Sprattus sprattus (Linnaeus, 1758)
Шпрот, европейский шпрот, атлантическая килька, северный шпрот, балтийский шпрот, балтийская килька, ревельская килька, южноевропейский шпрот, черноморский шпрот, черноморская килька, средиземноморский шпрот, средиземноморская килька (Rus),
European sprat, Baltic sprat, Sprat, Black sea sprat
(Eng)
Синонимы:
Clupanodon phalerica Risso, 1827
Clupea latulus Cuvier, 1829
Clupea papalina Bonaparte, 1846
Clupea phalerica Smitt, 1896
Clupea schoneveldii Krøyer, 1846
Clupea sprattus balticus Schneider, 1908
Clupea sprattus Linnaeus, 1758
Clupea sulinae Antipa, 1904
Meletta phalerica Fage, 1908
Meletta vulgaris Valenciennes, 1847
Spratella pumila Valenciennes, 1847
Spratella serdinka Nikolskii, 1923
Spratella sprattus baltica Berg, 1932
Spratella sprattus phalerica Berg, 1931
Sprattus sprattus balticus (Schneider, 1908) Балтийский шпрот, балтийская килька, ревельская килька (Rus),
Sprattus sprattus phalericus (Risso, 1827) Черноморский шпрот, южноевропейский шпрот, черноморская килька, средиземноморская килька (Rus),
Sprattus sprattus sprattus (Linnaeus, 1758) Северный шпрот, атлантическая килька (Rus),
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
Шпрот балтийский — Sprattus sprattus balticus (Schneider, 1904)
У представителей рода рот верхний, маленький. Последние лучи анального плавника нормальной длины. Брюшные плавники лежат впереди или под началом основания спинного плавника. Тело сравнительно высокое. В киле 31—34 чешуйки. Морская рыба, зоопланктофаг. Длина тела примерно 17 см. Встречается в Балтийском море.
Шпрот черноморский — Sprattus sprattus phalericus (Risso, 1826)
Обычный подвид в Черном море. От балтийского подвида отличается низким прогонистым телом и меньшим числом килевых чешуй. Длина тела не превышает 16 см.
Декоративное рыбоводство. А.М.Кочетов 1991 г.
При содержании рыб воду лучше подсаливать (5—18 ‰). В аквариуме содержат следующие виды:
Балтийский шпрот - Sprattus sprattus balticus (Schneider, 1904), 16 см.
Ихтиопланктон Черного моря. Т.В.Дехник 1973 г.
СЕМ. CLUPEIDAE — СЕЛЬДЕВЫЕ
Из трех собственно морских видов этого семейства, обитающих в Черном море (Расс, 1949, 1965; Световидов, 1964), достоверно размножается здесь один вид — Sprattus sprattus phalericus Rissо.
ШПРОТ — SPRATTUS SPRATTUS PHALERICUS (RISSО)
Условия размножения. Впервые нахождение икринок шпрота в Черном море было отмечено В. А. Водяницким (1930а), который указывает, что икринки этого вида встречаются исключительно в холодное время года при температуре воды 5—15°.
Дальнейшими исследованиями было установлено, что шпрот размножается на протяжении почти целого года на всем пространстве Черного моря. Нерест его происходит всегда в холодной воде. Зимой нерестовые популяции распространяются широко — от поверхности до 100 м глубины, летом нерестующий шпрот держится ниже слоя температурного скачка. Наиболее интенсивный нерест происходит зимой — с декабря по февраль (Сыроватский, 1934; Павловская, 1952, 1954; Зайцев, 1953a, 1959а; Асланова, 1954; Алеев, 1958; Горбунова, 1958; Cautie, 1958; Георгиев и др., 1960).
Икринки шпрота встречаются в Черном море при температуре воды от 5 до 19° и солености 13,5—19‰. Максимальные уловы отмечены при температуре воды 6,8—12,4° (Павловская, 1954). Сроки нереста этого вида в разных районах его распространения определяются, как отмечает Н. Н. Горбунова (1958), в основном температурой воды. Как в северных, так и в южных районах ареала разгар нереста происходит при температуре воды 8—12°. В результате строгой приуроченности нереста к определенному уровню температуры воды его сроки смещаются по мере продвижения с юга на север с зимнего на весенне-летний сезон.
Данных о размножении шпрота в Средиземном море в литературе мало. Икринки его встречаются здесь в планктоне очень редко (D'Ancona, 1933а). Холт (Holt, цит. по D'Ancona, 1933а) отмечает их нахождение в районе Марселя в марте, Фаж (Fage, 1920) — в том же Лионском заливе в феврале.
По Фюрнестену (Furnestin, 1948), нерест у берегов Франции происходит с декабря по апрель. В Адриатическом море икринки встречаются в планктоне в декабре и январе (Graeffe, цит. по D'Ancona, 1933а; Fage, 1920).
В южной половине Северного моря нерест этого вида начинается в марте, достигает максимума в июне и заканчивается в августе. (Ehrenbaum, 1905—1909; Aurich, 1942). В районе Плимута икра шпрота встречается непрерывно с середины февраля до середины июня, икринки обнаруживаются в планктоне до середины июля, но в небольшом количестве (Hefford, 1910), по более поздним данным (Lebour, 1921) — с января, наиболее многочисленны — в феврале — марте.
Нерест шпрота в Северном море происходит главным образом вблизи берегов (Ehrenbaum, 1905—1909; Hefford, 1910; Aurich, 1942).
Икра и личинки встречаются в Северном море при температуре воды от 5 до 16° и солености 34,5—35,0‰. Максимальная концентрация икринок отмечена при температуре 7—16° (Aurich, 1942).
В Балтийском море шпрот Sprattus sprattus balticus (Schneider) нерестится с марта по август. В южных районах разгар нереста наблюдается в конце мая — июне; в северной части моря в это время нерест только начинается (Казанова и Перцева-Остроумова, 1960). В Гданьской бухте нерест начинается в первой половине мая и продолжается до конца июля, разгар — в конце июня и первой половине июля (Mankowski, 1948). Нерест шпрота в Балтийском море происходит на всей его площади, однако преимущественно в открытом море: чем ближе к берегам, тем нерест менее интенсивен (Mankowski, 1948; Казанова и Перцева-Остроумова, 1960). Нерестовые популяции, как и в Черном море, распределяются в толще воды и у поверхности в зависимости от сезона. В апреле — мае шпрот размножается обычно в толще воды, в июне — июле главным образом в верхних слоях воды (Казанова, Перцева-Остроумова, 1960). Авторы, к сожалению, не приводят данных по температуре воды в районах концентрации нерестовых популяций. Температура воды в период массового нереста шпрота в центральной части Балтийского моря колеблется от 8,1 до 11,2°. Верхний уровень температуры, при которой отмечался нерест, достигал 16,2—17,9° (Mulicki, 1948). И. И. Казанова и Перцева-Остроумова (1960) отмечают значительную эвригалинность балтийского шпрота, который может размножаться при солености 6‰ и даже более низкой.
Приведенные данные свидетельствуют о том, что основным условием, ограничивающим период размножения шпрота, является температура воды. Интенсивный нерест шпрота повсеместно происходит при температуре от 6—7 до 12°. Наоборот, соленость воды в период размножения этого вида в пределах ареала колеблется в очень больших пределах — от 4—5 (Балтийское море) до 37—38‰ (Средиземное море). Таким образом, наряду с четко выраженной стенотермностью икринки и личинки этого вида характеризуются эвригалинностью.
В распределении нерестовых популяций шпрота, а следовательно икринок и личинок, выявляется интересная закономерность, отмеченная также для Engraulis encrasicholus (Дехник, 19636). В морях с низкой соленостью (Черное и Балтийское) шпрот широко распределяется по всему пространству; нерест его происходит как в открытом море, так и в незначительном удалении от берегов. В морях с высокой соленостью (Средиземное и Северное) шпрот нерестится в основном вблизи берегов.
Строение икринок. Икринки шпрота пелагические, сферической формы. Оболочка прозрачная, гладкая; перивителлиновое пространство небольшое. Желток состоит из крупных гранул. Жировой капли нет.
Величина икринок шпрота в Черном море колеблется от 0,90—0,92 до 1,22—1,23 мм (Водяницкий и Казанова, 1954; Горбунова, 1958; Георгиев и Др., 1960). По нашим данным, диаметр икринок, фиксированных формалином, изменяется от 0,98 до 1,35 мм. В Одесском заливе величина икринок колеблется от 0,98 до 1,21 мм (Зайцев, 1958а, 1959а). Размер икринок шпрота из разных районов Черного моря в разные сроки нерестового сезона приведен в табл. 1 (по Н. Н. Горбуновой, 1958). Средняя величина икринок (по южно-крымскому району) увеличивается от ноября к февралю от 1,0 до 1,15 мм, что Н. Н. Горбунова связывает с последовательным понижением температуры в этот период. Такой вывод согласуется с приведенными ниже данными для Северного и Балтийского морей. Сопоставляя размеры икринок шпрота с соленостью воды в пределах его ареала, Н. Н. Горбунова (1958) выявила закономерное увеличение размера икринок по мере понижения солености (табл. 2).
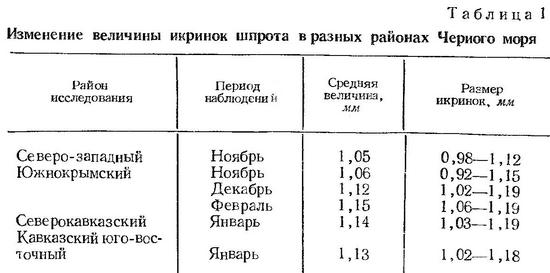
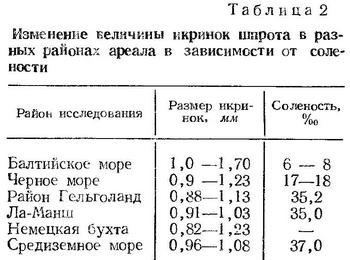
По данным Эренбаума (Ehrenbaum, 1905—1909), средний размер икринок шпрота уменьшается от марта к июлю от 1,07 до 0,93 мм. По Хеффорду (Hefford, 1910), средний диаметр икринок этого вида в районе Плимута в феврале — марте составляет 1,031 мм, в апреле — мае — 0,973 мм, в июне — июле — 0,912 мм. В Балтийском море средний размер икринок уменьшается от мая к августу от 1,20 до 1,05 мм (Ehrenbaum, 1905—1909).
Эмбриональное развитие шпрота прослежено в основном по фиксированному материалу. Изучено также строение живых икринок на V и VI этапах развития (рис. 1).
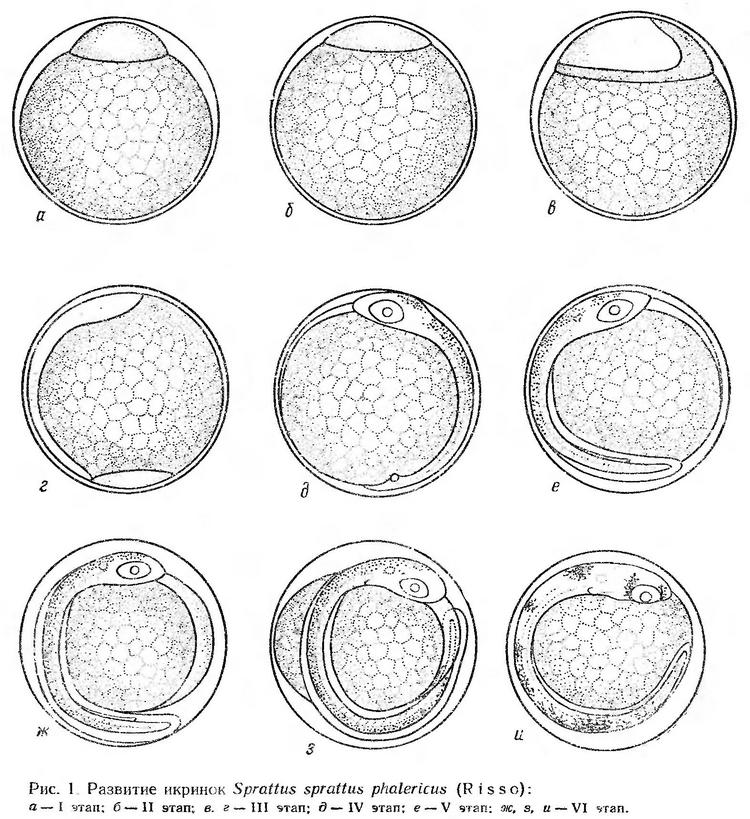
Эмбриональное развитие шпрота при колебаниях температуры воды от 5 до 13° продолжается 8 суток (Горбунова, 1958). Бластодиск на этапе бластомерной бластулы занимает очень небольшую часть поверхности желтка. Длина его основания около 0,55 мм, высота — 0,22—0,23 мм (рис. 1, а). На этапе эпителиальной бластулы длина основания увеличивается до 0,60 мм, высота уменьшается до 0,17—0,18 мм (рис. 1, б). Зародышевое утолщение намечается вскоре после начала обрастания желтка бластодиском (рис. 1, в).
Незадолго перед замыканием бластопора появляются первые туловищные сегменты (рис. 1, г). После замыкания бластопора число миотомов быстро увеличивается, формируются глаза, намечается нервный тяж, мозговые доли, слуховые капсулы. Ко времени отделения хвоста от желтка в глазах образуются хрусталики. На голове и спинной стороне тела появляются мелкие точечные меланофоры (рис. 1, д). Когда эмбрион охватывает около 2/3 поверхности желтка четко прослеживается пульсация сердца (64—69 сокращений в минуту). Эмбрион подергивается. К этому времени формируется кишечник, намечается плавниковая кайма (рис. 1 е, ж).
К моменту выклева хвост эмбриона почти смыкается с головой, появляются зачатки грудных плавников. В слуховых капсулах закладываются отолиты. Мелкие точечные меланофоры расположены на голове (от уровня средней линии глаза) и по спинной стороне вдоль всего туловищного отдела (рис. 1, з). Эмбрион энергично двигается внутри оболочки. Такие энергичные подергивания происходят 2—3 раза в минуту. Моменты движения продолжаются 1—3 сек.
И. И. Казанова (1953) отмечает значительное разнообразие в характере и интенсивности пигментации эмбрионов черноморского шпрота. Наряду с икринками со слабой точечной меланинной пигментацией эмбриона, как у типичной формы, встречаются икринки с очень интенсивной пятнистой пигментацией. Установлено, что эти два типа пигментации связаны промежуточными вариациями.
Такие различия выявлены также и нами при просмотре массового материала (рис. 1, и). При этом икринки с различной пигментацией эмбрионов, как отмечено И. И. Казановой (1953), встречаются в одной и той же пробе, т. е. при одинаковых условиях. И, наоборот, икринки одинаково пигментированные вылавливаются с разных горизонтов, т. е. из различных условий освещения и при разных температуре и солености.
Постэмбриональное развитие. Недавно выклюнувшиеся личинки имеют большой яйцевидный желточный мешок. Голова плотно прижата к его переднему краю. Тело длинное, тонкое. Его высота примерно в 20 раз меньше длины. Анальное отверстие открывается в край плавниковой каймы в последней четверти тела. Антеанальное расстояние составляег 84—89% длины тела. Глаза непигментированы. На голове и вдоль всей спинной стороны туловища расположены мелкие точечные пигментные клетки (рис. 2, а). Длина только что выклюнувшихся личинок, по Н. Н. Горбуновой (1958), колеблется от 2,80 до 3,68 мм. По данным Ю. П. Зайцева (1958а, 1959а), длина выклюнувшихся личинок в Одесском заливе 2,91—3,30 мм, причем очень немного личинок менее 3,0 мм. Размер выклюнувшихся личинок (менее 2 мм) приведен В. А. Водяницким и И. И. Казановой (1954), по-видимому, по фиксированному материалу. Размер только что выклюнувшихся личинок этого вида из Северного моря (район Плимута) — 2,25 мм (Hefford, 1910).
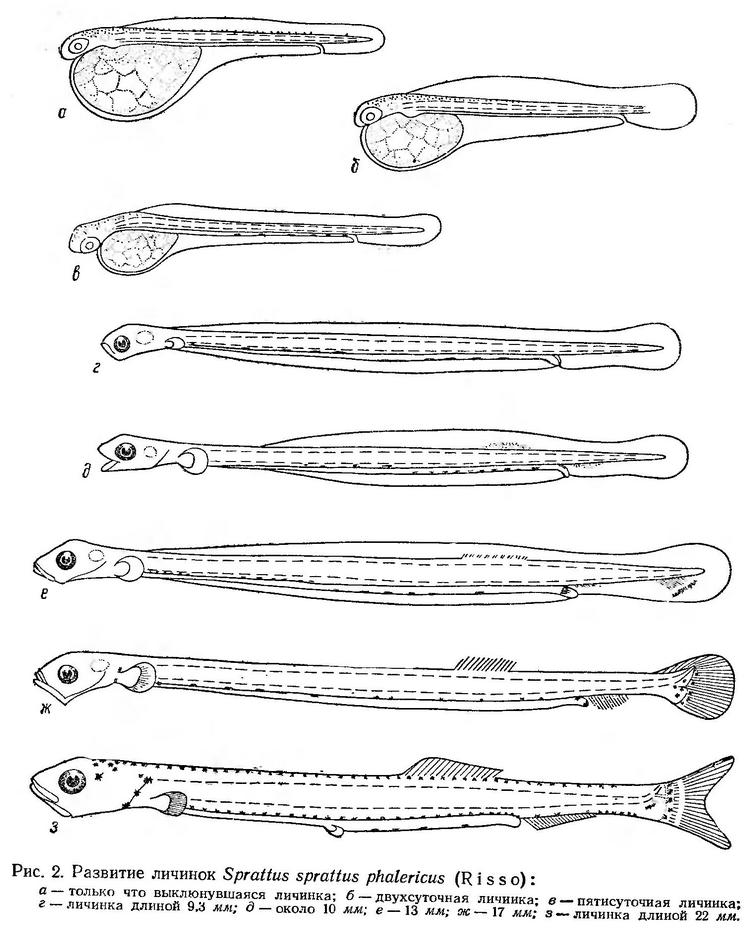
Личинки шпрота сразу после выклева начинают быстро и энергично двигаться при помощи волнообразных изгибов тонкого длинного тела. Периоды разового движения составляют 5—10 сек, затем наступает короткая пауза (от 5 до 30—40 сек), за которой следуют снова энергичные, сравнительно длительные перемещения. Дальнейшее описание личинок (за исключением личинки длиной 9,3 мм (рис. 2, г), зарисованной по нашим материалам) дано по Н. Н. Горбуновой (1958).
У двухсуточных личинок желточный мешок заметно уменьшается, составляя 31,8% длины тела. Распределение пигментных клеток остается без изменения (рис. 2,6). На шестые сутки желточный мешок уменьшается до 23%. Личинка достигает 4,5 мм длины. Антеанальное расстояние сокращается до 79,5%. Голова отделяется от желтка, хорошо выражена ротовая ямка. На спинной стороне пигмент сохраняется только в области головы. Обозначен четкий пигментный ряд вдоль кишечника в его задней половине. Начинается пигментация глаз (рис. 2, в).
По наблюдениям Н. Н. Горбуновой (1958), желточный мешок рассасывается при длине около 6 мм. Глаза к этому времени уже полностью пигментированы и личинки, по-видимому, могут различать предметы, что связано с началом активного (внешнего) питания. У личинок 6 мм длиной в кишечниках встречаются науплиальные стадии Copepoda и взрослые формы Oithona minuta (Горбунова, 1958). Период смешанного питания не установлен. Можно предположить, что он начинается при длине личинки около 5 мм (у личинки длиной 4,3—4,5 мм еще нет ротового отверстия).
У личинок длиной 9,3 мм (рис. 2, г) еще полностью сохраняется эмбриональная плавниковая кайма. Грудные плавники очень маленькие; движение осуществляется по-прежнему при помощи волнообразных изгибов тела. Пигментные клетки расположены вдоль верхнего (в передней половине тела) и нижнего (в задней половине) края кишечника.
При длине личинок около 10 мм (рис. 2, д) начинается образование оснований спинного и анального плавников. Плавниковая кайма несколько сокращается.
У личинок длиной 13—14 мм (рис. 2, е) в спинном плавнике обозначены зачатки 9 лучей, в анальном — 4. В хвостовом плавнике формируются гипуралии, намечается изгиб уростиля.
Когда личинки достигают 17—18 мм (рис. 2, ж) в спинном и анальном плавниках у них насчитывается по 13 лучей. Формирование хвостового плавника заканчивается. Уростиль изогнут кверху. Появляются лучи в грудных плавниках.
При длине личинок 22—23 мм (рис. 2, з) хвостовой плавник почти полностью сформирован. В спинном плавнике имеется 16 лучей, в анальном — 20. Появляются зачатки брюшных плавников. Личиночная пигментация исчезает. Появляются пигментные клетки вдоль всей спинной стороны тела и по нижнему краю кишечника в его задней половине. У основания хвостового плавника и в области жаберной крышки расположены крупные звездчатые меланофоры. У личинок длиной 24—25 мм, как указывает Н. Н. Горбунова (1958), период личиночного развития заканчивается.
Распределение и численность икринок и личинок. Обстоятельные данные по распределению и численности икры и личинок шпрота в Черном море содержатся в работах Р. М. Павловской (1954) и П. Н. Горбуновой (1958). Данные этих авторов, как и наши наблюдения, проведенные в декабре 1958 и в январе — феврале 1959 г. у берегов Кавказа, Крыма и в северо-западной части моря, показали, что икра и личинки шпрота встречаются на всем пространстве Черного моря, как у берегов, так и в открытых районах.
Вертикальное распределение икры подвержено некоторым изменениям в зависимости от сезона года. В начале нерестового сезона (конец августа начало сентября) икринки встречаютвся в слое 50—80 м. В конце сентября отдельные икринки появляются и в поверхностных слоях. В период массового нереста — с декабря по февраль икра распределяется во всем слое — от поверхности до 100 м глубины. В марте — апреле в поверхностных слоях икринки шпрота продолжают встречаться в значительном количестве. С конца апреля — середины мая они распределяются, как и в начале нерестового сезона, ниже слоя 50 м. Максимальные концентрации икры шпрота во все
сезоны года образуются в толще воды — в слое 25—50 или 50—100 м.
В табл. 3 показано вертикальное распределение икры шпрота в январе — феврале 1959 г. у побережья Крыма и Кавказа. Эти данные свидетельствуют о том, что в разгар нерестового сезона в период гомотермии основной нерест происходит в толще воды и икринки развиваются, по-видимому, в том же слое воды, в котором они были выметаны.
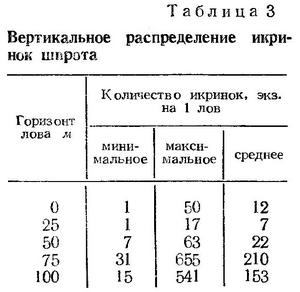
Таким образом, как летом в условиях температурной стратификации, так и зимой при гомотермии развитие икры шпрота происходит в основном в одних и тех же слоях моря.
Численность икры шпрота в планктоне Черного моря колеблется от 2—10 до более чем 200 экз/м2 (Горбунова, 1958).
В декабре 1958 г. количество икры шпрота, по нашим данньм, в северозападной части Черного моря колебалось от 2 до 90 экз/м2 (в среднем 24 экз/м2), у берегов Крыма — от 2 до 24 (в среднем 12 экз/м2). В январе — феврале 1959 г. у побережья Кавказа количество икры достигало 220 экз/м2. В среднем численность икры в планктоне в это время составляла 59 экз/м2.
В Одесском заливе количество икры шпрота в планктоне, по наблюдениям Ю. П. Зайцева (1959а), не превышало 1 экз/м2. Личинки шпрота распределяются широко по всему пространству моря. В их вертикальном распределении не выявлено строгой приуроченности к какому-либо горизонту. Они распространяются равномерно во всем слое от 0 до 100 м (Горбунова, 1958; наши наблюдения). В Новороссийской бухте личинки ловились от поверхности до глубины 30 м. Наибольшее количество (29 личинок) было собрано из поверхностного слоя перед входом в бухту (Пчелина, 1936, 1940). Держатся личинки шпрота обычно разреженно. В уловах чаще всего встречаются единичные экземпляры. Максимальное количество (до 30 экз/м2) было обнаружено Н. Н. Горбуновой (1958) в открытом море в районе между Керчью и Ялтой. По нашим наблюдениям, в Севастопольской бухте в 1966— 1967 гг. средняя численность личинок шпрота составляла около 2 экз/м2.
В Средиземном море, как было отмечено, икринки и личинки шпрота чрезвычайно малочисленны.
Детальные данные по численности икры этого вида в разных районах Северного моря приводит Аурих (Aurich, 1942). У восточного побережья Англии, у южного и юго-западного побережья Норвегии количество икры шпрота в планктоне обычно не превышает 100 экз/м2. В южной части Северного моря численность икры в разгар нереста достигает свыше 300—400 экз/м2.
Район от пролива Скагеррак на запад до 4° восточной долготы считается самым большим нерестилищем шпрота. Здесь плотность икринок достигает 1000—2000 экз/м2. Такая высокая концентрация нигде больше не была отмечена (Aurich, 1942).
Приведенные данные по количественному распределению икры прота в планктоне дают общее представление о сравнительной численности популяций этого вида в разных районах ареала.
Словарь названий пресноводных рыб СССР. Г.У.Линдберг и А.С.Герд 1972 г.
3.4.1 (1). Sprattus sprattus balticus (Schneider, 1904) — Балтийская килька (Св., 1952 : 112).
[1] русск. Балтийская килька — Б., 1932 : 99. Балтийский шпрот — Г. Ник., 1954 : 116. Брейтлинг — Гр. : 215. Килька Балт. м., Ревельская килька Балт. м., Шпрот Балт. м. — Гейнем.: 501. [4] польск. Bretlink, Bydlinek, Kilka, Sprat, Sprotta — Majew. Szprot — Majew.; Names of fishes : 37. [12] латыш. Bretlina — ZK. [13] лит. Silkelis — Majew. [14] дат. Brisling — Names of fishes : 36. [15] шведск. Hvassbuken — Smitt : 974. Skarpsill — Names of fishes : 37. Skarpsillen — Smitt : 974. Vassbuk — Б., 19406 : 9; Names of fishes : 37. [16] норв. Brisling — Names of fishes : 37. [17] исл. Brislingur — Names of fishes : 37. [18] англ. Baltic sprat Канада — Ricker : 152. Sprat — Smitt : 974; Poll : 133; Names of fishes : 36; Европа и Сев. Африка — Мартинсен, 1961 : 87. [19] нем. Breitling, Sprott — Names of fisches : 37. [20] голланд. Bliek, Sardijn, Schardijn, Sprot — Poll : 133. [21] флам. Sprot — Names of fishes : 36. [22] франц. Esprot, Sprot — Poll : 133. [36] финск. Kilo — Б., 19406 : 9. Kilohaili — Names of fishes : 36; Brofeldt : 157. [38] эст. Kilu — Б., 19406 : 9.
3.4.1 (2). Sprattus sprattus phalericus (Risso, 1826) — Черноморская килька (Св., 1952 : 107).
[1] русск. Килька — Г. Ник., 1954 : 116. Сардель Черное м.: одесск. — Б. : 158; н. т. Дуная — Гриц. Сердинка Черное м.: очаков. — Б. : 158. Средиземноморская килька — Св., 1952 : 107. Черноморская килька — Б., 1932 : 100. Черноморский шпрот — Г. Ник., 1954 : 116. Шпрот Черное м.: одесск. — Св., 1952 : 107. [2] укр. Сардель, Сарделька, Сердiнка, Чорноморська кiлька, Шпрот — Марк. : 61. [7] болг. Кайя — Пром. р. : 140. Копърка, Трицона, Цаца, Шпрот — Дренски : 47. [18] англ. Black sea sprat Канада — Ricker :
110. [24] исп. Espadin, Sardineta — Names of fishes : 37. [25] португ. Espadilha, Naval hinha — Names of fishes : 37. [26] рум. Ciacia, Tata — Виногр., 1960 : 38.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Европейский шпрот (Sprattus sprattus)
населяет моря Западной и Южной
Европы от Гибралтара до Лофотенских
островов (северный шпрот), Балтийское море
(балтийский шпрот, или килька),
северную часть Средиземного и Черного морей
(южноевропейский, или черноморский,
шпрот).
В Северном и Норвежском морях
северный шпрот (Sprattus sprattus sprattus)
держится ближе к берегу, чем сельдь, подходя
для нереста на глубины 20—40 м,
главным образом с апреля по июнь.
Промысловые скопления шпрота облавливаются
преимущественно в центральной и
северной частях Северного моря и у
берегов Южной Англии, Бельгии, Голландии
и Норвегии.
На втором году жизни северный шпрот
достигает 9—11,5 см длины и жирности
выше 7%. В это время он является
объектом интенсивного промысла.
Изготовляемые из шпрота консервы высоко
ценятся (частично шли под этикеткой
«сардины»).
Балтийский шпрот, или килька (Sprattus
sprattus balticus), встречается в
наибольшем количестве в бухтах юго-западных
берегов Балтийского моря и у входа
в Финский и Рижский заливы. Питается
планктонными рачками, главным образом
эвритеморой. На втором году жизни
достигает длины 7,5—11,2 см, на третьем —
10,6—14,1, на четвертом — 12,6—15,0 см,
накапливая от C,6) 4,1 до 15,2% жира.
Наиболее жирен бывает осенью и зимой,
наименее жирен в период нереста, с
апреля по июль. Половой зрелости достигает
обычно при длине 12 см, реже с 8,5—9 см
длины. Для нереста шпрот отходит от
берегов и выметывает свою плавучую икру
главным образом над глубинами 50—
100 м при солености от 4—5 до 17—18
промилле и температуре воды около
16—17° С. Балтийский шпрот, как и
салака, планктоноядная рыба, частично
конкурирует с ней из-за пищи. Балтийский
шпрот-килька — важная промысловая
рыба, дающая примерно от 10 до 20% всего
улова рыбы в Балтийском море.
Копченые шпроты очень вкусны. Консервы из
шпротов в масле пользуются заслуженной
популярностью. Точно так же хороша
соленая килька.
Черноморский шпрот (Sprattus
sprattus phalericus) наиболее многочислен в
Адриатическом и Черном морях, где его
промышляют Черноморский шпрот
придерживается обычно умеренно холодных слоев
воды, от 6—8 до 15—17°С, поднимаясь
зимой к поверхности, а в теплое время
предпочитая более прохладную воду на
глубине от 20—30 до 80—100 м. Он широко
распространен в открытом море, подходя
к берегам при ветрах, подгоняющих или
поднимающих массы воды
соответствующей температуры. Достигает половой
зрелости в возрасте одного года и
нерестится преимущественно в холодное время
(с октября по март) при температуре воды
7—10 (12)°С, частью в поверхностном слое,
главным образом, однако, на глубине
40—50 м. Черноморский шпрот достигает
длины 9,5—13 см, изредка бывает до
16 см; обычные размеры в уловах 6,5—11,5 см. Содержание жира в его теле
колеблется от 4,7 до 12,6%. Он не бывает
так жирен, как балтийская килька.
В Черном море шпрот—одна из самых
многочисленных рыб, играющая большую
роль в пище дельфинов, белуги, крупной
ставриды и других хищников. Но уловы
его относительно невелики, и попытки
развития промысла не увенчались
успехом вследствие того, что шпрот держится
чаще небольшими стаями, облов которых
на глубине затруднителен.
Фауна СССР. Рыбы. Том II, вып.1. Сельдевые (Clupeidae). А.Н.Световидов 1952 г.
*1. Sprattus sprattus (L.) — Шпрот.
Clupea sprattus Linne, Syst. nat., ed. X, 1758 : 318 (Habitat in oceano Europeo).
Сошник обычно без зубов, реже имеются зубы, есть слабые зубы на нёбных костях и на мезоптеригоиде, V I (5) 6 (7), общим числом обычно 7, изредка 6 и 8, А 17—21, sp. br. 42—59, на нижней половине жаберной дуги 30—41, vert. 46—50.
Морские рыбы, но заходят и в солоноватые воды, некоторые подвиды живут и размножаются в морях с пониженным содержанием солей.
Разделяются на 3 слабо различающихся подвида, каждый из которых распадается на несколько еще менее различающихся более низких систематических форм.
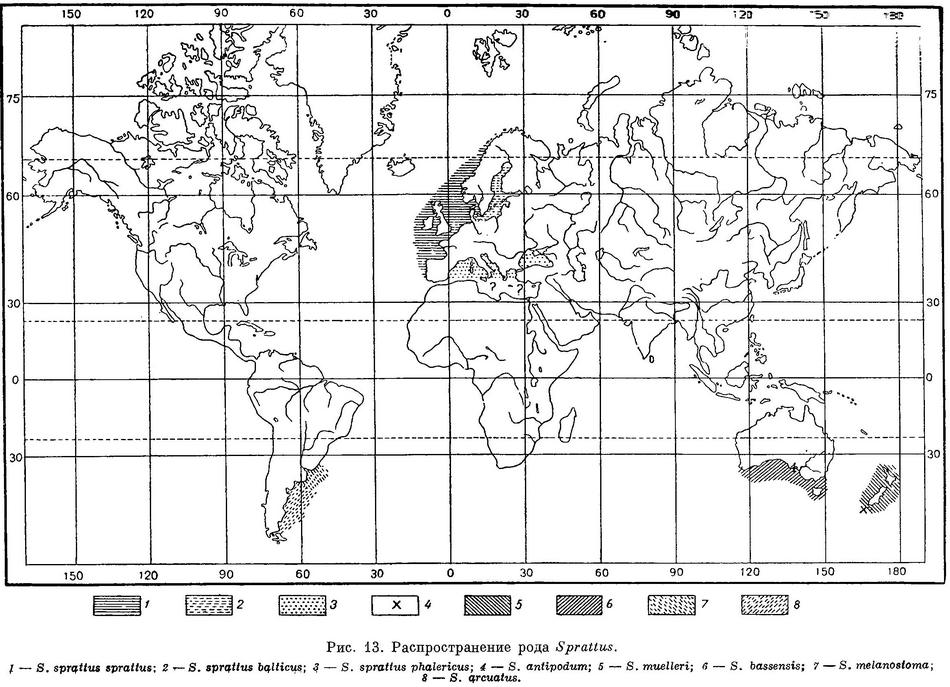
Распространение. Атлантический океан у берегов Европы от Гибралтарского прол. до Лофотенских о-вов, Балтийское море, европейские берега Средиземного и прилегающих к нему морей, Черное море (рис. 13).
 ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДВИДОВ
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДВИДОВ
1 (4). Общее число килевых чешуй в среднем не менее 33, за брюшными плавниками в среднем не менее 11.2. Голова широкая, межглазничный промежуток 16.5—21.5%, в среднем не менее 18.5% длины головы.
2 (3). Килевых чешуй позади брюшных плавников в среднем более 11.5. Атлантический океан у берегов Европы. Северное море. (Для Sprattus sprattus sprattus величины, приводимые в определительной таблице и во введении, получены по 30 экз. от берегов Испании (Виго).) Sprattus sprattus sprattus (L.)
3 (2). Килевых чешуй позади брюшных плавников в среднем менее 11.5. Балтийское море. *lb. Sprattus sprattus balticus (G. Schneider)
4 (1). Общее число килевых чешуй в среднем не более 33, за брюшными плавниками в среднем не более 11.3. Голова узкая, межглазничный промежуток 15.5—18.9%, в среднем около 16.5% длины головы. Средиземное, Адриатическое и Черное моря. * la. Sprattus sprattus phalericus (Risso)
 *la. Sprattus sprattus phalericus (Risso) — Черноморская (средиземноморская) килька, шпрот, сардель (у Одессы), сардинка (у Очакова) (табл. I).
*la. Sprattus sprattus phalericus (Risso) — Черноморская (средиземноморская) килька, шпрот, сардель (у Одессы), сардинка (у Очакова) (табл. I).
— Clupanodon phalerica Risso, Hist. nat. Europ. merid., III, 1826 : 452 (берега приморских Альп, залив у Ниццы).
— Clupea papalina Bonaparte, Cat. met. pesci Europ., 1846 : 34 (Адриатическое м.).
— Meletta phalerica Mоreau, Hist, nat. poiss. France, III, 1881 : 445, f. 196 (Средиземное м.: Ницца, Тулон, Сетт).
— Clupea phalerica Smitt, Scand. fish., II, 1896 : 976 (Генуя; 1 экз.).
— Clupea sulinae Antipa, Denkschr. Akad. Wiss. Wien, math.-naturw. Kl., LXXVIII, (1905), 1906 : 38, t. H, t. III, f. 1—6 (Черное м. перед Сулинским гирлом Дуная); Браунер, Тр. Бессарабск. общ. естеств. II, 2, 1912 : 127 (Бугаз у устья Днестровского лимана; 3 экз.).
— Clupea sprattus (Meletta phalerica) Fage, Rept. Danish Oceanogr. Exped. 1908—1910, 6, II (Biology), A 9, 1920 : 94 (Средиземное м. от Испании до Греции).
— Clupea papalina Fage, 1. с. : 94, 108 (Адриатическое м.).
— Clupea sulinae Fage, 1. c. : 96, 109 (Черное м. к северу от Босфора).
— Spratella serdinka Никольский, Бюлл. Гос. черном.-азовск. научно-пром. опытн. ст., 6—7, 1923 : 2 (Очаков); там же, 8—9, 1923 : 6, фиг.
— Clupea maeotica (non Grimm) Дренски, Тр. Българск. природоизпит. друж., XI, 1924 : 57 (обычен у берегов Болгарии).
— Spratella sprattus phalerica Дренски, Свед. землед., XII. 7—8, 1931 : 8 (Бургас, Месемврия, Варна); Берг, Докл. АН СССР, А, 1931 : 123 (сравнение с другими формами вида, синонимы).
— Sprattus sprattus phalericus Берг, Рыбы пресных вод, I, 1948 : 158; Казакова, в: Промысловые рыбы СССР, 1949 : 140.
Распространение, биология. Devedjian, Pcche et pecheries en Turquies, Constant., 1926 : 52 (у берегов Турции); Водяницкий, Раб. Новоросс. биолог. ст., I, 4, 1930 : 101 (Новороссийская бухта; икринки с половины ноября до апреля при температуре воды от 15 до 5°); Сыроватский, Зап. Гос. ихт. опытн. ст., IX, 21—22, 1930 : 49 (Березанский лиман); Попов, Докл. АН СССР, 9, 1930 : 211 (у берегов Крыма); Книпович, Тр. Азовско-черном. научно-пром. эксп., 5, 1932 : 361 (южная часть Азовского м.); Сыроватский, Докл. АН СССР, II, 3, 1934 : 199 (у о. Тендра и Одессы; нерест в июне); Водяняцкий, Тр. Севастоп. биолог. ст., V, 1936 : 1, 7, 35 (нерест в мае—июне); Пчелина, Тр. Новоросс. биолог. ст., 11, 1, 1936 : 27 (у Новороссийска; икринки); Клейненберг, Бюлл. Моск. общ. испыт. прир., отд. биолог., XLV, 5, 1936 : 349 (роль в питании дельфина); Докл. АН СССР, XV, 8, 1937 : 510 (Черное море против Ялты и северокавказского побережья; круглый год); Пилявская, Рыбн. хоз. СССР, 1937, 8 : 18 (миграции, возраст, уловы); Косякина, Тр. Новоросс. биолог. ст., II, 2, 1938 : 14 (у Новороссийска; икринки с начала ноября до середины мая, в массовом количестве в январе и феврале); Малятский, Природа, 1938, 5 : 96 (Черное м. между Очемчири и м. Пицунда на глубине 20—40 м, в галистатической области отсутствует); Fagе, Homage a oeuvre G. Antipa, Bucur., 1938 : 201 (верхний предел температуры размножения); Цалкин, Тр. Азовско-черном. научно-исслед. инст. морск. рыбн. хоз. и океаногр., 11, 1938 : 218 (роль в питании дельфина); Пчелина, Тр. Новоросс. биолог, ст., II, 3, 1940 : 52 (у Новороссийска; личинки в феврале—апреле); Клейненберг, Моск. общ. испыт. прир., юбил. изд., 1940 : 13 (роль в питании дельфина); Голенченко, Рыбн. хоз., 1940 : 6—14 (распределение в море против берегов Кавказа); Кротов, Докл. АН СССР, XXXIII, 2, 1941 : 162 (плодовитость); Голенченко, Рыбн. хоз., 1948, 4 : 5, рис. 2 (скопления); Смирнов, Докл. АН СССР, LXX, 1, 1950 : 129, рис. 1 (порционность икрометания); Алеев, там же, LXXXII, 1, 1952 : 161, рис. 1 (порционность икрометания, продолжительность нерестового периода); Павловская, там же : 165, рис. 1 (места и время икрометания).
D IV—V 11—14 (12.6), общим числом 16—18 (17.0), А II—IV 15— 19 (16.8), общим числом 17—21 (19.0), V I 6, sp. br. 49—59 (52.7), на нижней половине жаберной дуги 34—40 (36.8), килевых чешуй 20—22 + 10—12 (20.9 + 10.6), общим числом 30—34 (31.4), vert. S. 47—49 (47.50), caud. vert. 28—31 (29.45). Голова узкая и удлиненная, межглазничный промежуток 15.3—17.8 (16.5)% длины головы, длина головы 19.9—22.0 (21.0)% длины тела. Тело низкое, наибольшая высота 15.0—18.3 (16.0)% длины его, высота головы у затылка 12.8—14.9 (14.5)% той же длины. Спина сине-аспидного цвета, переходящего без резких границ в серо-серебристый цвет боков тела. Вершина рыла и нижней челюсти черноватые. Радужина серебристая с черными продолговатыми пятнами сверху. Размеры в Черном море наичаще 60—80 мм, наибольшие до 130, изредка до 160 мм. Описание дано по 30 экз. длиной от 89 до 112, в среднем 103.5 мм, из северо-западной части Черного моря у Очакова.
Морская форма, заходит в опресненные предустьевые пространства.
По данным А. А. Майоровой, у экземпляров из восточной половины Черного моря длиной 65—128 (90.8) мм sp. br. 46—62, позвонков 46—49 (47.7), килевых чешуй 29—33 (31.1); по ее исследованиям, шпроты из разных районов восточной половины Черного моря не различаются. По исследованиям Фажа (Fage, 1920), у экземпляров из Адриатического моря килевых чешуй 20—22 + 10—12 (21.0 + 10.3), общим числом 30—32 (31.3), позвонков 46—49 (47.5) (146 экз.); у экземпляров из Лионского зал. килевых чешуй 20—22 + 10—12 (21.3 + 11.3), общим числом 31—34 (32.7) (23 экз.)
Этот подвид, повидимому, распадается на несколько форм. В частности, у экземпляров из Адриатического моря антедорсальное расстояние меньше, чем у экземпляров из Черного моря, а количество килевых чешуй меньше, чем у экземпляров из Лионского зал. Идентичность этому подвиду Spatella serdinka Nikolsky установлена Бергом (1931). Неправильность мнения Фажа (Fage, 1920), ошибочно принявшего процентные отношения длины головы и антедорсального расстояния за абсолютные величины и считавшего поэтому Clupea sulinae Antipa особым видом, установлена также Бергом (1931).
Распространение. Европейские берега Средиземного моря и прилегающих к нему морей от Испании до Греции, наиболее обычен в Адриатическом море; Черное море. В Черном море повсеместно, но в наибольшем количестве временами у юго-восточных берегов Крыма, Кавказа в районе Поти—Новороссийска и в северо-западной части моря в районе Тилигульский берег — Аджиаск — Очаков. Заходит в Керченский прол. и прилегающие к нему части Азовского моря, в Днепровско-Бугский, Березанский, Днестровский лиманы, в теплое время года в Тилигульский, Григорьевский, Дофиновский и Сухой. Проникает единичными экземплярами, по всей вероятности, из моря при прорывах пересыпей, отделяющих лиман от моря. Нерест против всего советского побережья Черного моря, как у берегов, так и в отдалении до 60—80 миль (Здесь и далее — морские мили (1.85 км).), главным образом в северо-западной и юго-восточной частях моря, менее интенсивно у южного берега Крыма и в северовосточной части.
Биология. Икрометание происходит в течение почти всего года, с конца июля по май; начало его приходится на более ранние сроки и длительность больше в западной части моря, чем в восточной. В северозападной части моря икринки встречаются с конца июля по май, у берегов Крыма, к югу от Керченского прол. и у северных берегов Кавказа — с конца августа по май, у берегов Грузии (Сочи—Батуми) — с конца сентября по март. В начале икрометания, с конца июля по сентябрь в западной половине моря и с августа по ноябрь в восточной, нерестуют отдельные особи, на глубине 40—80 м при температуре воды 7—10°. В конце икрометания, в апреле—мае, на этих же глубинах встречаются единичные икринки. В северо-западной части моря единичные икринки обнаружены в конце июля на глубинах не менее 15—20 м при температуре воды 8—9°. Интенсивный нерест здесь начинается с конца октября, в наибольшем количестве икринки найдены в опресненной части, примыкающей к Днестровскому лиману. Против южного берега Крыма, к югу от Керченского прол. и у северных берегов Кавказа единичные икринки найдены в конце сентября на глубине 45—80 м при температуре воды 6—8°. В поверхностных слоях при температуре 20—23° икринок здесь не было; они обнаружены в поверхностных слоях в конце сентября при температуре воды 16.5°. В период массового нереста шпрот держится как у берегов, так и в отдалении от них (до 60—80 миль), как у поверхности, так и до глубины 100 м, не образуя больших промысловых концентраций, а встречаясь в виде небольших разреженных косяков. У берегов Грузии единичные икринки обнаружены в конце сентября на глубине 25—50 м при температуре воды 9.5—13°. В апреле здесь встречались только личинки и мальки, обнаруженные в этом районе в наибольшем количестве (Павловская, 1952). У берегов Румынии икрометание длится до июля (ст. ст.) (Antipa, 1906), в Средиземном море и в прилегающих морях нерестует с декабря по апрель, в северной части Адриатического моря при температуре 8—10° и солености не ниже 35‰ (Fage, 1920). Плодовитость от 575 до 2398 икринок (Пилявская, 1937), по другим указаниям (Кротов, 1941), — от 1180 до 2416, в среднем 1398 икринок (длина рыб 50—75 мм). Икрометание порционное, икра выметывается в три порции, причем большая длительность нерестового периода обусловлена, судя по всему, не растянутостью нереста отдельных особей (Смирнов, 1950), а неодновременностью их созревания (Алеев, 1952). Личинки выклевываются через 4—5 дней по оплодотворении (Водяницкий, 1930). С повышением температуры воды молодь от берегов, повидимому, отходит, так как в теплое время года мальков-сеголетков в Новороссийской бухте не найдено (Водяницкий, 1936; Косякина, 1938; Пчелина, 1940). Наибольшее количество мальков, по наблюдениям А. А. Майоровой, встречалось на расстоянии от 3 до 5 миль от берега, в наибольшей концентрации в районе Абхазии и Анакрии. В Средиземном море встречается в незначительном количестве, наиболее обычен в глубине заливов, перед устьями в наибольшем количестве встречается в Адриатическом море. У берегов северной Африки отсутствует по причине высоких температур воды, близких даже в зимнее время (декабрь—февраль, в среднем 14°) к верхнему пределу температурных условий нереста (Fage, 1920). В открытых частях Черного моря, повидимому, держится в значительном количестве круглый год, образуя временами в отдельных местах более значительные концентрации, но в галистатической области, по наблюдениям С. М. Малятского, отсутствует. В восточной части Черного моря образует скопления весной, в марте—мае, в 30—40 милях против берегов Кавказа между Туапсе и Новороссийском. Позднее, в мае—июне, образует скопления в 20—30 милях против берегов Крыма между м. Сарыч и м. Меганом, которое держится на одном месте около 3 месяцев, исчезая в августе—сентябре. Осенью, в сентябре—ноябре, образуется скопление против берегов Кавказа на тех же местах, что и весной. (По сообщению Н. Аслановой, Черноморской экспедицией ВНИРО такие постоянные скопления в открытых частях Черного моря не наблюдались. Повидимому, в нерестовое время держится разреженно на большом пространстве, как об этом свидетельствует и Павловская (1952).) Эти скопления сопровождаются косяками дельфинов, усиленно питающимися шпротами (Голенченко, 1940; Цалкин, 1938; Клейненберг, 1936, 1940). В летнее время против кавказских берегов в открытом море (в 80 милях от берега) держится мелкий шпрот длиной около 30 мм, ближе к берегу (в 15 милях) — более крупный, длиной около 50 мм; в прибрежной зоне над глубинами 20—30 м встречается шпрот длиной от 50 до 70 мм и крупнее. После нереста производители подходят к берегам, молодь же первый год жизни, по наблюдениям С. М. Малятского, проводит в открытом море, отходя от берегов в теплое время года (Пчелина, 1940). В прибрежную область в промысловом количестве у анатолийских берегов вместе с хамсой ловится с декабря по март и с мая по конец сентября вместе с атеринкой (Devedjian, 1926). К кавказским берегам подходит в ноябре—декабре и держится в течение всей зимы до мая—июня. В районе Адлера вылавливается вместе с хамсой, с декабря по апрель, составляя, по наблюдениям А. А. Майоровой, в большинстве случаев преобладающую массу улова. По ее же наблюдениям, в большом количестве зашел в Азовское море в 1930 г., ловясь в июне—августе вдоль северного побережья Керченского прол. (Мама, Юргаков кут), а также в 1925 г. (Книпович, 1932). Единичными экземплярами был обнаружен весной 1935 г. в Керченском прол. (Пилявская, 1937). В северо-западной части Черного моря в промысловом количестве подходит к берегам с апреля—мая и ловится до сентября—октября, наибольшие уловы бывают в июле, в некоторые годы в июне и августе, иногда в мае. В годы слабого подхода младших возрастных групп, (двухгодовалых) наибольшие уловы бывают в мае и в июне. В годы массового подхода мелкой кильки наибольшие уловы бывают в июле и августе. Вначале подходят старшие возрастные группы, в конце апреля, в мае и июне главным образом трехгодовалые, частично четырехгодовалые, в июле—августе — двухгодовалые. Косяки обычно состоят в основном из одной возрастной группы, иногда с небольшой примесью рыб другого возраста. Подходят главным образом в районе Тилигульский берег — Аджиаск — Очаков, к о. Тендру обычно подходит незначительное количество (Пилявская, 1937). К берегам Румынии в массе подходит в июне и октябре (Antipa, 1906). У берегов Болгарии обычен летом (Дренски, 1931). К берегам в наибольшем количестве подходит при температуре воды 15—20°, при 11—12° уловов нет или они незначительны, точно так же как и при повышении температуры воды до 25° (Пилявская, 1937). В северо-западной части Черного моря наибольшие уловы совпадают с северо-западными и восточными ветрами. Молодь здесь в массе подходит к берегам в июле—августе, когда температура воды здесь наиболее высока (Пилявская, 1937). Размеры шпрота, ловимого у берегов Кавказа, колебались от 35 до 110 мм, наичаще 55—80 мм, причем у Сухуми, по наблюдениям А. А. Майоровой, шпрот ловится более крупный, чем у Батуми. В северо-западной части Черного моря ловится шпрот несколько более крупный, чем в восточной половине моря, размеры его от 35 до 130 мм, средние размеры колеблются от 61 до 80 мм, предельный возраст 5 лет. Рост, по данным Пилявской (1937), происходит следующим образом:
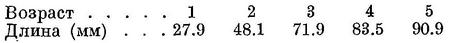
Содержание жира сравнительно не велико, в северо-западной части Черного моря около 2.34%.
Хозяйственное значение в Черном море в настоящее время небольшое, но уловы могут быть во много раз увеличены, так как имеющиеся большие запасы не используются. Уловы в Черном море в 1937—1938 гг. составляли 13.2—34.7 тыс. ц, из них в северо-западной части моря добыто 11.2—31.6 тыс. ц, у берегов Крыма и у входа в Керченский прол. 0.8, у берегов Кавказа 1.2—2 тыс. ц. В Средиземном море и прилегающих к нему морях, кроме Адриатического, промыслового значения не имеет. Ловится у берегов совместно с другой рыбой в северозападной части моря с апреля по август, отчасти до октября, у берегов Крыма — в июне—июле, у берегов Кавказа — в декабре, затем, в апреле—мае.
*lb. Sprattus sprattus balticus (G. Schneider) — Балтийская килька, шпрот (табл. I).
— Clupea sprattus Linnaeus, Fauna Svecica, ed. II, 1761 : 128 (Habitat in mari); Pallas, Zoogr. rosso-asiat., III, 1811 : 214 (Ревель, Балтийский порт).
— Meletta vulgaris Кeсслер, Рыбы СПб. губ., 1864 : 190 (Финский зал., на восток до устья Наровы, по указанию рыбаков — до Кронштадта).
— Clupea sprattus Smitt, Scand. fish., II, 1895 : 974, pl. XLIV, f. 2 (частью: Балтийское м., Финский и Ботнический заливы); Heincke, Naturgesch. d. Herings, I, 1898 : LXIX, 177—185, 202—203, Taf. VII, f. 3, 5, 6, Taf. XX, f. 1, 2 (Киль, Гела, Стокгольм; измерения).
— Clupea sprattus balticus G. Schneider, Korrespondenzbl. Naturf.-Ver. Riga, XLVII, 1904 : 66 (nomen).
— Clupea sprattus var. baltica G. Schneider, Rapp. Proces-Verb., IX, 1908 : 94, 96 и след. (Балтийское м.; измерения, нерест, рост).
— Spratella sprattus baltica Берг, Рыбы пресных вод, I, 1932 : 99.
— Sprattus sprattus balticus Берг, Рыбы пресных вод, I, 1948 : 157; Телегин, в : Промысловые рыбы СССР, 1949 : 137, табл. 34.
Распространение, биология. Benecke, Fische, Fischer, u. Kischz. Ostpreuss., 1881 : 172 (мертвые после нереста); Möbius u. Heincke, Fische d. Ostsee, 1883 : 139 (Балтийское м.); Hinkelmann, Mitt. Deutsch. Seefischerei-Ver., XVI, 6, 1899 : 127 (Кильский канал); Schneider u. Levandеr, Acta Soc. Fauna Flora fennica, XX, 1, 1900 : 9, 46 (питание, нерест и развитие икры); G. Schneider, Zool. Anz., XXV, 659, 1901 : 9 (Финский зал.; нерест); Acta Soc. Fauna Flora fennica, 22, 4, 1901 : 50 (время нереста); ibid., 22, 2, 1902 : 50 (паразиты); Jenkins, Wiss. Meeresunters. (N. F.), Abt. Kiel, VI, 1902 : 35 (Кильская бухта, Кильский канал; рост); Ehrenbaum u. Strodmann, Wissensch. Meeresunters. (N. F.), Abt. Helgol., VI, 1904 : 51—126 (диаметр икринок); Strodmann, ibid., VII, 2, 1906 : 133—216 (биология); Mela, Vert. Fennica, 1909 : 478 (Ботнический зал., в северной части); Суворов, Тр. Балтийск, эксп., I, 1910 : 91, 94, 97, 98, 99, 100, 101, 107 (Финский и Рижский заливы; биология); там же, II, 1913 : 54, 59—60, 62, 96—97, табл. X (измерения, нерест, питание); Нeidrich, Wissensch. Meeresunters. (N. F.), Abt. Kiel, XX, 1, 1923—1925 : 3—38 (Кильская бухта; размножение); Михин и Антипова, Изв. Всесоюзн. Научно-исслед. инст. озерн. и речн. рыбн. хоз., XV, 1932 : 54 (Нарвская губа: Шепелево); Dixon, Rapp. Proces-Verb., CII, Append., 1933 : 1—12 (Данцигская бухта; уловы, возрастной состав, рост, жирность); Robertson, Fish. Invest., ser. II, XVI, 2, 1938 : 3—103 (сводка наблюдений и литературных данных для вида в целом); Нöglund, Svensk. Hydrogr.-Biol. Komiss. Skrift. (N. S.), II, 3, 1938 : 3 (Скагеррак и Каттегат; распределение икринок); Molander, ibid., II, 4, 1940 : 5 (западвые берега Швеции); Берг, Изв. Всесоюзн. Научно-исслед. инст. озерн. и речн. рыбн. хоз., XXIII, 1940 : 6 (Финский зал., на восток до Бьёркэ и Шепелева; рост, питание); Дементьева, Рыбн. хоз., 1947, 8 : 27 (нерест); Борисов, там же, 1948, 8 : 31 (берега Эстонии и Латвии; возрастной состав); Канд и Горбачев, Рыбн. хоз., 1950, 5 : 39 (содержание жира); Дрягин, Вестн. Ленингр. унив., 8, 1950 : 27 (улов в советских водах Финского зал. 1 тыс. ц, уловы в водах Эстонии могут быть увеличены в 3 раза).
D III—V 12—16 (13.6), общим числом 15—19 (17.1), А II—III 14—18 (16.1), общим числом 17—21 (18.3), V I 6 (Гейнке указывает V I 5 (1898 : 178, 179, 183) и один случай V I 7 (1898 : 185).), sp. br. 42—55 (50.0), на нижней половине жаберной дуги 30—41 (34.9), килевых чешуй 20—23 + 10—13 (22.0 + 11.4), общим числом 31—35 (33.3), vert. S. 47—50 (48.06), caud. vert. 29—32 (31.01). Голова широкая и укороченная, межглазничный промежуток 16.7—21.3 (18.9)% длины головы, длина головы 18.4—20.6 (19.6)% длины тела. Тело сравнительно высокое, наибольшая высота 13.2—19.6 (17.6)%, высота головы у затылка 12.8—14.8 (13.7)% длины его. Спина темноголубая, бока и брюхо блестяще-серебристые. Размеры наичаще 11—13 см, наибольшие до 15—17 см. Описание дано по 50 экз. длиной от 10.1 до 14.0, в среднем 12.1 см, собранным в марте 1941 г. в Финском зал. между Койвисто и Кургаловым.
По данным Гейнке (Heincke, 1898), у шпрота из Кильской бухты, от Гела и от Стокгольма килевых чешуй 20—24 + 9—13 с колебаниями средних от 21.7 до 22.3 и от 11.2 до 11.5, позвонков 45—49 с колебаниями средних от 47.5 до 48.1.
Морская форма, держащаяся и размножающаяся в воде как с почти нормальным, так и с пониженным содержанием солей.
По исследованиям Гейнке, распадается на несколько форм, незначительно различающихся некоторыми систематическими и биологическими особенностями. У экземпляров из восточной части моря икринки крупнее, количество их меньше и развиваются они в менее соленой воде, чем у экземпляров из западной части моря и из Скагеррака. По скорости роста и количеству позвонков в Каттегате и Скагерраке различают (Molander, 1940) шпрота трех типов: морского — наиболее быстрого роста и с наибольшим количеством позвонков (в среднем от 47.9 до 48.1), фиордового — нахтболее медленного роста и с наименьшим количеством позвонков (в среднем от 47.6 до 47.7) и открытых фиордов (skargird) — с ростом и количеством позвонков (в среднем от 47.7 до 47.9), промежуточным между первыми двумя типами. Обособленности нерестилищ этих типов шпротов не наблюдается; эти типы рассматриваются как результат развития и роста в различных условиях.
Распространение. Балтийское море, где встречается почти повсеместно, но в наибольшем количестве у входа в Финский зал. в районе Таллина, Хельсинки и Ханко, в Рижском зал. у входа и у западных берегов его, у островов Саарема и Хиума, у берегов Пруссии, в Мекленбургской и Кильской бухтах и ряде других мест. В Финский зал. на восток по северному берегу заходит до Бьёркэ, по южному до Шепелева, отмечался у Кронштадта (По указанию рыбаков (Кесслер, 1864).). В Ботническом зал. встречается повсюду, до 64—65° с. ш. На запад — до Кильской бухты и, возможно, южной части Каттегата (у Гётеборга, судя по всему, типичная форма вида). Нерест, повидимому, по всему морю, на север до Аландских о-вов, в Рижском зал., в Финском зал. на восток до Хельсинки и Таллина, в северной части Каттегата и в восточной Скагеррака, в фиордах этих проливов, как правило, за некоторыми исключениями (фиорд Уддевалла) нереста нет.
Биология. Икрометание происходит весной и летом (Гейнке (Möbius u. Heincke, 1883) указывает на нерест в Кильской бухте весной и осенью, считая весенне- и осенненерестующего шпрота разными расами. Осенний нерест шпрота (в сентябре) отмечает также Бенеке (Benecke, 1881), сообщая одновременно о гибели после нереста в Данцигском зал. у Путцига.), в западной части ареала с конца апреля по конец августа, к востоку от о. Борнгольм с начала мая по начало августа включительно, к востоку от о. Готланд и в Рижском зал. с апреля—мая по начало августа (Schneider, 1908; Дементьева, 1947). Нерестится как в открытом море, так и в заливах, но не у самых берегов и не в фиордах, а на некотором расстоянии от них, в Каттегате и в Скагерраке главным образом над глубинами 50—100 м (Molander, 1940). В открытом море икринки во второй половине июня 1947 г. были обнаружены на всем пространстве от Данцигской бухты до Ирбинского прол. (Борисов, 1948). Икринки в воде как почти с нормальным содержанием солей (в восточной части Скагеррака), так и с пониженным (6—8‰ у о. Борнгольм, в Финском зал. даже 4—5‰) держатся в поверхностных слоях воды или на некоторой, главным образом 25—50 м, глубине, что обусловлено более крупными размерами их и меньшим удельным весом в восточной части ареала, чем в западной. Диаметр икринок в Финском зал. равен 1.1—1.7 мм, у о. Борнгольм 1.25—1.48 мм, у типичной формы вида в Северном море у о. Гельголанд лишь 0.83—1.13 мм (Schneider, 1908). Икра откладывается несколькими, но наблюдениям в Кильской бухте (Heidrich, 1923), 8—9 порциями с интервалами в 8—10 дней и весь процесс икрометания каждой самки длится около 2.5—3 месяцев. В каждой порции соответственно возрасту (одно-, двух- и трехгодовалые) выметывается в среднем 1230, 2760 и 3560 икринок. Общая плодовитость соответственно тем же возрастам исчисляется приблизительно в 11, 27 и 36 тыс. икринок. Развитие икринок длится 3—4 дня (Hoglund, 1938). Взрослые по окончании нереста подходят к берегам и держатся здесь с сентября до весны (Schneider, 1908), у Таллина на короткое время появляется в мае (Суворов, 1913). Заходит в шхеры, фиорды и бассейны, временами отделяющиеся от моря; в Нильском канале ловилась вместе с сельдью, камбалой и пресноводными рыбами — плотвой, окунем (Hinkelman, 1899). Питается ракообразными — Calanidae, в Нарвской губе главным образом Erytemora affinis var. hirundoides и его яйцами (Берг, 1940). Самки численно, повидимому, преобладают над самцами, количество их колеблется от 55.5 до 73.5%, количество самцов — от 26.5 до 44.5% (Jenkins, 1902; (Möbius u. Heincke, 1883; Schneider, 1908). Половой зрелости достигает при длине, начиная с 12 см, в двух- трехгодовалом возрасте, шпрот Каттегата — на втором году жизни, реже в возрасте одного года при длине 8.5—9 см. В уловах в западной части ареала (у берегов Швеции) встречаются двух- и трехгодовалые, четырехгодовалые редки, в Нарвской губе — трех- и четырехгодовалые (Jenkins, 1902; Molander, 1940; Берг, 1940). Весной у северозападных берегов Рижского зал. (1946 г.) и юго-западных Финского (1947 г.) в уловах преобладали двухгодовалые (93.7—95.5%), в незначительном количестве (4.5—6.3%) встречались трехгодовалые (Борисов, 1948). В Скагерраке и Каттегате шпрот урожайных годов мельче шпрота менее урожайных лет. Основной возрастной группой в уловах Скагеррака и Каттегата являются одногодовалые, двухгодовалые встречаются в небольшом количестве, трехгодовалые — единицами процентов и не каждый год, причем процент двух- и трехгодовалых бывает тем больше, чем меньшие размеры имеют двухгодовалые поколения этого года. В самих проливах процент старших возрастных групп выше (Molander, 1940). Рост в Финском зал. (Нарвская губа; Берг, 1940), Кильской бухте (Jenkins, 1902) и в Каттегате и его фиордах (Molander, 1940) происходит следующим образом (абсол. длина, в см):
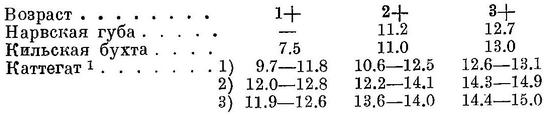
1 Колебания средних шпрота 1) фиордового типа, 2) типа открытых фиордов и 3) морского типа.
Содержание жира у шпрота Каттегата (фиорд Уддевалла) колеблется от 4.11 до 15.19%, наименее жирен в период нереста — в апреле—июле (3.30—7.40%), в августе содержание жира повышается (до 9.30%), наиболее жирен бывает в сентябре—январе (10.62—15.19%) (Molander, 1940). В Данцигской бухте наибольшее количество жира содержит в декабре—январе (14.2%), наименьшее — в июне (3.6%) (Dixon, 1933), в Финском зал. наименьшее количество — в мае—августе (6.5—8.1%), наибольшее — в сентябре—феврале (10.6—13.7%) (Канд и Горбачев, 1950).
Хозяйственное значение. Важная промысловая рыба Балтийского моря. Общий улов не превышает, повидимому, 250 тыс. ц. Большая часть его добывается в западной части моря: в 1936—1939 гг. у берегов Польши поймано 104—151 тыс. ц, Германии — 5.5—70 тыс. ц. В восточной части моря у берегов Финляндии добывалось 4—7 тыс. ц, Швеции — около 1.5 тыс. ц, Эстонии — 9—43 тыс. ц, Латвии — 6—10 тыс. ц, Литвы — до 0.4 тыс. ц. У берегов РСФСР ловится в небольшом количестве в Нарвской губе Финского зал. Уловы могут быть увеличены в открытом море. Ловится главным образом осенью, в сентябре—ноябре, и отчасти весной и летом.
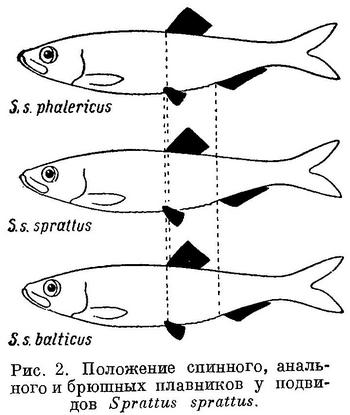
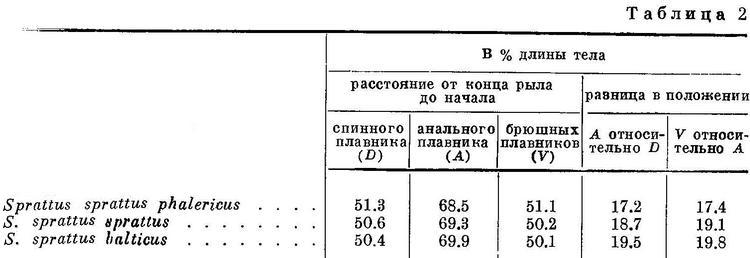
Соответственно положению спинного плавника изменяется положение анального и брюшных плавников и у подвидов в пределах некоторых видов подсем. Clupeinae. Естественно, что степень различия рассматриваемых признаков между подвидами менее степени различий их у видов.
Положение анального и брюшных плавников относительно переднего конца тела, точно так же как первого относительно спинного и второго относительно анального у подвидов Sprattus sprattus соответственно положению спинного плавника относительно переднего конца тела, изменяется таким же образом, как и у родов и у видов подсем. Clupeinae (табл. 2, рис. 2).
Рыбы пресных вод СССР и сопредельных стран. Л.С.Берг 1948 г.
Sprattus sprattus balticus (Schneider). — Балтийская килька, шпрот
(Финны — kilo, эсты — kilu, шведы — vassbuk)
Clupea sprattus Pallas, Zoogr. rosso-asiat., III, 1811, p. 214 (Ревель, Балтийский порт).
Meletta vulgaris Кесслер, Рыбы СПб. губ., 1864, стр. 190 (доходит до устья Наровы; по словам рыбаков, бывает и у Кронштадта; 100—125 мм, брюшных шипов 31, позвонков 47).
Clupea sprattus Smitt, Scandin. fishes, II, 1895, p. 974, pi. XLIV, f. 2. — Heincke, Naturgeschichte des Herings, 1898, p. LXIX, tab. VII, fig. 3, 5, 6; tab. XX, fig. 1, 2, p. 177—185 (Киль, Гела, Стокгольм), p. 202—203.
Clupea sprattus var. baltica Gu. Schneider, Korr.-Bl. Naturf.-Ver. Riga, XLVII, 1904, p. 41—45; Rapp. et proc.-verb. Conseil intern, pour l'explor. de la mer, IX, 1908, p. 96 (Балтийское море).
Clupea sprattus Михин и Антипова, Изв. Инст. озерн. и речн. рыбн. хоз., XV, 1932, стр. 54 (Нарвская губа — Шепелево).
Spratella sprattus baltica Берг, Рыбы пресн. вод СССР, I, 1932, стр. 99; Изв. Инст. озерн. и речн. рыбн. хоз., XXIII, 1940, стр. 9.
D IV—V 12—14 (всего 17—19), A III—V 15—17(всего 18—22), V I 6 (Как исключение на одной стороне бывает I 5 и I 7 (см. Heincke, p. 183, 185).), squ. 47—50, брюшных шипов впереди брюшных плавников 20—24 (обычно 22—23), позади 9—13 (обычно 11), жаберных тычинок на 1-й дуге 46—51. Позвонков 45—49, наичаще 47—48. Балтийская килька отличается от кильки Северного моря более низкой головой, более длинным рылом, несколько меньшим числом брюшных шипов, которых у балтийской за V в среднем 11.2—11.5 (у атлантической 11.5—11.8). У экземпляров из Нарвского залива длиной 113—126 мм (абс.) наибольшая высота тела в длине всего тела 17.4—19.5, длина головы 18.7—19.8, антедорсальное расстояние 45.0—46.5, наименьшая высота тела 6.1—7.3. У балтийской кильки несколько больший диаметр икринок: в Финском заливе диаметр 1.1—1.7 мм, тогда как у Гельголанда всего 0.88—1.13 мм. Поэтому даже в слабосоленой воде Финского залива икринки не опускаются на дно, а развиваются пелагически. В Финском заливе нерест вдали от берегов, с конца мая по начало августа. Длина (абс.) в Финском заливе до 150—160 мм, обычно меньше: 106 экз., добытых В. С. Михиным в Нарвском заливе (д. Конново) 5 декабря 1929 г., имели от 107 до 133 мм, в среднем 117 мм (абс.). У д. Конново и у Кургалова осенью составляет предмет промысла. В. С. Михин доставил мне 1 экз. кильки длиною 113 мм (абс.), добытый 2 марта 1930 г. у Шепелева (западнее Красной Горки). Есть у островов Лавансари, Гогланда и Коивисто (Бьёркё).
Килька очень похожа на мелкую сельдь (салаку), но отличается от нее меньшей длиной тела, меньшим числом поперечных рядов чешуй, жаберных тычинок, позвонков, брюшных шипов, пилорических придатков, большим числом лучей в анальном плавнике, положением брюшного плавника (под началом D или даже впереди его), более короткой и более высокой головой, резко выдающимися брюшными шипами, более короткой нижней челюстью, хватающей кзади обычно только до вертикали переднего края глаза или чуть дальше (у сельди до вертикали передней трети глаза), отсутствием зубов на сошнике, свободно плавающими (пелагическими) яйцами (у сельди икра, прилипающая к субстрату).
Sprattus sprattus phalericus (Risso). — Черноморская килька
(В Очакове — сердинка, в Одесском районе — сардель.)
Clupanodon phalerica Risso, Hist. nat. de l'Europe merid., III, Paris, 1826, p. 452 (cotes des Alpes-Maritimes, golfe de Nice).
Clupea sulinae Antipa, Denkschr. Akad. Wien, math.-naturw. Kl., LXXVIII, 1906, p. 38, tab. III, fig. 1—6 (Черное море перед Сулинским устьем Дуная, D 16—17, А 19—20, squ. 42—46, брюшных чешуй 29—33, длина до 100 мм). — Браунер, Тр. Бессарабск. общ. ест., II, вып. 2, 1912, стр. 13 (Бугаз у устья Днестровского лимана).
Clupea sprattus (Meletta phalerica) Fage, Report Danish Oceanogr. Exp. 1908—1910, № 6, (vol. II, Biology, A. 9), Copenhagen, 1920, p. 94 (Средиземное море: от Испании до Греции), р. 101 (икрометание январь—март), р. 108 (Адриатическое море), р. 109 (Лионский залив).
Clupea sulinae Fage, 1. c., p. 109 (Черное море несколько севернее Босфора).
Spratella serdinka А. Никольский, Бюлл. Черном.-азовск. ст., Очаков, № 6—7, 1923, стр. 2 (Очаков); № 8—9, 1923, стр. 6, фиг.
Spratella sprattus phalerica Берг, Доклады Акад. Наук, А, 1931, стр. 123 (Очаков); Рыбы пресн. вод СССР, I, 1932, стр. 100; II, 1933, стр. 837. — Пилявская, Рыбн. хоз., 1937, № 8, стр. 18 (сев.-зап. часть Черн. м.; улов в 1934 г. 25 тыс. ц). — Косякина, Тр. Новоросс. биол. ст., II, вып. 2, 1938, стр. 14 (икра у Новороссийска с начала XI до середины V). — Голенченко, Рыбн. хоз., 1940, № 6, стр. 14.
D 16—17, А 19—20, V I 6, squ. 42—46, брюшных шипов впереди брюшных плавников 19—22, позади 9—12 (всего 29—33) в среднем 31.1 (У адриатических в среднем 31.3, у килек из Лионского залива 32.7.), жаберных тычинок на 1-й дуге 45—56. Позвонков (46) 47—48 (49) (У 146 адриатических 46—49, в среднем 47.5; у 26 черноморских 46—49, в среднем 47.4, т. е. одинаковое (Fage). У атлантических 47.5—47.7.). Подвид этот в Средиземном, Адриатическом и Черном морях заменяет атлантическую кильку (Sprattus sprattus), к которой он весьма близок, отличаясь несколько более вытянутым телом, более длинной головой, несколько более выдвинутым вперед спинным плавником, более крупной чешуей, несколько меньшим (в среднем) числом брюшных шипов: 31.1—32.7, тогда как у атлантических 33.3—33.6. У экземпляров из Очакова длиной (абс.) 90—112 мм, в процентах длины всего тела: наибольшая высота тела 16.5—17.9, длина головы 20.5—21.9, антедорсальное расстояние 44.6—47.9. Длина (абс.) в среднем 95—110 мм, максимум до 120 мм.
Черное море, повсеместно: перед устьями Дуная (июль), Днестра (август), в лиманах Тилигульском и Березанском (Западнее Очакова (И. Я. Сыроватский, Зап. Ихтиол. ст. Херсон, вып. 21—22, 1930, стр. 49). Относительно Тилигульского лимана и соседних см. у А. Пилявской. — Есть и у Бургаса (Болгария; Дрънски, 1931).), в Днепровско-бугском лимане (ловится у Очакова с мая до осени, бывает здесь и в то время, когда вода совершенно пресная), зимой появляется у Севастополя (В Зоологическом институте Академии Наук есть много экземпляров из Севастополя длиною (абс.) 80—92 мм, добытых С. А. Зерновым 2 марта 1922 г. н. ст. (№ 21050).), по кавказскому берегу составляет предмет промысла. По свидетельству Н. М. Книповича, в 1925 г. зашла в большом количестве в Азовское море (северное побережье Керченского полуострова). Икра этого вида встречается поздней осенью, зимою и весною у Новороссийска (Водяницкий, Работы Новороссийск, биол. ст., IV, 1930, стр. 101, фиг. 2. — Косякина, 1938. Следует отметить, что мальки кильки вылупляются из икры через 4—5 дней по оплодотворении.). С. М. Малятский обнаружил в конце мая и начале июня 1939 г. молодь кильки по всему Черному морю, в открытой части. Нерест с ноября по июнь.