколом
больших медуз (Cyanea). Питается
ракообразными и мелкой рыбой — песчанкой,
сельдью и др. Промысел мерланга дает
до 1/4 миллиона тонн в год, больше
всего добывают его в Северном и
Ирландском морях.
Черноморский мерланг, или мерланка
(Odontogadus merlangus euxinus), отличается от
атлантического меньшей величиной (обычно до
25 см, очень редко до 45—50 см),
желтовато-серой окраской, наличием
маленького усика на подбородке, более
длинными парными плавниками.
Живет в холодных слоях Черного моря,
на глубине 50—60 м. Изредка заходит в
опресненные зоны северо-западной части
Черного моря.
У берегов Крыма мерланка появляется
почти всегда после сильных сгонных
ветров, при которых происходит подъем
глубинных холодных вод. Половой зрелости
достигает в возрасте 2—3 лет.
Икрометание происходит в течение большей части
года в пределах всего ареала, наиболее
интенсивно зимой.
Молодь длиной 3—7 см встречается и в
отдалении от берегов над большими
глубинами под куполом медуз-ризостом
(Rhizostoma pulmo).
Промыслового значения не имеет,
служит пищей хищным рыбам — катрану,
белуге, камбале-калкану, а также
дельфинам.
Рыбы северных морей СССР. А.П.Андрияшев 1954 г.
1. Odontogadus merlangus (Linne) — Мерланг (рис. 78).
Gadus merlangus Linne, Syst. Nat., ed. X, 1758 : 258 (Атлантич. океан у берегов Европы). — Smitt, Scand. Fish., I, 1893 : 487, pl. XXIV, fig. 1 (синонимия, описание, распространение, биология). — Scott, Ann. Rep. Fish. Board Scotl., XX, pt. 3, 1902 : 513 (питание). — Collett, Christ. Vidensk.-Selsk. Forhandl., 9, 1903 : 44 (на север до Варангер-фиорда; дл. до 552 мм; плодовитость). — Ehrenbaum, Eier und Larven, 1905—1909 : 231, fig. 86 (развитие). — Дерюгин, Фауна Кольск. зал., 1915 : 588 (по данным Эксп. научн.-пром. иссл. Мурмана — в Кольск. зал., редок). — Книпович, Определитель, 1926 : 154, фиг. 118. — Saemundsson, Synops. Fish. Iceland, 1927 : 26 (вокруг Исландии; биология). — Ehrenbaum, Handb. Seefischer. N. Europas, II, 1936 : 111, fig. 83 (биология, уловы). — Есипов, Промысл. рыбы Баренц. м., 1937 : 109 (уловы). — Sund in: Andersson, Fiskar Fiske Norden, I, 1942 : 178, pl. 50 (биология, цветн. фотогр.). — Mаслов, Тр. Полярн. инст. морск. рыбн. хоз. океаногр., VIII, 1944 : 168 (не редок в Кольск. и Мотовск. заливах; дл. до 45 см).
Gadus merlangus merlangus Световидов, Докл. АН СССР, I, 6, 1935 : 421 фиг. 1 (сравнение с черноморским подвидом).
Odontogadus merlangus merlangus Световидов, 1948 : 146, фиг. 23, табл. VIII, XLVI, LXX (литература, измерения, сводка по биологии).

I D 12—15, II D 18—25, III D 19—22, I А 30—35, II А 21—23, Р 19—20, позвонков 53—57.
Усика на подбородке нет или он рудиментарный. Верхняя челюсть немного выступает вперед по отношению к нижней. Длина основания первого анального плавника лишь немного менее антеанального расстояния и составляет 30—34% L. Верхняя часть головы и тела буровато-коричневая с серовато-фиолетовым оттенком. Бока сероватые с желтоватыми пятнышками, которые часто соединены в сетчатый узор или образуют узкие продольные волнистые полоски. Боковая линия темнее окраски боков. Низ головы и брюхо белое. У берегов Европы длина обычно до 30—35 см, наибольшая — до 55 см. В исландских водах достигает больших размеров: обычно 40—50 см, наибольшая — до 68 см (Сэмундсон).
Распространение. Атлантический океан у берегов Европы; Средиземное море. На север доходит до юго-западной части Баренцова моря (Кольский залив, губа Дальне-Зеленецкая); молодь обычна вокруг Исландии, но взрослые преимущественно у южной стороны острова. Наиболее обычен у берегов Великобритании и Ирландии, в Северном море и юго-западных берегов Скандинавского полуострова. Западная часть Балтийского моря, единично до Готланда. В Черном море (возможно также в Эгейском и Мраморном морях) заменяется подвидом Odontogadus merlangus euxinus (Nordm.), который отличается сравнительно хорошо развитым усиком и более длинными грудными и брюшными плавниками.
Биология. Держится на небольших глубинах, часто и вблизи берегов, обычно небольшими стаями, как у дна, так и в пелагиали. Заходит в предустьевые пространства и даже в низовья рек. Нерестует повсюду, кроме крайне восточных и северных районов своего ареала, на небольших глубинах (20—80 м) при температуре 5—10°. Период нереста растянут, в северных районах приходится на весенне-летнее время (апрель—июль). Плодовитость 109—285 тыс. икринок диаметром около миллиметра (1.0—1.3). Выклюнувшиеся личинки, имеющие в длину 3.2—3.5 мм, долгое время держатся под колоколом крупных медуз (Cyanea). К осени мальки достигают 10—13 см в длину, а еще через 2 года — 24 см. Исландский мерланг растет значительно быстрее, достигая половозрелости в 3—4-годовалом возрасте при длине 30—40 см. Питается ракообразными, а также мелкими рыбами (песчанка, молодь сельди и камбал).
Хозяйственного значения в наших водах не имеет. Уловы западноевропейских стран достигают 700—800 тыс. ц в год (главным образом в Северном море).
Фауна СССР. Рыбы. Том IX, вып.4. Трескообразные. А.Н.Световидов 1948 г.
*la. Odontogadus merlangus merlangus (Linne )— Мерланг (табл. VIII, XLVI, LXX)
— Gadus merlangus Linne, Syst. nat., ed. X, 1758 : 258 (Oceano Europeo).
— Merlangus vulgaris Kröyer, Danm. Fiske, II, 1843—1845 : 83.
— Gadus merlangus Steindachner, Sitzungsber. Akad. Wiss. Wien, math.-naturw. CI., VLII, 1, 1868 : 703 (ex parte: Копенгаген, Берген, Виго, Адриатическое море).
— Merlangus linnei Malm, Göteb. Bohusl. Fauna, 1877 : 485.
— Gadus euxinus (non Nordm.) Ninni, Atti Soc. Ital. Sci. Nat., XIII, 1880 : 75 (Адриатическое море); Graeffe, Arb. Zool. Inst. Univers. Wien. u. Zool. Stat. Triest, VII, 1880 : 10 (Адриатическое море).
— Gadus merlangus Williamson, 26th. Ann. Rept. Fish. Board Scotl., pt. 3, (1907), 1909 : 97 (описание черепа).
— Gadus merlangus merlangus Световидов, Докл. Акад. Наук, I, 6, 1935 : 431, ф. 1 (сравнение с черноморской формой).
Распространение, биология. Mobius u. Неinсkе, Fische Ostsee, 1883 : 235 (западная часть Балтийского моря, на восток до о. Готланда); Smitt, Scand. fish., I, 1893 : 487 (распространение, биология); Fulton, Publ. Circonst., 8—9, 1904 : 4; Hjort, Rapp. Proces-Verb., X, 1909 : 56, f. 15 (места нереста); Damas, ibid. : 34 (биология); J. Schmidt, ibid. : 57, 142 (биология); Дерюгин, Фауна Кольского залива, 1915 : 588 (Кольский залив); Saemundsson, Medd. Komm. Havunders., Fisk., VIII, 1925 : 26 (возраст и рост); Bull, Journ. Mar. Biol. Assoc., XV, 1928 : 207 (зрелость, химический состав); Desbrosses, Rev. Trav. Off. Peches Marit., XIII, 1—4, 1939—1943 : 177 (атлантические берега Франции; размножение, развитие, рост); Маслов, Тр. Полярн. научно-исслед. инст. морск. рыбн. хоз. и океаногр., 8, 1944 : 168 (Кольский зал. в 1936 и 1937 гг., Мотовский зал.).
I D 12—15, II D 18—25, III D 19—22, I А 30—35, II А 21—23, sp. br. 21—22, vert. 54 (По J. Schmidt'e, 53—57 позвонков.) Усика на подбородке обычно нет, а если есть, то едва заметный. Из 14 исследованных экземпляров едва заметный рудиментарный усик имели 6 (3 экз. длиной 92, 107 и 265 мм из Северного моря, 1 экз. длиной 202 мм с Мурманского побережья и 2 экз. 207 и 236 мм длиной из Адриатического моря). В процентах к длине тела: антеанальное расстояние 34.1—38.2, антедорсальное расстояние 28.6—30.0, длина грудного плавника 13.8—15.6, длина брюшного плавника 7.4—8.6, длина основания 1-го спинного плавника 11.6—14.5, длина основания 2-го спинного 19.8—22.1, длина основания 1-го анального 30.4—33.1, высота 1-го анального 30.4—33.1, высота 1-го спинного 9.4—12.4, высота хвостового стебля 4.7—5.5, длина его 9.6—11.7, длина головы 23.9—25.2. В процентах к длине головы: длина рыла 32.2—35.6, продольный диаметр глаза 21.6—28.8, длина верхней челюсти 40.6—44.1, длина нижней челюсти 50.7—56.0, ширина лба 18.3—20.7. Окраска крайне разнообразная, у крупных спина и верхняя часть головы желтовато-коричневая с серовато-фиолетовым оттенком, бока желтовато-серые, вдоль и под боковой линией желтые пятнышки, частично слитые в зигзагообразные или сетевидные полосы; брюшко и нижняя часть головы молочно-белые. Непарные плавники приблизительно одной окраски с верхней частью тела, спинные несколько темнее анальных. Грудные плавники желтоватые, с черным пятном у основания сверху; брюшные плавники молочно-белые. Обычные размеры 30—35 см, реже до 68 см.

Распространение. У европейских берегов Атлантического океана и в прилегающих морях. На север до берегов Мурмана (Дальне-Зеленецкая губа, Екатерининская гавань) и Исландии; у берегов Норвегии и Фарерских о-вов, в Северном море, в западной части Балтийского моря единично до о. Готланда, вокруг Великобритании и Ирландии, единично кругом Пиренейского п-ова, в Средиземном и Адриатическом морях. В Эгейском и Мраморном морях типичная форма или Odontogadus merlangus euxinus. Heuglin (Reisen nach d. Nordpolarmeer in d. Jahren 1870 u. 1871, III, 1872—74 : 213), повидимому, ошибочно указывает у о. Вайгача.

Биология. Держится обычно вблизи берегов, не только у дна, но часто в промежуточных и поверхностных слоях воды. Заходит в опресненные предустьевые пространства и даже в низовья рек. Нерест происходит почти по всему ареалу, за исключением самых северных пределов. Основные нерестилища расположены вдоль берегов южной части Северного моря, вдоль восточных и северо-западных берегов Великобритании, вокруг Ирландии, у южных и юго-западных берегов Исландии, вдоль берегов Норвегии на север до 65° с. ш. Менее интенсивно нерестится в центральной и северной части Северного моря, у Фарерских о-вов, в Скагерраке и в западной части Балтийского моря, в Ламанше и в северной части Бискайского залива. По времени нереста это одна из самых позднонерестующих форм тресковых; нерест начинается с конца января и продолжается до середины июля, в северных частях ареала несколько позже, чем в южных. Есть указания на возможный нерест осенью (в октябре, Fulton, l. c.). Нерест происходит на глубинах, начиная с 20 м, обычно на 40—80 м, при температуре от 5 до 10° С в Северном море и 9.5—11.5° С у берегов Ламанша и Бретани, в воде нормальной океанической солености, от 30 до 35.25‰. Молодь очень долго ведет пелагический образ жизки. Миграции взрослых хорошо не изучены, но, повидимому, больших миграций не совершает, держится более или менее локально, не собирается и в большие стаи. Питается ракообразными и мелкой рыбой (мелкой сельдью, и др.). Рост у берегов Исландии происходит следующим образом (Saemundsson, l. c.):
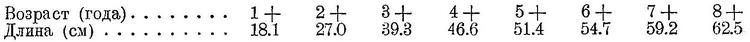
У берегов Норвегии достигает меньшего возраста и растет несколько медленнее. Самки достигают большего возраста и растут, повидимому, немного быстрее самцов. Исландский мерланг в среднем несколько крупнее норвежского; здесь наичаще встречаются особи 40—55 см.
Хозяйственного значения в наших водах не имеет. У западноевропейских берегов ежегодно вылавливается около 700 тыс. ц,. главные места вылова — Северное море, Скагеррак, Каттегат, у берегов Великобритании и Ирландии.

*1b. Odontogadus merlangus euxinus (Nordmann) — Черноморский мерланг, пикша (неправ.), мерланка (у батумских рыбаков по сообщению И. И. Пузанова) (табл. VIII, XLVII).
— Gadus euxinus Nordmann, Faune pontiquo, 1830 : 526, pl. XXVI, f. 2 (повсюду вдоль берегов Черного моря).
— Gadus merlangus (non Linne) Steindachner, Sitzungsber. Akad. Wiss. Wien, math.-naturw. CI., LVII, 1, 1868 : 703 (ex parte : Черное море).
— Gadus euxinus Кесслер, Рыбы Арало-касп.-понт, обл., 1977 : 233 (ex parte : Черное море у берегов Крыма).
— Gadus merlangus euxinus Световидов, Докл. Акад. Наук, I, 6, 1935 : 426, ф. 2 (сравнение с основной формой).
Распространение, биология. Киселевич, Сборн. студ. биол. кружка Новоросс. унив., 3, 1908 : 11 (Черное море у Днестровского лимана, Одесский порт); Зернов, Зап. Акад. Наук, (VIII), XXXII, 1913 : 174 (Черное море у Севастополя); Дренски, Спис. Българск. Акад. Наук., XXV, 1928 : 108 (ex parte: Черное море у Варны, Бургаса, Созополя); Водяницкий, Тр. Севастоп. биол. ст., V, 1936 : 7 (Севастопольская бухта, икринки); Пчелина, Тр. Новоросс. биол. ст., II, 1, 1936 : 32 (мальки.); Косякина, Тр. Новоросс. биол. ст., II, 2, 1938 : 26 (икрометание в районе Новороссийска); Малятский, Зоологич. журн., XVII, 4, 1938 : 667, 674 (роль в питании белуги); Пчелина, Тр. Новоросс. биол. ст., II, 3, 1940 : 71 (мальки в Новороссийской бухте).
I D 14—17, II D 16—19, III D 18—22, I А 28—32, II А 19—22, sp. br. 20—23, vert. 53. На подбородке всегда хорошо заметный усик (У рыб длиной 12—18 см длина усика 2.3—3.2 мм.). В процентах к длине тела: антеанальное расстояние 35.6—38.2, антедорсальное расстояние 27.6—30.2, длина грудного плавника 15.4—18.2, длина брюшного плавника 8.1—11.3, длина основания 1-го спинного плавника 12.3—15.8, длина основания 2-го спинного 17.3—19.9, длина основания 1-го анального 29.0—31.6, высота 1-го спинного 11.7—13.8, высота хвостового стебля 3.6—4.5, длина его 10.5—12.5, длина головы 23.7—25.8. В процентах к длине головы: длина рыла 32.2—34.8, продольный диаметр глаза 22.8—26.5, длина верхней челюсти 38.8—43.3, длина нижней челюсти 49.0—52.2, ширина лба 18.1—20.1. Верхняя часть головы и тела буроватые или желтовато-серые с фиолетовым и розоватым оттенком. Нижняя часть тела белая с серебристым оттенком. Плавники, в особенности спинные и хвостовой, желтоватые с желто-серой полосой спереди и посередине вдоль плавника. 1-й анальный плавник молочно-белый с желтоватым оттенком. Размеры 15—20 см, изредка до 50 см и свыше.
Распространение. Вдоль берегов Черного моря, Керченский пролив и в прилегающих к нему ближайших частях Азовского моря (Казантип) (По любезному сообщению В. Абрамова.).
Биология изучена мало. Редок. У Севастополя в большом количестве появляется осенью, когда температура воды спадает до 16°, и держится в октябре, ноябре, декабре. Половые продукты развиты в ноябре, декабре, апреле. Икрометание против Новороссийска с ноября по май, в глубинных слоях открытого моря (40 м) икра ловится в сентябре; в 1936 г. в глубинных слоях моря значительное количество икринок было обнаружено в мае. В Севастопольской бухте икринки были находимы в мае—июле. Мальки длиной 9.2—28 мм у поверхности и на глубине 30 м при температуре от 8 до 12° С ловились в Новороссийской бухте и у входа в нее в марте—апреле, один малек 11 мм длиной пойман в открытом море на глубине 30 м в конце июня.
Хозяйственного значения не имеет, но играет большую роль в питании белуги, являясь в зимнее время против Севастополя основной ее пищей.
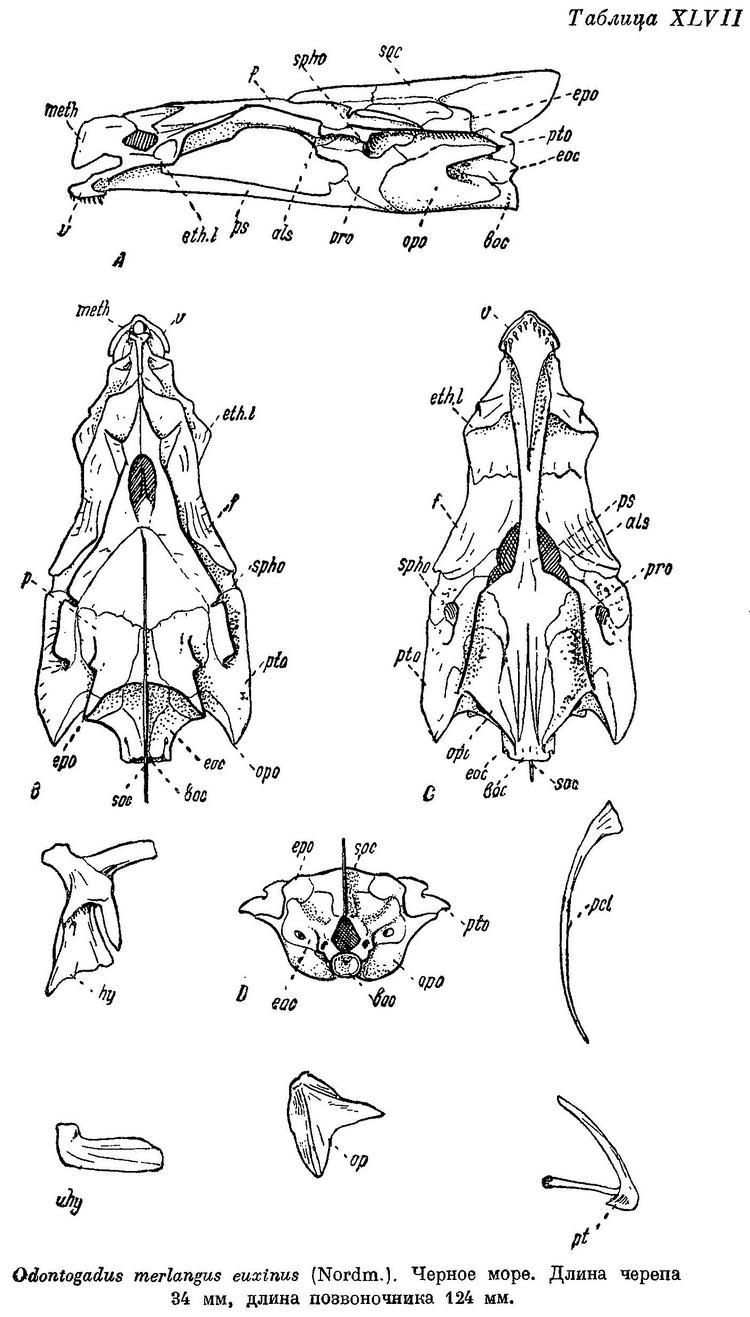
Сравнительные заметки. Главным отличием этой формы, описанной первоначально как самостоятельный вид, от основной являлось наличие усика на подбородке у первой и отсутствие его у второй. Благодаря этому ареал черноморской формы некоторые авторы (Ninni, l. c.; Graeffe, l. с.) считали значительно шире — до Адриатического моря включительно, где были найдены экземпляры, имеющие усики, другие же (Steindachner, l. с.) считали эту форму идентичной с основной после того, как были обнаружены экземпляры с усиками и среди последней. Сравнение экземпляров из Баренцева, Северного, Адриатического и Черного морей (Световидов, l. с.) показало, что черноморские экземпляры существенно отличаются от всех остальных, помимо постоянного наличия усика, рядом и других признаков, причем все эти признаки, как видно из диагноза обеих форм, являются трансгрессивными, вследствие чего черноморский мерланг рассматривается как подвид. Экземпляры (2) из Адриатического моря по всем признакам относятся к типичной форме. Сравнение скелетов обеих форм подтверждает их близость (см. табл. XLVI и XLVII), небольшие различия обнаружены лишь в том, что у Odontogadus merlangus merlangus череп несколько шире и передний конец frontalia выдается несколько более вперед, чем у Odontogadus merlangus euxinus, причем на другом исследованном черепе Odontogadus merlangus euxinus frontalia спереди простираются вперед так же, как у Odontogadus merlangus merlangus. Возможно, что в Адриатическом море окажется форма, промежуточная между ними.