Ctenolabrus rupestris (Linnaeus, 1758)
Гребенчатый губан (Rus),
Goldsinny-wrasse
(Eng)
Синонимы:
Labrus rupestris Linnaeus, 1758
Иллюстрированная энциклопедия рыб. Ст.Франк 1983 г.
В Средиземном море, Атлантическом океане и в Балтийском море живет Ctenolabrus rupestris, откладывающий пелагическую икру. Основная окраска серо-зеленая с темным пятном на верхней части основания хвостового плавника. Размеры до 15 см. Иногда содержится в аквариумах.
Ихтиопланктон Черного моря. Т.В.Дехник 1973 г.
ГРЕБЕНЧАТЫЙ ГУБАН — CTENOLABRUS RUPESTRIS (LINNE)
Условия размножения. Нерест гребенчатого губана в Черном море начинается в середине — конце мая и продолжается до конца августа (Водяницкий, 1936; Косякина, 1938; Дехник и Павловская, 1950; Водяницкий и Казанова, 1954; Дука, 1959; Зайцев, 1959а; Овен,1959; Георгиев и др., 1960).
Температура воды в местах нахождения икринок колеблется в довольно значительных пределах. По нашим наблюдениям (Дехник и Павловская, 1950), икринки встречались при температуре 20,0—23,4°. Ю. П. Зайцев (1959а) отмечает нахождение икринок при температуре воды 12—22°. По данным Л. А. Дука (1959), нерест губана начинается при температуре 12,5°; в период массового нереста температура колебалась от 14,0 до 19,5°.
Развивающиеся икринки гребенчатого губана были найдены в различных районах Черного моря при солености 15,55—18,21‰ (Дехник и Павловская, 1950). В Одесском заливе соленость в местах нахождения икринок колебалась в пределах — 15,93—18,04‰ (Зайцев, 1959а).
Интересные экспериментальные наблюдения Л. С. Овен (1962а) показали, что гребенчатый губан выметывает икру в опытных условиях регулярно через 1—2 суток.
В Средиземном море нерест гребенчатого губана начинается в апреле и продолжается до конца августа (Sparta, 1956а).
В Северном море, как и в Черном, икрометание происходит с мая по июль — август. В Английском канале нерест начинается уже в апреле (Ehrenbaum, 1905—1909, 1936; Hefford, 1910).
Судя по времени размножения этого вида в разных районах ареала можно сделать заключение, что температурные условия в период нереста колеблются в сравнительно небольших пределах. В то же время колебания солености, при которой происходит нормальное развитие икры и личинок, значительные — 15,5—38‰.
Строение икринок. Икринки гребенчатого губана можно легко отличить от личинок других видов. Они сферической формы, сравнительно небольшие, без жировой капли. Желток гомогенный, совершенно прозрачный, оболочка тонкая, гладкая, с фиолетовым или розоватым отливом. Перивителлиновое пространство очень узкое. Диаметр икринок губана в Черном море, по данным различных исследователей и нашим измерениям, колеблется от 0,69 до 1,00 мм (Дехник и Павловская, 1950; Водяницкий и Казанова, 1954; Зайцев, 1959а; Овен, 1959; Георгиев и др., 1960).
Размеры икринок этого вида из Северного моря близки к приведенным для Черного — 0,72—0,94 мм, у южного побережья Англии — до 1,01 мм (Ehrenbaum, 1905—1909). Средний размер икринок в районе Плимута от мая к июлю уменьшается от 0,877 до 0,822 мм (Hefford, 1910).
В Средиземном море икринки Ctenolabrus rupestris несколько мельче — 0,70—0,83 мм (Holt, цит. по Sparta, 1956а).
Эмбриональное развитие прослежено на икринках, собранных в море в ночное время (23—24 ч). Выловленные в это время икринки находились на этапе дробления. Температура воды в опытах колебалась от 19 до 23°.
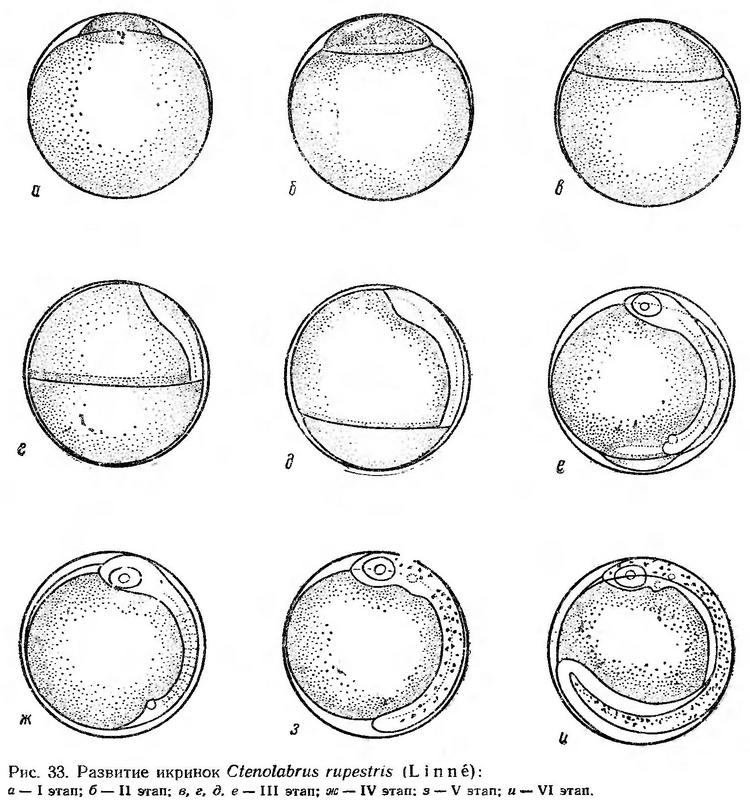
Диск дробления занимает незначительную часть поверхности желтка. Высота бластодиска на стадии морулы мелких клеток равна 0,08—0,10 мм, длина основания — 0,43—0,45 мм (рис. 33, а).
Образование эпителиальной бластулы при температуре 22,6° происходит в течение 1,5—2 ч. Перемещение бластомеров к поверхностному слою сопровождается значительным увеличением бластодиска. Длина его основания достигает 0,75—0,78 мм, высота — 0,20—0,21 мм (рис. 33, б). Вскоре после начала обрастания желтка бластодиском образуется зародышевое утолщение (рис. 33, в). Когда края бластодиска достигают экватора, намечается закладка нервного валика (рис. 33, г). Через 6 ч после начала гаструляции зародышевая полоска приобретает четкие очертания. Хорошо выражен головной конец. Появляются первые туловищные сегменты, намечаются глазные бокалы (рис. 33, д). Бластодиск охватывает 2/3 поверхности желтка. III этап при температуре воды 19—22,8° продолжается 8,5—10 ч. К моменту замыкания бластопора насчитывается 10—12 туловищных сегментов. В глазах намечаются хрусталики. Закладывается хорда. Появляется купферов пузырек (рис. 33, е). После замыкания бластопора продолжается сегментация туловища, закладываются кишечник, сердце. Формируются слуховые капсулы (рис. 33, ж). IV этап при температуре воды 21,6—22° продолжается около 2 ч.
По мере отделения хвоста от желточного мешка происходит сегментация хвостового отдела. Развивается кровеносная система, кишечник, формируются мозговые доли. На спинной стороне зародыша, обращенной книзу, появляется мелкий точечный серовато-черный пигмент (рис. 33, з). При температуре воды 21,2—21,6° V этап продолжается около 4 ч.
По мере развития эмбриона и роста хвостового отдела подергивания учащаются и усиливаются. Эмбрион начинает энергично двигаться. Когда хвост достигает 2/3 поверхности желтка, эмбрион готов к выклеву. Сердце пульсирует 80—95 раз в минуту. Появляются зачатки грудных плавников. Широкая плавниковая кайма охватывает тело (рис. 33, и).
Выклев эмбриона начинается через 6—7 ч после появления первых слабых подергиваний тела и начала пульсации сердца. Процесс эмбрионального развития от начала II этапа при указанной выше температуре воды продолжается 22—25 ч.
Постэмбриональное развитие. Только что выклюнувшиеся личинки имеют длину 2,1—2,6 мм. Они характеризуются стройным, прозрачным телом. Желточный мешок большой, овальной формы, чуть выступает за голову. Анус открывается на значительном расстоянии от желточного мешка, так что преанальная плавниковая кайма хорошо выражена. Антеанальное расстояние составляет 56—58% длины тела. Грудные плавники зачаточные. На спинной стороне тела разбросаны мелкие точечные меланофоры (рис.34, а). Личинки плавают в горизонтальном перевернутом положении или под углом к поверхности, головой вверх. Периоды покоя сравнительно короткие — 5—10 сек. При движении личинки сохраняют горизонтальное положение или чуть поворачиваются на бок. Движения осуществляются с помощью быстрых изгибов тела. Моменты движения продолжаются не более 1—3 сек. За это время личинки проплывают 2—4 мм.
В течение первых суток развития личинок происходят заметные изменения в их строении. Желточный мешок значительно сокращается, в связи с чем передняя часть головы освобождается от него и сразу же намечается ротовая ямка. Антеанальное расстояние относительно длины тела уменьшается до 48—50% за счет удлинения хвостового отдела. Увеличиваются зачатки грудных плавников. Начинается пигментация глаз по периферии. Меланофоры приобретают паутинообразную форму и распространяются со спины на боковые поверхности. В задней части хвостового отдела появляются пигментные клетки на вентральной стороне (рис. 34, б). Личинки по-прежнему очень подвижны. Периоды покоя составляют не более 5—10 сек. Движения принимают характер стремительных бросков, при которых личинки, не меняя положения в воде, «проскакивают» расстояние в 3—7 мм.
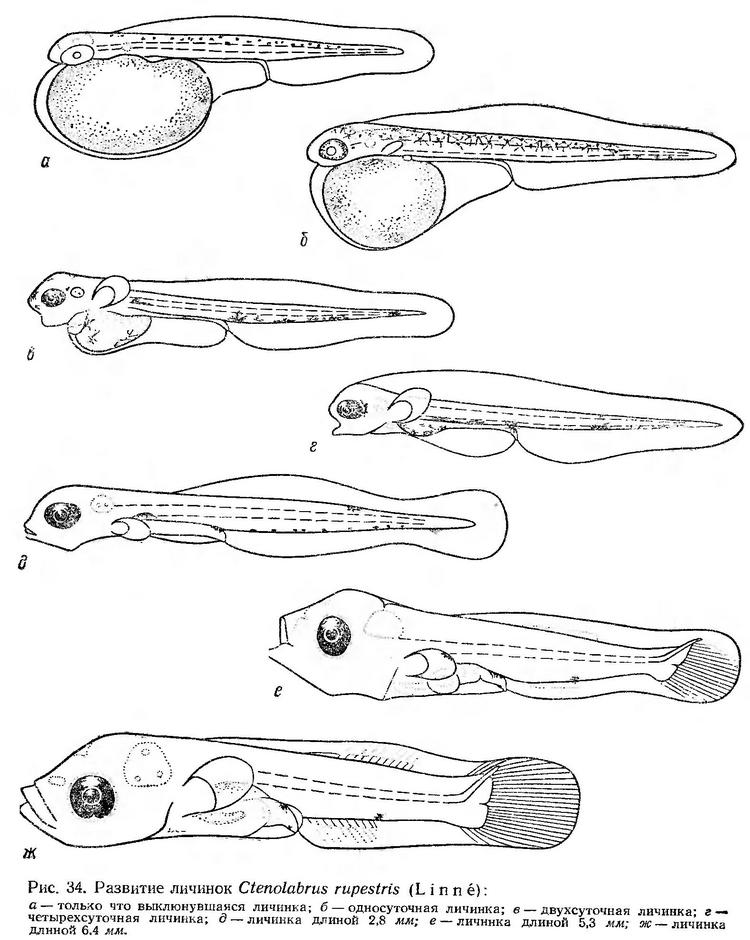
К двухсуточному возрасту остается небольшой остаток желточного мешка. Прорезается рот. Глаза интенсивно пигментированы. Грудные плавники приобретают подвижность. Пигментные клетки перемещаются на вентральную сторону тела (рис. 34, в).
На четвертые сутки после выклева желточный мешок резорбируется, рот приобретает подвижность. Пигментация ослабевает (рис. 34, г). К концу четвертых суток развития сохраняется 3—4 точечных меланофора на кишечнике, одна клетка у ануса и одна в средней части хвостового отдела. Личинки начинают плавать в нормальном горизонтальном положении. В условиях опыта они находятся в угнетенном состоянии. Периоды покоя длительные, перемещения кратковременные, скачкообразные. На пятые сутки личинки в опытах погибают.
Дальнейшее описание сделано по фиксированному материалу. У личинок длиной 2,8 мм желточный мешок резорбирован, оформляются челюсти и жаберный аппарат; плавательный пузырь наполнен воздухом, кишечник имеет вид прямой трубки. В нижней лопасти хвостового плавника намечаются опорные элементы. Спинная часть плавниковой каймы сокращается. Она начинается на уровне заднего края плавательного пузыря. Меланофоры образуют ряд вдоль брюшной стороны хвостового отдела. Пигментные пятна расположены на спинной стороне хвостового отдела и над плавательным пузырем (рис. 34, д).
У личинок длиной 5,3 мм тело более высокое. Челюсти сформированы. Жаберная крышка прикрывает жаберные лепестки. Кишечник петлеобразно изгибается. В хвостовом плавнике образовались лучи. Закладываются лучи в спинном и анальном плавниках. В хвостовом отделе на брюшной стороне остается одна пигментная клетка (рис. 34, е).
У личинок длиной 6,4 мм заканчивается формирование хвостового плавника, продолжается образование спинного и анального плавников. Грудные плавники большие, веерообразные. Верхняя челюсть становится выдвижной. Пигментное пятно в средней части хвостового отдела сохраняется (рис. 34, ж).
Распределение и численность икринок и личинок. Икринки гребенчатого губана входят в состав прибрежного планктона. Их распределение ограничено 5-мильной зоной. Икринки распределяются во всем верхнем теплом слое моря (до границы температурного скачка). В больших концентрациях икринки этого вида в Черном море не встречаются (Дехник и Павловская, 1950; Зайцев, 1959а; Овен, 1959). Сравнительно высокие (для этого вида) уловы отмечены Л. А. Дука (1959) в Севастопольской бухте в мае — до 424 икринок в одном поверхностном 10-минутном лове; в среднем на один лов — 124 икринки. В период наших наблюдений в районе Севастополя количество икринок Ctenolabrus rupestris не превышало 3—4 экз/м2, обычно насчитывалось не более 2 экз/10 м2.
Личинки губана встречаются в планктоне очень редко. В наших материалах найдено 6 личинок. На 0 м выловлено 2 личинки (2 лова), на 10 м — 1 личинка (1 лов), в слое 25—0 м — 3 личинки (3 лова).