Mullus barbatus ponticus Essipov, 1927
Синонимы:
Mullus barbatus ponticus Essipov, 1927
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
Барабулька, или султанка — Mullus barbatus ponticus (Essipov, 1927)
Профиль крутой головы плавно закруглен. Хвостовой плавник сильно вырезан. Зубы щетинковидные. D1 VIII. D2 I 8. А II 6. В срединном ряду 37—42 крупных легкоспадающих чешуй. Окраска тела красно-желтая с серебристыми «просветами», плавники желтые. Длина тела до 20 см. Прибрежная рыба, населяющая Черное море и прилежащие части Азовского моря.
Ихтиопланктон Черного моря. Т.В.Дехник 1973 г.
СУЛТАНКА — MULLUS BARBATUS PONTICUS ESSIPОV
Условия размножения. По наблюдениям многих исследователей нерест султанки в Черном море происходит летом. Икринки и личинки встречаются в планктоне с середины — конца мая по сентябрь. Массовый нерест начинается в середине — конце июня и заканчивается к августу (Зернов, 1913; Devedjian, 1926; Водяницкий, 1930а, 1936; Есипов, 1934; Косякина, 1938; Данилевский, 1939; Борисенко, 19406; Виноградов, 1949; Смирнов, 1949, 1953; Дехник и Павловская, 1950; Павловская, 1954; Ткачева, 1955; Зайцев, 1959а; Дука, 1959; Овен, 1959; Георгиев и др., 1960, 1961; Дехник и др., 1970).
Нерест султанки происходит у берегов и на мелководье на глубине не более 20 м. Распределение икры и личинок ограничено в основном прибрежной 5—10-мильной зоной (Дехник и Павловская, 1950; Павловская, 1954; Овен, 1959; Георгиев и др., 1960, 1961; Дехник и др., 1970). Нахождение икры и личинок в значительном удалении от берегов (Смирнов, 1949, 1953; Павловская, 1954; Георгиев и др., 1960, 1961) объясняется выносом их в эти районы течениями.
Температура воды в местах нахождения икринок колеблется в сравнительно небольших пределах — в основном от 15—17 до 25°. (Дехник и Павловская, 1950; Смирнов, 1949, 1950; Георгиев и др., 1960, 1961). Ю. П. Зайцев (1959а) отмечает нахождение икринок султанки при температуре воды 12°, А. И. Смирнов (1953) — при температуре 13,1°.
Соленость в районах лова икры и личинок султанки в Черном море колеблется от 13,5 до 19,3‰ (Дехник и Павловская, 1950; Георгиев и др., 1960, 1961). В южной части Азовского моря, где также размножается султанка, соленость не выше 11—12‰.
А. И. Смирнов (1953) отмечает, что икра султанки лишь изредка встречается в воде с соленостью выше средней солености черноморской воды и высказывает предположение о тяготении этого вида в период размножения к несколько опресненным районам.
В то же время опыты Л. С. Овен (1960) по влиянию различной солености на развитие и выживание икры и личинок султанки показали, что нормальное развитие икры и выклев жизнестойких личинок происходят при солености от 16 до 24‰. В воде с соленостью 9—15‰ развитие идет неправильно, наблюдается большая смертность икры и выклюнувшихся личинок.
Наблюдениями Ю. П. Зайцева (1959а) установлено, что в опресненном Одесском заливе султанка систематически не размножается. Здесь она нерестится только в годы, когда соленость воды не опускается ниже 13,5‰. В Средиземном море этот вид интенсивно размножается при очень высокой солености (37—38‰).
Из приведенных данных вытекает, что размножение и развитие икринок и личинок рассматриваемого вида происходит при значительном диапазоне солености.
В Мраморном море икринки и личинки обоих видов встречаются единичными экземплярами со второй половины мая до конца июля (Arim, 1957).
В Средиземном море оба обитающие там вида (Mullus barbatus и Mullus surmuletus) размножаются также в наиболее теплое время года — с мая по август (Montalenti, 1933).
Температура воды в Средиземном море в период размножения султанки близка к отмеченной для Черного моря, тогда как соленость, как известно, очень высокая (37—38‰).
Суточный ритм размножения. Проведенные ранее исследования (Дехник, 1959, 1961) показали четко выраженный суточный ритм размножения султанки. Все последующие наблюдения подтверждают закономерность ежесуточного поступления икринок в планктон в промежуток времени между 20 и 24 ч. Икринки на I этапе развития встречаются и в последующие часы (до 06). Однако в это время они находятся не на начальных стадиях развития, а в середине — конце I этапа (крупноклеточная и мелкоклеточная морула). В течение суток прослеживается последовательность развития каждого суточного вымета.
При температуре воды 23—24° в планктоне одновременно развиваются две суточные порции икринок — вымета ближайших ночных часов и вымета предшествующих суток. В определенный отрезок времени, когда инкубационный период икринок вымета предшествующих суток закончился, в планктоне встречаются икринки только одной суточной порции вымета ближайших ночных часов.
По данным Е. Г. Косякиной (1938), икрометание султанки начинается около 19 ч. Из наблюдений, проведенных А. И. Смирновым (1953), вытекает, что в ясную тихую погоду нерест султанки начинается в 19—21 ч, в пасмурную — в 15—16 ч. В ясную погоду нерест обычно продолжается до утра, в пасмурную — дольше. Нашими исследованиями не установлено нахождение свежевыметанных икринок султанки ранее 19—20 и позднее 04 ч.
Опыты Л. С. Овен (1961а, 1962а, 19626) показали, что каждая особь султанки ежесуточно выметывает большие порции икры в продолжение почти трех месяцев. Нерест в аквариумах наблюдался всегда между 22 и 24 ч (Овен, 1962а, 19626). Таким образом, ежесуточное поступление в планктон свежевыметанных икринок является результатом не только растянутого нереста популяции в целом, но и многопорционного нереста.
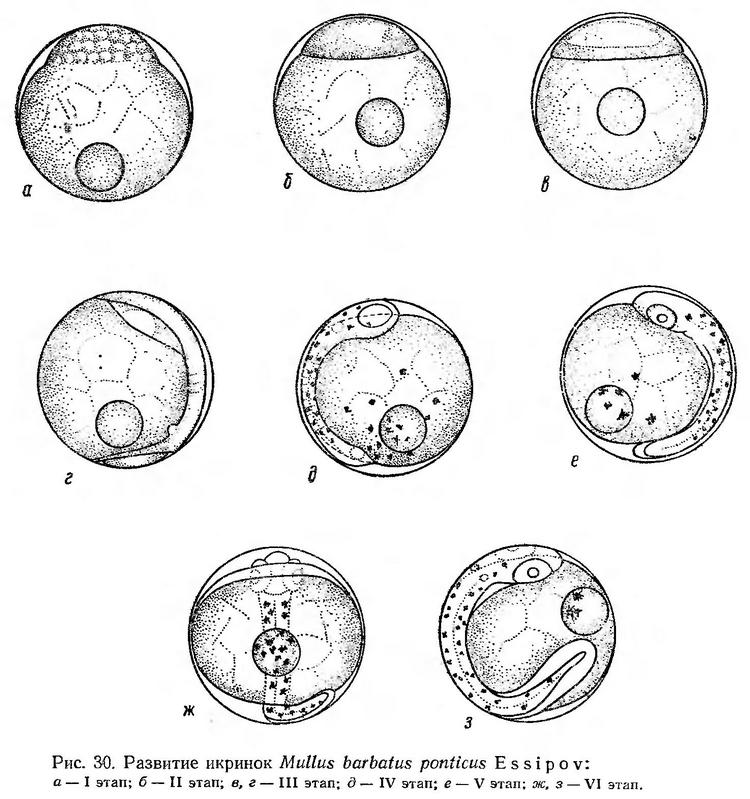
Строение икринок. Икринки султанки пелагические, сравнительно мелкие, с одной жировой каплей. Оболочка тонкая, прозрачная, желток в основном гомогенный и только поверхностный слой его разделен на крупные дольки (рис. 30). Диаметр икринок султанки в Черном море, по данным различных исследователей, колеблется от 0,61 до 0,82 мм, жировой капли — от 0,16 до 0,25 мм (Дехник и Павловская, 1950; Водяницкий и Казанова, 1954; Зайцев, 1959а; Георгиев и др., 1960). Размеры живых икринок по нашим измерениям в разные сроки нерестового сезона колеблются от 0,75 до 0,88 мм, жировой капли — от 0,18 до 0,23 мм; средний размер икринок — 0,81 мм, жировой капли — 0,21 мм.
По строению и развитию икринки султанки сходны с икринками ставриды. Икринки обоих видов имеют заходящие размеры, тонкую прозрачную оболочку, близкий темп развития при идентичных условиях. Жировая капля в икринках султанки в среднем несколько мельче, чем у ставриды, хотя встречаются также икринки с одинаковой по величине жировой каплей. Основные различия следующие: 1) в массе икринки и жировые капли у султанки мельче, чем у ставриды; 2) поверхность жировой капли у первого вида гладкая, блестящая в отличие от шероховатой матовой поверхности у второго вида; 3) у султанки сегментирован только поверхностный слой желтка, у ставриды желток сегментирован полностью; 4) в процессе развития у эмбрионов ставриды на VI этапе появляется каротиноидный пигмент, у эмбрионов султанки его нет.
Икринки и личинки Mullus barbatus из Средиземного моря не описаны. Монталенти (Montalenti, 1933) отмечает, что различия в строении икринок Mullus barbatus и Mullus surmuletus не известны.
Эмбриональное развитие султанки детально описано А. И. Смирновым (1949, 1953). Наши наблюдения за развитием султанки проводились на искусственно оплодотворенных икринках и на икринках, собранных в море в период между 21 и 24 ч, когда в планктоне встречались только что выметанные икринки. Опыты по развитию проводились в лабораторных условиях с нормальным суточным освещением при температуре воды 22,1—23,4°.
При указанной температуре стягивание протоплазмы к анимальному полюсу происходит в течение 1 ч 30 мин — 1 ч 45 мин, процесс дробления бластомеров длится около 4 ч.
На стадии крупноклеточной морулы длина основания бластодиска составляет 0,62—0,65 мм, высота — 0,18—0,20 мм (рис. 30, а; табл. 26). Эпителиальная бластула при указанной температуре формируется в течение 2—2,5 ч. Бластодиск чуть уплощен; при поворотах икринки хорошо просматривается бластоцель (рис. 30, б).
Через 8—8,5 ч после оплодотворения начинается обрастание желтка бластодермой и гаструляция (рис. 30, в). Когда бластодерма охватывает около 1/3 поверхности желтка, четко обозначается зародышевое утолщение. Через 16 ч от начала развития остается необросшей небольшая желточная пробка. Головной конец зародыша заметно расширен, намечаются глаза и два первых туловищных сегмента (рис. 30, г). Этап гаструляции продолжается 10—10,5 ч.
После замыкания бластопора на IV этапе развития продолжается сегментация туловищного отдела, обособляются глаза, намечаются слуховые пузырьки, формируются мозговые доли. На теле, на жировой капле и на желтке появляются меланофоры (рис. 30, д). Следует отметить, что на желтке меланофоры имеются не всегда. При температуре воды 23,1—23,4° этап продолжался 5,5—6 ч.
В возрасте 21—22 ч появляется хвостовая почка и начинается быстрый рост хвостового отдела. С ростом хвостового отдела продолжается дальнейшая дифференцировка миотомов, появляются зачатки сердца, печени, кишечника, намечается плавниковая складка (рис. 30, е).
Пульсация сердца, как правило, начинается в возрасте 28—29 ч. В минуту насчитывается 110—115 биений сердца. К этому времени эмбрион охватывает около 2/3 поверхности желтка (рис. 30, ж). Одновременно с пульсацией сердца наблюдаются слабые подергивания эмбриона. Перед выклевом эмбрион энергично двигается внутри яйцевой оболочки. Голова прижимается к оболочке, выпячивает ее и разрывает (рис. 30, з). Эмбрион сильными толчками довольно быстро (в течение нескольких минут) освобождается от оболочки.
Эмбриональное развитие при температуре воды 22,1—23,4° продолжается 34—35 ч (табл. 26). Понижение температуры значительно удлиняет процесс развития. Так, при температуре 17,5—18,5и развитие продолжается около 50 ч (Смирнов, 1953; Овен, 1960).
Постэмбриональное развитие. Только что выклюнувшиеся личинки султанки очень мелкие (1,5—1,9 мм длиной). Они слабо развиты. Грудных плавников нет; голова плотно прижата к большому яйцевидному желточному мешку, передняя часть которого сильно выдается за голову. Тело выклюнувшихся личинок пигментировано только меланофорами в отличие от ставриды, у которой уже на VI этапе эмбрионального периода появляется каротиноидный пигмент. Меланофоры расположены на спине и боковых поверхностях тела, на жировой капле и желточном мешке (рис. 31, а).
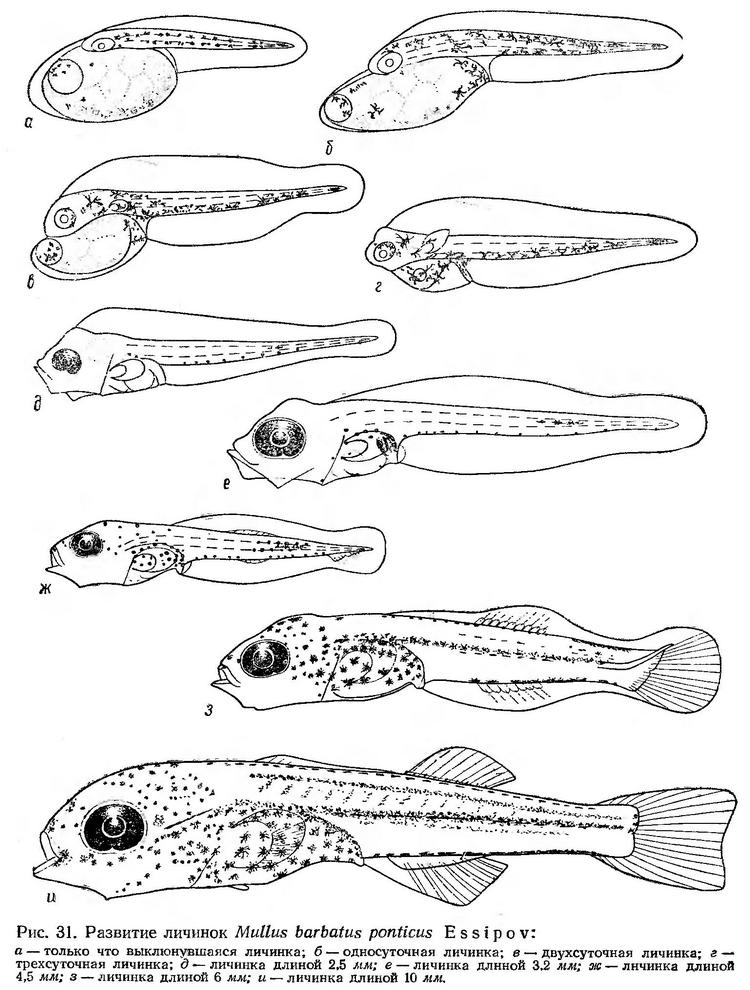
Жировая капля находится в верхней части переднего края желточного мешка и в основном определяет положение личинок в воде. Они как бы подвешиваются почти вертикально к поверхностной пленке и пассивно парят в воде или медленно погружаются не меняя положения.
Длительное время личинки находятся в покое. Слабые круговые движения вокруг оси тела или винтообразные, направленные вверх, движения осуществляются с помощью волнообразных изгибов хвоста. Иногда личинки совершают быстрые короткие броски. Поведение выклюнувшихся личинок султанки и ставриды очень сходно.
В течение первых суток развития желточный мешок еще более вытягивается и далеко выдается за голову (рис. 32, б). Меланинная пигментация усиливается, меланофоры распространяются на вентральную сторону тела. Длина личинки увеличивается в среднем до 2,2 мм (без желточного мешка). Личинки принимают наклонное положение. Они плавают под углом к поверхности, головой и брюшной стороной ориентированы кверху. Заметно удлиняется постанальный отдел. Подвижность личинок возрастает, круговые движения заменяются прямолинейньми. Периоды покоя в несколько раз длиннее периодов движения. Сердце пульсирует быстро (140—150 биений в минуту), хотя наполнение очень слабое.
К двухсуточному возрасту желточный мешок уменьшается в размере, но голова еще плотно к нему прижата. Появляются зачатки грудных плавников. В спинной плавниковой кайме образуется оводненная полость (рис. 31, в). Положение личинок в воде снова несколько изменяется. Теперь они плавают в горизонтальном положении брюшной стороной вверх. Сверху личинки имеют булавовидную форму с блестящей каплей в передней расширенной части, снизу личинки имеют нитевидную форму и, по-видимому, мало заметны. Меланофоры в основном перемещаются на вентральную сторону. Периоды покоя колеблются от 5 — 10 до 60 сек. Движения по-прежнему кратковременные, продолжительностью не более 1 — 2 сек. Личинки производят стремительные броски, не меняя положения тела в воде.
К трехсуточному возрасту голова начинает освобождаться от желточного мешка, который к этому времени значительно уменьшается. Заметно сокращается и жировая капля. На нижней стороне головы намечается ротовая ямка. Увеличиваются зачатки грудных плавников; появляется пигмент в глазах (рис. 31, г). Спинная полость увеличивается. Меланофоры почти полностью перемещаются на вентральную сторону. Личинки продолжают плавать в перевернутом положении. Периоды покоя по-прежнему длительные — до 1,5—2 мин, но и периоды движения заметно увеличиваются. Теперь личинки делают стремительные броски на расстояние до 5—7 мм. На прикосновение какого-либо предмета реагируют резким броском в сторону.
В возрасте четырех суток у личинок султанки прорезается рот. Сохраняется лишь небольшой остаток желтка. Личинки принимают нормальное положение — плавают спиной вверх. Значительно возрастает скорость их движения — от 8,6 см/мин в двухсуточном возрасте до 21,5 см/мин — в четырехсуточном. В естественных условиях личинки в этом возрасте переходят на внешнее питание. В пищевом комке личинок длиной 2,2—2,5 мм встречаются до 7—8 видов планктонных организмов (Дука, Синюкова и др., 1970).
В возрасте 5 суток все личинки султанки в наших опытах погибали. Дальнейшее описание дано по фиксированному материалу.
У личинок длиной около 2,5 мм сохраняется незначительный остаток желтка; кишечник утолщается и образует петлю; формируются челюстной и жаберный аппараты (рис. 31, д). Глаза приобретают интенсивно черную окраску, и с этого времени личинки, по-видимому, начинают различать плавающие в воде предметы. Такие личинки, по данным В. И. Синюковой (1969), питаются в основном науплиусами Copepoda, которые составляют более 80% общего количества заглоченных организмов.
У личинок длиной 3,2 мм (рис. 31, е) тело прогонистое с коротким туловищем и длинным хвостовым отделом. Грудные плавники большие, веерообразные. Излюбленной пищей у этих личинок по-прежнему остаются науплиусы Copepoda. Некоторое значение в питании приобретают и более крупные зоопланктеры, такие, как взрослые формы Oithona minuta. Науплиусы и взрослые Oithona minuta составляют 90—95% веса пищевого комка, а только науплиусы — 77—78% (Дука, Синюкова и др., 1970).
У личинки длиной около 4,5 мм начинают формироваться непарные плавники (рис. 31, ж). Челюстной и жаберный аппараты сформированы. В питании личинок значительно возрастает роль Oithona minuta, которая составляет в отдельных случаях до 70% веса пищевого комка. Заметную роль начинают играть также такие организмы, как Podon polyphemoides (до 27% по весу) и Penilia avirostris (до 5%).
У личинок длиной 6—10 мм заканчивается формирование непарных плавников, появляются зачатки брюшных плавников (рис. 31, з, и). В питании таких личинок науплиусы играют незначительную роль (до 2% веса пищевого комка). Излюбленным пищевым организмом остается Oithona minuta, которая составляет 70% по весу и по количеству от всего наполнения кишечника. Заметную роль в питании этих личинок играют крупные зоопланктеры, как Penilia avirostris и Podon polyphemoides (Дука, Синюкова и др., 1970).
Личинки султанки в светлое время суток с пустыми кишечниками не встречаются. Обладая значительно большей скоростью движения, чем скорость их жертв, они, по-видимому, быстро находят скопления планктона и держатся на этих скоплениях, перемещаясь вместе с ними.
Распределение и численность икринок и личинок. В летнее время вдоль всего побережья Черного моря встречаются развивающиеся икринки и личинки султанки.
Икринки султанки, как и других тепловодных рыб, распределяются во всем верхнем теплом слое, однако концентрируются они в основном у поверхности. Горизонтальные ловы на поверхности и на глубине 10 м показывают заметное преобладание икринок султанки в поверхностном слое моря (табл. 27).
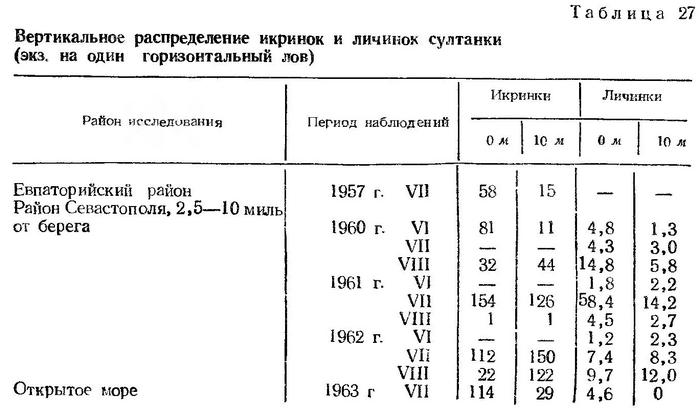
В распределении личинок султанки в отличие от двух других массовых видов — хамсы и ставриды — выявляется тяготение к поверхностному слою. При проведении параллельных ловов на 0 и 10 м почти во всех случаях отмечено преобладание личинок в верхнем горизонте (табл. 27).
Численность икринок и личинок султанки в планктоне существенно изменяется в течение нерестового сезона и значительно колеблется по годам.
В мае при температуре воды 12—13° в планктоне начинают встречаться единичные икринки султанки. Личинки появляются значительно позже, обычно в середине июня, когда температура воды повысится до 18—19°, при этом численность их в это время очень невелика (табл. 28).
Разгар нереста султанки приходится на июль. В это время отмечены наиболее высокие концентрации икринок в планктоне и сравнительно высокая численность личинок (табл. 28). В нересте принимают участие все нерестовые популяции, вымет икры происходит ежесуточно, о чем свидетельствуют суточные изменения коэффициента зрелости гонад (Овен, 1961а, 1962а) и прослеживаемая последовательность развития суточных выметов икринок (Дехник, 1961).

В августе интенсивность нереста султанки снижается. Численность икринок в планктоне уменьшается, а количество личинок, наоборот, в это время наибольшее за счет расширения размерного ряда и преобладания крупных личинок.
В сентябре икринки и личинки встречаются очень редко, причем единичными экземплярами.
Многосуточные ловы ихтиопланктона в районе Севастополя свидетельствуют о дисперсности распределения личинок.