(Scombridae Rafinesque, 1815) Семейство Скомбриды, Скумбриевые, Макрелевые, Family Scombridae Rafinesque, 1815 (Mackerels, Tunas, Bonitos) 15 родов 51 вид
Информация из Интернета
Семейство Scombridae впервые описал в 1815 году американский учёный Константэн Рафинеск (Константэн Самюэль (Константайн Сэмюел) Рафинеск (Рафинеск-Шмальц) — Constantine Samuel Rafinesque-Schmaltz, 1783 — 1840, американский натуралист, зоолог и ботаник, работал во многих других областях науки, в том числе в малакологии, метеорологии, антропологии, геологии, лингвистике, он был в основном самоучкой, никогда не посещал университет, в 1808 году избран членом Американской академии искусств и наук (American Academy of Arts and Sciences), совершил множество научных экспедиций по Северной Америке, в 1819 году стал профессором ботаники в Университете Трансильвании (Transylvania University) в Лексингтоне (Lexington), затем профессором естественной истории в Институте Франклина (The Franklin Institute) в Филадельфии, открыл и описал (по его собственным словам) не менее 250 видов растений и животных, а также несколько высших таксонов и 65 новых видов рыб, в его честь названо 2 вида рыб).
Скомбриды, Скумбриевые, Макрелевые (Scombridae) — семейство лучепёрых рыб из отряда окунеобразных (Perciformes Bleeker, 1859), обитающих в водах тропического, субтропического и умеренного районов Атлантического, Индийского и Тихого океанов и в прилегающих морях, некоторые виды встречаются также в солоноватой и даже в пресной воде. В данном семействе, согласно недавним филогенетическим исследованиям, выделяют два подсемейства: Gasterochismatinae Poey, 1869 (монотипический род, один вид — Gasterochisma melampus Richardson, 1845) и Scombrinae Rafinesque, 1815 (Четырнадцать родов и 50 видов).
Много литературных изданий посвящены этому подсемейству, но, прежде всего, следует отметить публикации Б. Коллетта с соавторами (Collette et al., 2001), на которых основано наше современное знание о разнообразии и систематике этой важной промысловой группы рыб и приведенная ниже классификация (см. кладограмму). Кладограмма представляет собой инструмент для визуализации и сравнения эволюционных отношений между таксонами и читается слева направо, как временная шкала. Данная кладограмма иллюстрирует взаимоотношения между тунцами и прочими трибами семейства скумбриевых. Например, видно, что полосатые тунцы более близкородственны Thunnus, по сравнению с южными тунцами (наиболее примитивные из тунцов), а следующей наиболее близкородственной группой являются пеламиды.
Подсемейство Gasterochismatinae Poey, 1869 (гастерохизматины). Чешуя среднего размера, примерно 80 в боковом ряду; у неполовозрелых — брюшной плавник длиннее, чем голова, укладывается в глубокую брюшную борозду. Один аберрантный (отклоняющийся от нормального строения, расположения или состояния) вид — Gasterochisma melampus Richardson, 1845, главным образом в Южном океане.
Подсемейство Scombrinae Rafinesque, 1815 (скомбрины) (см. схему). В настоящее время разделено на четыре трибы, только триба Scombrini подтверждена молекулярными данными, для трёх других — необходимы дополнительные исследования. Чешуя крошечная или отсутствует.
• Триба Scombrini (mackerels — макрели). Два рода: Rastrelliger Jordan & Starks, 1908 и Scomber Linnaeus, 1758 (7 видов). Как и в случае со многими другими обиходными названиями, название «макрель» (mackerel) используется для видов из многих разных семейств.
• Триба Scomberomorini (spanish mackerels — испанские макрели). Три рода (21 вид): Acanthocybium Gill, 1862, Grammatorcynus Gill, 1862 и Scomberomorus Lacepède, 1801. Несмотря на то, что Scomberomorus sinensis (Lacepède, 1800) морской вид, он встречается и в эстуариях и известен на расстоянии до 300 км вверх по Меконгу.
• Триба Sardini (bonitos — бонито). Четыре рода (7 видов): Cybiosarda Whitley, 1935, Gymnosarda Gill, 1862, Orcynopsis Gill, 1862 и Sarda Cuvier, 1829.
• Триба Thunnini (tunas — тунцы) (см. кладограмму). Пять родов (15 видов): Allothunnus Serventy, 1948, Auxis Cuvier, 1829, Euthynnus Lütken, 1883, Katsuwonus Kishinouye, 1915 и Thunnus South, 1845.
Название семейства Scombridae образовано от имени типового рода Scomber Linnaeus, 1758, название которого происходит от греческого слова σκομβρος (skombros) — «скумбрия», «макрель», древнегреческого названия тунца или скумбрии.
Найденные окаменелости (fossils) ранних скомбрид были датированы нижним эоценом, 55,8 млн лет назад. Время появления некоторых современных родов семейства Скумбриевые [Sepkoski, Jack. A compendium of fossil marine animal genera // Bulletins of American Paleontology — Paleontological Research Institution, 2002. — Vol. 364. — P. 560.] (см. диаграмму):
• † Acanthocybium Gill, 1862 — в начале эоцена, 55,8 млн лет назад (окаменелые останки найдены эоцен США (Вирджиния); миоцен Бельгии, Мексики, США (Вирджиния), Венесуэлы; плиоцен США (Северная Каролина));
• † Euthynnus Lütken, 1883 — в середине миоцена, 15,97 млн лет назад (миоцен Алжира, Мексики);
• † Grammatorcynus Gill, 1862 — в начале олигоцена, 33,9 млн лет назад;
• † Gymnosarda Gill, 1862 — в начале эоцена, 55,8 млн лет назад (эоцен Соединённого Королевства, США (Алабама); олигоцен Венгрии);
• † Sarda Cuvier, 1829 — в начале эоцена, 55,8 млн лет назад (эоцен Казахстана, США (Мэриленд, Вирджиния); олигоцен Грузии, Германии, Российской Федерации; миоцен Алжира, Азербайджана, Италии, США (Калифорния, Северная Каролина); плиоцен Италии, США (Северная Каролина); плейстоцен Италии; четвертичный период Италии);
• † Scomber Linnaeus, 1758 — в начале олигоцена, 33,9 млн лет назад (олигоцен Чешской Республики, Германии, Румынии; миоцен Алжира, Азербайджана, Италии, Японии, Мексики, Российской Федерации, США (Калифорния); плиоцен Греции, Италии; плейстоцен Италии; четвертичный период Италии);
• † Scomberomorus Lacepède, 1801 — в начале эоцена, 55,8 млн лет назад (эоцен Казахстана, Туркменистана, США (Алабама, Вирджиния); олигоцен Грузии, Германии; миоцен Японии, Польши, США (Вирджиния); плиоцен Греции);
• † Thunnus South, 1845 — в середине эоцена, 48,6 млн лет назад (эоцен Грузии, Италии, Украины; олигоцен Грузии, Азербайджана, Чехии, Украины; миоцен Мексики, Чили, США (Калифорния, Северная Каролина, Вирджиния); плиоцен Чили, США (Калифорния, Северная Каролина); четвертичный период Греции, Соединенного Королевства).
Скумбриевые рыбы имеют удлинённое, веретеновидное, у некоторых родов умеренно сжатое, тело, равномерно покрытое мелкой, гладкой циклоидной чешуёй или голое в задней части. В передней части крупные чешуи образуют панцирь (чешуйчатый корсет — область за головой и вокруг грудных плавников покрыта крупной толстой чешуей). Два спинных плавника, обычно разделённых, могут складываться в углубление в виде бороздки. Позади второго спинного и анального плавников расположены 5—12 отчётливых плавничков, помогающих избегать образования водоворотов при быстром движении рыбы. Первый спинной плавник имеет 9—27 лучей, его начало далеко позади головы. Грудные плавники сидят на теле высоко. Брюшные плавники умеренные или маленькие, с 1 колючим и 5 мягкими лучами, расположены на груди ниже грудных плавников. Хвостовой стебель тонкий, с двумя маленькими килями с каждой стороны, между ними у более продвинутых видов имеется более крупный киль. Хвостовой плавник глубоко раздвоенный, с опорными хвостовыми лучами, полностью закрывающими гипуральную пластинку. Рыло заострённое. Рот умеренно большой. Предчелюстная кость (premaxilla — парная кость черепа перед верхнечелюстной костью) клювовидная, без носовых костей, разделённая решетчатой костью (ethmoid — решётчатая непарная кость в черепе, который отделяет носовую полость от головного мозга, расположенная между двумя глазницами). Поэтому верхняя челюсть спереди похожа на клюв. Зубы в челюстях сильные, умеренные или слабые, настоящих клыков нет. На нёбе и на языке могут быть зубы. Жаберные перепонки не приращены к истмусу. Имеется специализированная подкожная система сосудов для теплообмена у видов рода Thunnus и близких родов. У некоторых видов имеется прозрачное жировое веко. Отличительным признаком семейства является костное кольцо вокруг глаз. Позвонков — 31—66. Боковая линия простая, полная, изогнутая или волнообразная, иногда с поперечными ветками. Размер представителей семейства варьирует от 20 см (Rastrelliger faughni) до 4,58 м (Thunnus thynnus) (см. диаграмму).
Распространение (карта): океаны и моря тропического, субтропического и умеренного климата. Некоторые виды обитают или заходят в пресную воду. Скомбриды — пелагические рыбы, жизненный цикл которых не связан с дном. Это быстрые пловцы, хорошо приспособленные к активной жизни в водной толще. Одни виды встречаются в прибрежных водах, другие — далеко от берега (см. схему).
Скумбриевые — быстрые, эпипелагические хищники. На охоте они могут развивать большую скорость благодаря очень обтекаемому телу и убирающимся плавникам. Некоторые виды скумбриевых имеют несколько более высокую температуру тела, чем окружающая вода. В частности тунцы, отличаются частично эндотермичностью (теплокровностью), что также помогает им поддерживать высокую скорость и активность. Другие приспособления включают большое количество красных мышц, что позволяет им сохранять активность в течение длительного времени. Скомбриды, такие как желтопёрый тунец, могут развивать скорость до 75 км/ч. Также у некоторых видов жабры в течение эволюции срослись с фильтровальными приспособлениями и более неподвижны. Скумбрии (Scomber и Rastrelliger) фильтрует планктон своими длинными жаберными тычинками. Испанские скумбрии, бонито и тунец питаются более крупной добычей, включая мелкую рыбу, ракообразных и кальмаров. Основными хищниками более мелких скомбрид являются другие хищные рыбы, особенно крупные тунцы, марлины и парусники (семейство Istiophoridae Rafinesque, 1815).
Макрелевые — раздельнополые рыбы и большинство из видов практически не имеют полового диморфизма в строении или цветовой гамме. Самки многих видов достигают больших размеров, чем самцы. Групповой нерест большинства видов происходит в тропических и субтропических водах, часто вблизи берега. Икра пелагическая, из них вылупляются планктонные личинки. Икра и личинки встречаются только вблизи побережий.
Скумбриевые — одни из важнейших промысловых и спортивных рыб, являются популярными объектами спортивного и промышленного рыболовства. Мясо крупных видов скумбрий накапливает ртуть, содержащуюся в морской воде, в связи с чем, Управление контроля качества продуктов и лекарств США (FDA) порекомендовало воздержаться от употребления скумбрий вида королевская макрель (Scomberomorus cavalla) беременным и кормящим женщинам, а также детям. Макрель — общее название многих разновидностей рыб, в основном семейства скумбриевых (скумбрии, пеламиды и другие). В России под торговым названием «макрель» понимаются все виды рыб из семейства скумбриевых, кроме атлантической скумбрии (Scomber scombrus), которая поступает на рынок как «скумбрия». Также часто «макрель» продается под названием «скумбрия», вводя потребителя в заблуждение. Всемирный фонд дикой природы и Лондонское зоологическое общество совместно 16 сентября 2015 года выпустили свой «Living Blue Planet Report», который гласит, что резкое падение запасов скумбриевых рыб на 74% произошло в мире в периоде между 1970 и 2010 годами, и «число млекопитающих, птиц, рептилий, земноводных и рыб сократились в среднем вдвое всего за 40 лет».
Вымершие представители семейства Скумбриевые:
• † Acanthonotos Sauvage, 1870 — (типовой вид — Acanthonotos armatus Sauvage, 1870) — 7,246–5,332 млн лет назад, миоцен Ликата (Licata), находится в мессинском морском горизонте в Италии. [H. E. Sauvage. 1870. Synopsis des poissons tertiaires de Licata. Anales des Sciences Naturalles, Zoologie et Paleontologie 14:1-26]
• † Ardiodus White, 1931 — (типовой вид — Ardiodus mariotti White, 1931) — палеоцен, слои Олдхавен (Oldhaven Beds), Упнор (Upnor), Кент (Kent), Англия.
• † Caucombrus Bannikov, 2020 — (типовой вид — Caucombrus histiopterygius Bannikov, 2020) — нижний олигоцен, нижний майкоп, пшехский горизонт Северо-Западного Кавказа и Абхазии (реки Белая, Пшеха и Гумиста). Новый таксон отличается от остальных известных Scombridae, как ископаемых, так и современных, высоким парусоподобным первым спинным плавником. Остеологические признаки Caucombrus gen. nov. позволяют сближать его с королевскими макрелями (подсемейство Scomberomorinae). [A. F. Bannikov. 2020. A new genus and species of scombrid fish (Perciformes, Scombroidei, Scombridae) from the Lower Oligocene of the Caucasus. Paleontological Journal 54(1):59-67]
• † Cybium Cuvier, 1829 — 55,8—2,588 млн лет назад:
• † Cybium bartonense Woodward, 1901 — 37,2—33,9 млн лет назад, нижняя челюсть, эоцен Великобритании. [A. S. Woodward. 1901. Catalogue of Fossil Fishes in the British Museum (Natural History), Part IV 1-636]
• † Cybium excelsum Woodward, 1901, череп. [A. S. Woodward. 1901. Catalogue of Fossil Fishes in the British Museum (Natural History), Part IV 1-636]
• † Cybium loczyi Weiler, 1938 — 33,9—28,4 млн лет назад, Будапешт (Budapest), Уйлаки Теглагьяр (Ujlaki Teglagyar), который находится в рупельском морском горизонте в Венгрии. [W. Weiler. 1938. Neue Untersuchungen an Mitteloligozanen Fischen Ungarns. Geologica Hungarica, Series Palaeontologica 15:1-31]
• † Cybium stormsi Leriche, 1905 — 40,4—37,2 млн лет назад, эоцен США (Алабама).
• † Eoscomber Claeson et al., 2015 — (типовой вид — Eoscomber senegalicus Claeson et al., 2015) — 55,8—48,6 млн лет назад, частичный скелет, известняковые карьеры Деспла (Desplats, Нижняя часть), которая находится в прибрежном мергеле / известняке Ипра (Ypresian) в формации Тиес (Thies Formation) в Сенегале. [K. M. Claeson, R. Sarr, R. V. Hill, El Hadji Sow, R. Malou and M. A. O'Leary. 2015. New fossil scombrid (Pelagia: Scombridae) fishes preserved as predator and prey from the Eocene of Senegal. Proceedings of the Academy of Natural Sciences of Philadelphia 164(1):133-147]
• † Eoscombrus David, 1946 — (типовой вид — Eoscombrus chaneyensis David, 1946) — 48,6—33,9 млн лет назад, коха с чешуёй, Jergins Oil Company, ранчо Чейни № 1, Калифорния. [L. R. David. 1946. Some typical Upper Eocene fish scales from California. Contributions to Paleontology, Carnegie Institute of Washington Publication 551:47-112]
• † Gasteronemus Agassiz, 1835 — (типовой вид — Gasteronemus rhombeus Agassiz, 1835 и Gasteronemus oblongus Agassiz 1835) [L. Agassiz. 1835. Recherches Sur Les Poissons Fossiles. Tome IV (livr. 4). Imprimerie de Petitpierre, Neuchatel 33-52]
• † Godsilia Monsch, 2006 — (типовой вид — Godsilia lanceolata Agassiz, 1835) — 55,8—48,6 млн лет назад, встречается только в Пешиаре (Pesciara), зона Alveolina dainelli, эоцен Италии.
• † Landanichthys Dartevelle & Casier, 1949 — 66,0—61,6 млн лет назад, датском период палеоценовой эпохи.
• † Ocystias Jordan & Gilbert, 1920 — (типовой вид — Ocystias sagitta Jordan & Gilbert, 1920) — 11,608—7,246 млн лет назад, скелет, карьер Ломпок ( Lompoc Quarry), который находится в тортонских базальных (силикокластических) диатомитах в формации Монтерей (Monterey Formation) в Калифорнии. [D. S. Jordan and J. Z. Gilbert. 1920. Fossil Fishes of Diatom Beds of Lompoc, California. Leland Stanford Junior University Publications University Series]
• † Ozymandias Jordan & Gilbert, 1919 — (типовой вид — Ozymandias gilberti Jordan & Gilbert, 1919) — 11,608—7,246 млн лет назад, Сан-Педро (San Pedro), который находится в миоценовом морском песчанике в Калифорнии. [D. S. Jordan and J. Z. Gilbert. 1919. Fossil Fishes of the Miocene (Monterey) Formations. Leland Stanford Junior University Publications, University Series 13-60]
• † Palaeothunnus Bannikov, 1978 — (типовой вид — Palaeothunnus parvidentatus Bannikov, 1978) — 55,8—48,6 млн лет назад, обнаружен только у Улья-Кушлюк (Uilya-Kushlyuk), эоцен Туркменистана.
• † Pneumatophorus Jordan & Gilbert, 1883 — (типовой вид — Pneumatophorus euodus Nolf, 1972) — 48,6—40,4 млн лет назад, встречается только на песчаном карьере Остероден (Osteroden), эоцен Германии.
• † Pseudauxides Monsch, 2006 — (типовой вид — Pseudauxides speciosus Agassiz, 1835). Вымерший вид † Pseudauxides speciosus в ископаемом состоянии известен со среднего эоцена (лютет, 45 млн лет назад) из формации Монте-Болька (Monte Bolca) недалеко от Вероны, северная Италия. Сланцы Монте-Болька возникли из отложений древнего океана Тетис и особенно знамениты остатками эоценовых рыб и другой морской фауны. Отпечатки рыб из Монте-Болька великолепно сохранились, и на них зачастую можно различить самые мелкие косточки и чешуйки, а иногда даже внутренние органы и рисунок кожи животных. Подобное наблюдается также и у окаменелостей из Зольнхофена (Бавария). Такая сохранность объясняется погребением останков в илистом дне мелководных лагун и заливов, где образование окаменелостей идёт в условиях с низким содержанием кислорода, и трупы не могут съесть животные-падальщики. [Monsch, K.A. 2006. A revision of scombrid fishes (Scombroidei, Perciformes) from the Middle Eocene of Monte Bolca, Italy. Palaeontology 49: 873-888.]
• † Scombridarum Rifinesque, 1815 — 61,7—28,4 млн лет назад, палеоцен Украины, олигоцен Бельгии, Германии.
• † Scombrosarda Danilt'chenko, 1962 — (типовой вид — Scombrosarda limata Danil'chenko, 1980) — олигоцен, Северный Кавказ. [P. G. Danil'chenko. 1980. Otryad Perciformes. Trudy Paleontologicheskogo Instituta Akademii Nauk SSSR 178:115-169]
• † Thunnoscomberoides Monsch, 2006 — (типовой вид — Thunnoscomberoides bolcensis Agassiz, 1835) — 55,8—48,6 млн лет назад, встречается только в Пешиаре (Pesciara), зона Alveolina dainelli, эоцен Италии. [Monsch, K.A. 2006. A revision of scombrid fishes (Scombroidei, Perciformes) from the Middle Eocene of Monte Bolca, Italy. Palaeontology 49: 873-888.]
• † Tunita Jordan & Gilbert, 1919 — (типовой вид — Tunita octavia Jordan & Gilbert, 1919) — 23,03—5,332 млн лет назад, Эль-Модена (El Modena), которая находится в миоценовых морских сланцах в формации Монтерей (Monterey Formation) в Калифорнии. [D. S. Jordan and J. Z. Gilbert. 1919. Fossil Fishes of the Miocene (Monterey) Formations. Leland Stanford Junior University Publications, University Series 13-60]
• † Turio Jordan & Gilbert, 1920 — (типовой вид — Turio wilburi Jordan & Gilbert, 1920) — 11,608—7,246 млн лет назад, скелет, карьер Ломпок (Lompoc Quarry), который находится в тортонских базальных (силикокластических) диатомитах в формации Монтерей (Monterey Formation) в Калифорнии. [D. S. Jordan and J. Z. Gilbert. 1920. Fossil Fishes of Diatom Beds of Lompoc, California. Leland Stanford Junior University Publications University Series]
• † Wetherellus Casier, 1966 — 55,8—48,6 млн лет назад, встречается только в ипрском морском аргиллите в формации Лондонская глина (London Clay Formation), остров Шеппи (Isle of Sheppey), эоцен Соединенного Королевства. [E. Casier. 1966. Faune ichthyologique du London Clay 1-510]:
• † Wetherellus brevior Casier, 1966 — 55,8—48,6 млн лет назад, эоцен Соединенного Королевства.
• † Wetherellus cristatus Agassiz, 1844 — 55,8—48,6 млн лет назад, частичный скелет, эоцен Соединенного Королевства.
• † Wetherellus longior Casier, 1966 — 55,8—48,6 млн лет назад, эоцен Соединенного Королевства.
• † Xestias Jordan, 1920 — (типовой вид — Xestias iratus Jordan, 1920) — миоцен Южной Калифорнии. [Jordan D. S. 1925. The fossil fishes of the Miocene of Southern California, Stanford University Publications. University Series 4 1, 1-51]
Рыбы открытого океана. Н.В.Парин 1998 г.
Интересную группу скумбриевидных представляют тунцы — рыбы с идеально приспособленным для скоростного плавания веретеновидным телом. Их место в системе рыб вызывало до недавнего времени много разногласий. Дело в том, что у тунцов имеется совершенно уникальная система подкожных кровеносных сосудов (они снабжают кровью боковую мускулатуру), и эта особенность анатомии, впервые обнаруженная японским морфологом К. Кисинуйе, послужила основанием для предложенного им выделения тунцов в особый отряд, принятый вслед за этим автором академиком Л. С. Бергом и некоторыми другими исследователями. Такая точка зрения сейчас полностью отвергается, и близкое родство тунцов с неритическими скумбриями, королевскими макрелями и пеламидами, вместе с которыми они входят в состав семейства Scombridae (в нем, по Б. Коллетту и К. Науэн, 15 родов и 49 видов), никем не оспаривается. Необычность кровообращения представляет собой лишь одно из приспособлений тунцов к столь характерному для них продолжительному быстрому (до 90 км/ч) плаванию, сопряженному с большими энергетичесними затратами. В моменты, требующие максимального расхода энергии, температура тела тунцов значительно повышается — разница между ней и температурой окружающей среды может достигать нескольких градусов. Подкожная система кровеносных сосудов, лучше всего развитая у синего тунца, обеспечивает специальную доставку кислорода к тем мышцам, которые выполняют наибольший объем работы (По мнению А. Н. Заикииа, эта система может обеспечивать также увеличение упругости тела путем эрекции (наполнения кровью) приповерхностных тканей, что позволяет рыбе значительно увеличить частоту колебательных движений хвоста. Аналогичный механизм был недавно обнаружен у китообразных.). Достижению той же цели способствует и высокая кислородная емкость крови этих рыб, у которых содержание гемоглобина в эритроцитах доходит до 21% (у пеламид оно не превышает 8—14%).
Постоянное плавание — наиболее характерная особенность биологии тунцов, и они прекрасно приспособлены к такому образу жизни. Как показал Ю. Г. Алеев, у этих скоростных пловцов (а также у скумбрий, пеламид, меч-рыбы, марлинов) функция движителя целиком перенесена на хвостовой плавник, а короткое обтекаемое тело практически не выполняет локомоторной работы. При остановке у этих рыб даже затрудняется дыхание, так как вода попадает в жаберную полость лишь через постоянно открытый рот при поступательном движении. Таким образом, специализация как неритических, так и океанических тунцов направлена на достижение высоких скоростей плавания и активное перемещение на дальние расстояния.
Я уделил здесь тунцам так много внимания, поскольку именно они могут служить, на мой взгляд, символическим олицетворением некоей «идеальной» эпипелагической рыбы. Любой тунец независимо от видовой принадлежности и размера превосходно приспособлен к жизни в безбрежной и бездонной толще океана - он быстр, резок в движениях, обладает высокой маневренностью, неутомим. Удивительна форма его обтекаемого тела: оно действительно веретеновидное, как справедливо пишут в определителях и описаниях, но, чтобы по-настоящему осознать, что это значит, нужно обязательно увидеть такое «веретено» воочию. Особенно впечатляет почти идеально круглое в любом участке поперечное сечение головы и туловища (ведь у рыб с привычной нам «рыбовидной» формой тела оно всегда сильно уплощено с боков). Поразительно в изящество исключительно тонкого хвостового стебля перед могучим движителем — прочным и жестким хвостовым плавником полулунной формы, который способен вибрировать с невероятной частотой. Первый тунец которого я увидел живым,— это была маленькая ауксида длиной сантиметров в 30, - навсегда остался в моей памяти, а следующие гиганты (мне, правда, не приходилось иметь дело с очень большими экземплярами, но и метр от конца рыла до выемки хвоста — не так уж мало!) немного добавили к исходному образу.
Плодовитость эпипелагических рыб, как правило, очень велика. В качестве примера можно указать, что змеиная макрель и ауксида выметывают до 1 млн икринок, полосатый тунец — до 2 млн, большеглазый тунец и ваху — по 6 млн, синий тунец — 10 млн, меч-рыба — 16 млн, сельдяной король — не менее 20 млн, луварь — 47 млн, большой синий марлин — почти 100 млн, рыба-луна — даже 300 млн икринок. Для преобладающего большинства эпипелагических рыб характерна порционность икрометания, приводящая к большой растянутости сроков нереста. В тропических водах в связи с относительным постоянством условий среды размножение может идти даже круглогодично (например, у летучих рыб и тунцов). Высокая плодовитость и ускоренное прохождение ранних стадий индивидуального развития направлены у костистых рыб на компенсацию огромной смертности икры и личинок при полном отсутствии родительской заботы о потомстве.
Хорошо известно, что границы ареалов эпипелагических рыб, принадлежащих к нектону (т. е. способных к активным горизонтальным перемещениям) , определяются главным образом температурой воды. Связь распространения с температурными условиями показана для многих рыб — акул, лососей, летучих рыб, тунцов, марлинов и др. Минимальные значения температуры, отмеченные для разных видов массовых скумбриевидных рыб открытого океана в период нагула, варьируют от 10—12° у длинноперого тунца и меч-рыбы до 18—20° у желтоперого тунца и синего марлина. Подобные примеры, количество которых может быть многократно увеличено, вполне убедительно демонстрируют значение температуры воды как главного фактора, лимитирующего географическое распространение нектонных рыб.
Активные горизонтальные миграции некоторых эпипелагических рыб, обусловленные пространственным разобщением районов нереста и нагула в соответствии с различиями абиотических (прежде всего температурных) и биотических (в основном кормовых) условий, достигают очень большой протяженности и часто имеют довольно сложный характер. Они еще недостаточно изучены и лишь в немногих случаях подтверждены данными мечения, позволяющими прослеживать их пути. Иногда, впрочем, такие миграции хорошо заметны по перемещениям массовых скоплений и районов промысла. У океанических скумбриевидных рыб они достигают наибольшего размаха: у них отмечены даже сверхдальние трансокеанические миграции.
Способность к дальним активным миграциям представляет собой одно из весьма существенных приспособлений рыб к обитанию в эпипелагиали. Такие миграции делают возможным более полное использование пищевых ресурсов океана, приводя в некоторых случаях даже к полному разобщению репродуктивной и вегетативной частей ареала (особенно характерны в этом отношении миграции, в ходе которых рыбы пересекают границу между тропической и бореальной областями). Именно поиски путей к расширению области нагула — основная причина активных перемещений. Нерестовые и зимовальные миграции служат, как правило, только обратному возвращению мигрантов из районов, в которых происходил их откорм.
Рассматривая миграции эпипелагических рыб, нельзя обойти молчанием вопрос о возможных механизмах их ориентации в открытых водах. Эта проблема активно разрабатывается в последние годы, но все ещё очень далека от разрешения, хотя высокая чувствительность рыб к таким факторам, как температура, соленость, химический состав воды, давно доказана в опытах и не вызывает сомнений. Среди предложенных к настоящему времени гипотез, частично проверенных экспериментально, следует отметить астронавигацию — ориентацию по Солнцу (в том числе по поляризации солнечного света) и другим небесным телам, ориентацию по магнитным полям (у некоторых рыб, в том числе тунцов и марлинов, в черепных костях обнаружен биогенный магнетит), по электрическим полям (восприятие сверхслабых токов может обеспечить ориентацию относительно течений) и даже по гравитационным полям. Все эти гипотезы не дают полного объяснения наблюдаемым в природе процессам, однако твердо установленный факт направленных перемещений некоторых видов рыб требует продолжения научного поиска. Решение вопроса, возможно, будет найдено в результате комплексного подхода.
Современное океаническое рыболовство (т. е. рыболовство за пределами шельфов) имеют недолгую историю. В крупных масштабах оно стало развиваться только в последние годы, но есть все основания считать, что локальный промысел океанических объектов в некоторых районах существовал издавна. Это касается в первую очередь тех участков, где склон начинается в непосредственной близости от берега. В Полинезии, например, существовали (а на некоторых островах существуют и сейчас) традиционные способы лова многих рыб открытого океана — летучек, тунцов, корифен, руветты, акул. Для каждого объекта были разработаны специальные приемы и методы промысла, основанные на прекрасном знании повадок этих рыб. Летучих рыб с незапамятных времен ловили там, используя их положительную реакцию на искусственный свет: в темные ночи их привлекали к лодкам горящими факелами. При ужении мелких тунцов островитяне применяли снасти, изготовленные из перламутровых раковин жемчужниц. Такая «блесна» имеет форму слегка изогнутой стилизованной рыбки длиной 8—10 см, к хвосту которой привязывается под небольшим углом костяное, деревянное или каменное острие, заменяющее крючок, а сзади прикрепляется пучок перьев или свиной щетины. Обнаружив стаю мелких тунцов или пеламид (рыб ищут, наблюдая за их пищевыми конкурентами — морскими птицами) и приблизившись к ней, рыбак тянет блесну за лодкой, а при поклевке тунца резким рывком, продолжающим его движение в воде, выбрасывает добычу в каноэ. Нужно добавить, что полинезийцам прекрасно известна привычка мелких рыбешек собираться у плавучих предметов, где их и находят тунцы. Поэтому, увидев плавник — бревно, ветки или что-нибудь подобное, рыбаки забирают его в свое каноэ, под которое переходят укрывавшиеся там рыбки, после чего спокойно ожидают появления тунцов или корифен. Крупных тунцов и марлинов ловят на наживленные крючки, иногда с использованием живой приманки, сохраняющейся в плавучих корзинах. Особенно специфичен промысел руветты (местное название «уравена», или «куравена»), при котором попадаются и другие хищные рыбы, обитающие в верхней части склона. Для этого рыбаки выходят в океан ночью и опускают на дно на глубину около 200 м наживку, насаженную на своеобразный деревянный крючок довольно сложной конструкции (эти крючки, как и тунцовые «блесны», имеют свою специфику на разных островах Океании).
Для промысла крупных, постоянно находящихся в движении хищных рыб находят широкое применение плавные яруса — гигантские крючковые снасти, устроенные по типу переметов. Ярус состоит из отдельных секций (их называют «корзинами»), которые, соединяясь между собой, образуют порядок любой желаемой длины. Обычная длина рабочего яруса составляет 50—75 км, но иногда достигает 100 км и более. Основной несущей частью снасти служит «хребтина» из жестко закрученной высокопрочной веревки, к которой подвязываются поводки с крючками, опускаемыми на глубину 100—200 м (наживкой служит мороженая рыба или кальмары), и буйрепы с прикрепленными к ним стеклянными или пластиковыми поплавками (кухтылями), поддерживающими ярус на нужном горизонте. Постановку производят раз в сутки с таким расчетом, чтобы он находился в работе во время утренней зари, когда рыба клюет лучше всего. Этот тип промысла использует главным образом рассредоточенные концентрации крупных тунцов, живущих в подповерхностных слоях воды — желтоперого, большеглазого, альбакора, а также марлинов, меч-рыбы и эпипела-гических акул. В лучших промысловых районах улов может достигать 25 рыб общей массой более 1 т на 100 крючков, однако даже поимка 3—6 тунцов (150—250 кг) на сотню крючков считается более чем достаточной для ведения рыболовства. Поэтому ярусный промысел производится практически на всей акватории тропической зоны, хотя в основном тяготеет, естественно, к ее продуктивным районам.
Очень своеобразен и удебный лов тунцов, применяемый для эксплуатации поверхностных скоплений полосатого тунца (скипджека), неполовозрелого желтоперого тунца и других некрупных скумбриевидных рыб. Когда тунцеловный клиппер (так называется специализированное для такого промысла судно) приблизится к замеченному косяку, включают пульверизационную систему, разбрызгивающую воду (имитация всплесков мелкой рыбешки), и начинают выбрасывать в море живую приманку — анчоусов или сардин, которых содержат в проточных живорыбных цистернах. Тунцы набрасываются на приманку, жадно поедая ее, и в это время хватают и подкинутые им наживленные крючки. Этот промысел требует большой сноровки, так как рывки тунцов во время клева очень сильны. Бывают случаи, когда неопытные ловцы вместе с удочкой оказываются за бортом. Хороший рыбак, почувствовав натяжение лески, должен резко откинуться назад и, используя инерцию броска тунца, одним движением выбросить его на палубу.
Для лова тунцов у поверхности используют также тролловую снасть (дорожку), которая обычно примениется с клипперов и ярусных судов во время переходов. С этой целью по обеим сторонам судна устанавливают перпендикулярно его продольной оси длинные шесты (выстрелы), с каждого из которых буксируют по несколько лесок, несущих крючки, замаскированные султанчиками из конского волоса или капрона.
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
Семейство Скумбриевые (Scombridae)
Тело торпедовидное. Хвостовой плавник гомоцеркальный, его стебель тонкий с мелкими мягкими плавничками сверху и снизу. Рот конечный, большой. Хищные океанические рыбы толщи воды. В наших водах обычны два вида.
Иллюстрированная энциклопедия рыб. Ст.Франк 1983 г.
Семейство Скумбриевые (Scombridae) известно со времен эоцена. Ныне живет около 50 видов. У этих рыб отсутствует плавательный пузырь.
Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Часть 4. (Perciformes). Г.У.Линдберг, З.В.Красюкова 1975 г.
CLXIII. Сем. SCOMBRIDAE — СКУМБРИЕВЫЕ
Тело удлиненное, веретеновидное, более или менее умеренно сжатое с боков. Хвостовой стебель с каждой стороны с двумя небольшими килями между лопастями хвостового плавника и обычно также с большим килем впереди них. Тело, покрыто мелкой чешуей или голое сзади; в передней части своеобразный, более или менее выраженный панцирь из увеличенных чешуй. Боковая линия слегка изогнута или с волнообразными изгибами, часто с поперечными ветками. Жировое веко есть или отсутствует. Два более или менее отделенных друг от друга или соприкасающихся спинных плавника. Анальный с 1—3 колючими лучами. Последние лучи второго спинного и анального плавников обособлены в виде отдельных небольших плавничков. Грудные плавники расположены высоко, брюшные на груди, с колючкой и 5 ветвистыми лучами. Зубы мелкие или крупные, конические или более или менее сжатые с боков, иногда ножевидные, на челюстях, иногда зубы имеются на нёбных костях и на сошнике. Жаберные тычинки есть или отсутствуют, немногочисленные. Позвонков 31—66 (Световидов, 1964 : 384).
Около 13 родов. Родовой состав этого семейства анализирует Коллетт (Collette, 1962, 1966). Тропические, субтропические и отчасти умеренные моря. Большинство представителей имеют очень большое значение в мировом промысле (Линдберг, 1927; Расс, 1948, 1960, 1965; Линдберг, 1971). Биологическая и технологическая характеристика многих скомброидных рыб приведена В. Г. Осиновым и А. Ф. Мякшой (1961). В Японском море известно 8 родов.
Следует отметить, что все упоминаемые нами в тексте промеры представителей подотряда Scombroidei соответствуют схеме, предложенной и широко используемой ихтиологами (рис. 211) (Jones a. Silas, 1961 : 372).
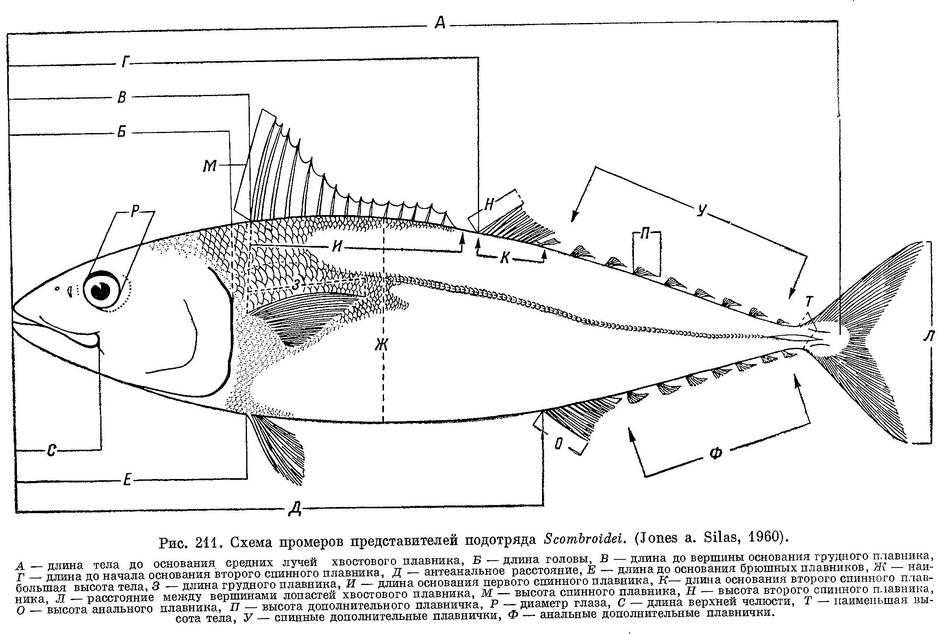
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ СЕМ. SCOMBRIDAE
(По Мацубаре (Matsubara, 1955 : 514), с дополнениями.)
1 (16). Зубы на челюстях слабые, конусообразные.
2 (11). Брюшные межплавниковые лопасти отделены друг от друга и от плавников (рис. 212, А, Б). Оба спинных плавника соприкасаются или если только сближены, то расстояние между ними не более диаметра глаза.
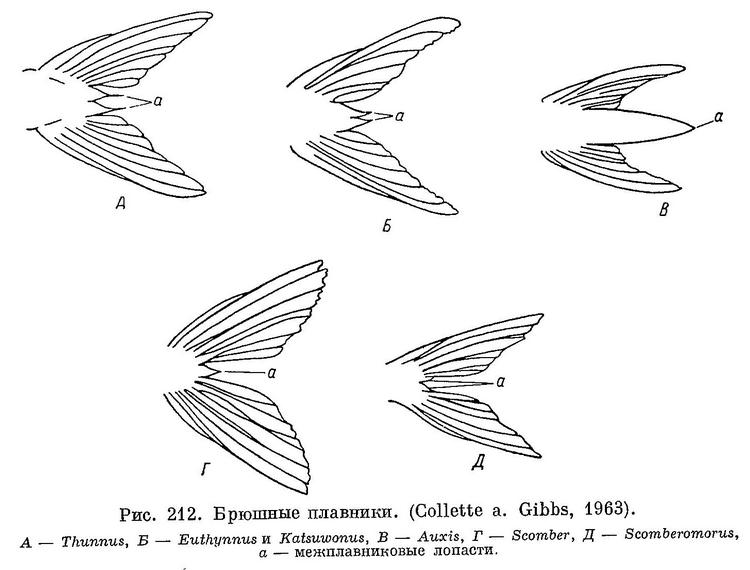
3 (6). Тело полностью покрыто чешуей. Чешуя корсета и боковой линии обычно крупнее, чем чешуя других частей тела.
4 (5). Тело округлое в поперечном сечении, не сжатое с боков. Нет темных
продольных полос вдоль верхней половины тела. На сошнике и нёбных костях имеются мелкие бархатистые зубы. Позвонков 39—41. 1. Thunnus South.
5 (4). Тело немного сжато с боков. От 5 до 10 узких темных продольных полос на верхней половине. На сошнике нет зубов; на нёбных костях один ряд сильных конических зубов. Позвонков 44. 2. Sarda Cuvier.
6 (3). Тело голое без чешуи, за исключением корсета и боковой линии.
7 (8). Спинные плавники соприкасаются. Верхний край первого спинного плавника прямой, без выемки. Распространен от Красного моря до Австралии, Индонезии и Японии. [Gymnosarda Gill, 1862].
8 (7). Спинные плавники не соприкасаются. Верхний край спинного плавника с выемкой.
9 (10). Зубы имеются не только на обеих челюстях, но и на нёбных костях, иногда и на сошнике. Позвонков 39. Продольные черные полосы на нижней части тела отсутствуют. 3. Euthynnus Jordan et Gilbert.
10 (9). Зубы только на челюстях. Позвонков 41. 3—5 заметных продольных черных полос на нижней части тела. 4. Katsuwonus Linne.
11 (2). Брюшные межплавниковые лопасти сливаются друг с другом (рис. 212, В, Г) или каждая из них прирастает к прилежащему плавнику (рис. 212, Д). Спинные плавники широко расставлены, расстояние между ними более диаметра глаза.
12 (13). Тело не полностью покрыто чешуей, хорошо развит корсет. В задней части хвостового стебля хорошо выраженный срединный киль и два маленьких киля в основании хвостового плавника. На сошнике имеются зубы, нет зубов на нёбных. Позвонков 39. 5. Auxis Cuvier.
13 (12). Тело полностью покрыто чешуей, корсет либо плохо развит, либо отсутствует. В задней части хвостового стебля нет срединного киля, а только 2 маленьких киля. Позвонков 31.
14 (15). На сошнике и нёбных имеются зубы. Жаберные тычинки умеренно длинные и тонкие; их число на нижней ветви первой жаберной дуги меньше 35. Тело имеет веретенообразную форму. Высота тела меньше длины головы. 6. Scomber Linne.
15 (14). На сошнике и нёбных нет зубов. Жаберные тычинки очень длинные, расширенные, их число — больше 35. Тело сжато с боков. Высота его почти равна длине головы. Распространен в Индийском и в западной части Тихого океана (Жаров и др., 1961 : 19). [Rastrelliger Jordan et Dickerson, 1908].
16 (1). Зубы на челюстях сильные, заметно сжатые с боков, имеют почти треугольную форму или ножеобразны.
17 (18). Боковая линия разделяется на 2 ветви позади головы (под первым спинным плавником). Одна из них проходит по верху спины, другая — по низу брюха. Задний край верхнечелюстной кости простирается не далее вертикали переднего края глаза. Позвонков 31. Известен в Красном море, у берегов о-вов Ява, Целебес, Рюкю, Филиппинских, Маршалловых, Гавайских и у побережья Австралии (Beaufort a. Chapman, 1951 : 216). [Grammatorcynus Gill, 1862].
18 (17). Боковая линия одна, не разделена на ветви.
19 (20). Жаберные лепестки образуют сеть. Жаберных тычинок мало. Колючих лучей в спинном плавнике 14—20. Позвонков 40—51. Рыло значительно меньше остальной части головы. 7. Scomberomorus Lacepède.
20 (19). Жаберные лепестки не образуют сеть. Жаберных тычинок нет. Колючих лучей в спинном плавнике 25—26. Позвонков 64. Рыло такой же длины, как остальная часть головы. 8. Acanthocybium (Cuvier et Valenciennes).
Ихтиопланктон Черного моря. Т.В.Дехник 1973 г.
СЕМ. SCOMBRIDAE — СКУМБРИЕВЫЕ
Представлено в Черном море 5 видами, относящимися к 4 родам: Scomber, Sarda, Thunnus, Euthynnus (Световидов, 1964).
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Семейство Скумбриевые (Scombridae)
Скумбриевидные — хорошо
обособленный подотряд, все
представители которого обитают в море и ведут
пелагический образ жизни, не будучи
связанными с дном ни в каком периоде
жизненного цикла. Они характеризуются
удлиненным веретеновидным телом, тонким
и более или менее сжатым с боков
хвостовым стеблем, снабженным двумя
небольшими килями между лопастями
хвостового плавника и у большинства родов
большим средним килем впереди них,
наличием дополнительных плавничков позади
мягкого спинного и анального плавников,
а также рядом других особенностей.
Внешний вид и многие анатомические признаки
скумбриевидных рыб свидетельствуют об
их принадлежности к числу быстрых
пловцов, хорошо приспособленных к
активной жизни в толще воды.
Этот подотряд, заключающий 15 родов
и около 40 видов, распадается на
несколько естественных групп, которые
нередко выделяются в ранге самостоятельных
семейств. Многие ученые считают,
например, что скумбрии, пеламиды,
королевские макрели и тунцы должны
рассматриваться обособленно. Действительно,
наиболее типичные представители этих
групп достаточно резко различаются
между собой и имеют, казалось бы, мало
общего. Между ними располагаются,
однако, роды, занимающие промежуточное
положение, в результате чего границы
между такими группами стираются и они
незаметно переходят одна в другую.
Поэтому всех скумбриевидных обычно
объединяют в одно семейство.
Наиболее примитивным видом
подотряда считают обычно гастрохизму (Gasterochisma melampus), которая в отличие от
других скумбриевидных имеет очень
крупную чешую и лишена килей на хвостовом
стебле.
К группе скумбрий относят два
близких рода — настоящих скумбрий и
тропических скумбрий, каждый из которых
содержит по три вида. Спинные плавники
этих рыб разделены широким
промежутком, а срединный киль на хвостовом
стебле у них отсутствует. Размеры скумбрий
относительно невелики (не более 60 см),
и все они относятся к числу планктоноядных рыб, обитающих в прибрежной
пелагиали и редко встречающихся за
пределами материкового шельфа и прилегающих
к нему вод. Икра у них пелагическая.
К группе пеламид относятся четыре
рода, характеризующиеся коротким веретеновидным телом, неудлиненным рылом,
сближенными спинными плавниками и
хорошо развитыми хвостовыми килями.
Королевские макрели, или Сьерры-макрели (род Scomberomorus,
заключающий 9 видов), имеют сильно вытянутое
тело и удлиненное рыло с большим ртом,
вооруженным крупными треугольными
зубами.
По внешнему облику королевских
макрелей несколько напоминает ваху (Асапthocybium solanderi) — прогонистая рыба
с длинным рылом, имеющая на боках
около 30 темных поперечных полос.
Последнюю, но, пожалуй, наиболее
интересную группу в составе семейства
скумбриевых образуют тунцы, место
которых в системе рыб вызывало до самого
последнего времени много разногласий.
Дело в том, что у тунцов имеется
совершенно уникальная система подкожных
кровеносных сосудов, снабжающих
кровью боковую мускулатуру, среди которой
обособляются прилегающие к
позвоночному столбу участки со своеобразным
строением — красные мышцы. Эта
особенность анатомии, впервые
обнаруженная японским морфологом К.Кисинуйе, послужила основанием для
предложенного им выделения тунцов в
особый отряд, принятый вслед за этим
автором и некоторыми другими
исследователями.
Такая точка зрения сейчас полностью
отвергается, и близкое родство тунцов
и остальных скумбриевых не подвергается
больше сомнению. Необычность
кровообращения представляет собой лишь одно
из приспособлений тунцов к
продолжительному быстрому плаванию,
сопряженному с большими энергетическими
затратами и столь характерному для этих рыб,
которые способны плыть со скоростью
90 км/час. В моменты, требующие
максимального расхода энергии, температура
тела тунцов значительно повышается —
разница между ней и температурой
окружающей воды может достигать
нескольких градусов. Подкожная система
кровеносных сосудов обеспечивает специальную
доставку кислорода к тем мышцам, которые
выполняют наибольший объем работы.
Достижению той же цели способствует и
высокая кислородная емкость крови этих
рыб, у которых содержание гемоглобина
в эритроцитах доходит до 21 г % (у
пеламид оно не превышает 8—14 г %).
Постоянное плавание — наиболее
характерная особенность биологии тунцов.
При остановке у них даже затрудняется
дыхание, так как механизм,
открывающий жаберные крышки связан у этих
рыб с поперечным изгибанием тела влево
и вправо при движении хвостового
плавника. Вода в жаберную полость поступает
у них лишь через постоянно открытый рот
при поступательном передвижении.
Таким образом, специализация тунцов
направлена на достижение высоких
скоростей плавания и активное перемещение
на дальние расстояния.
К группе тунцов относится пять родов.
Четыре из них объединяют сравнительно
некрупных рыб, из которых немногие
превышают 1 м в длину. Все они
характеризуются сильной редукцией чешуйного покрова, развитого только в области
грудного «корсета» и вдоль боковой
линии.
Южный тунец (Allothunnus fallai),
лишенный подкожной кровеносной системы,
очень близок к пеламидам. Эта планктоноядная рыба обитает в субтропических
водах южного полушария. Остальные виды мелких тунцов
приурочены в своем распространении к
тропической зоне. Все они ведут стайный образ
жизни в поверхностных слоях и часто
выпрыгивают из воды.
Промысловое значение тунцов очень
велико. Эти рыбы относятся к числу
наиболее ценных в пищевом отношении и
используются главным образом на
приготовление консервов и других
деликатесных рыбных продуктов. Особенно высоко
ценится на мировом рынке мясо
альбакора, а также полосатого, синего и
желтоперого тунцов. Гастрономические
качества малых тунцов, и особенно макрелевого,
значительно хуже.
При промысле этих рыб используются
различные методы лова, но наибольшее
значение в добыче тунцов принадлежит
плавным ярусам, кошельковым неводам,
дорожкам-троллам и удебному лову.
Плавные тунцеловные яруса, которые
особенно широко используются японскими
рыбаками, представляют собой гигантские
крючковые снасти, устроенные по типу
переметов. Ярус состоит из отдельных
секций, которые, соединяясь между собой,
образуют порядок любой желаемой
длины. Обычная длина рабочего яруса
составляет 40—75 км, но иногда достигает 100 км
и более. Основной несущей частью снасти
служит «хребтина», к которой
подвязываются поводки с крючками (наживкой
служит мороженая рыба или кальмары)
и буйрепы с прикрепленными к ним
стеклянными поплавками,
поддерживающими ярус на нужной глубине. Ярус
выметывают в предрассветные часы и после
недолгого дрейфа приступают к выборке.
Этот тип промысла использует главным
образом рассредоточенные концентрации
крупных тунцов, живущих в
подповерхностных слоях воды — желтоперого,
большеглазого, альбакора, а также марлинов
и меч-рыб. В лучших промысловых
районах улов может достигать 25 рыб (общим
весом более 1 т), однако даже поимка
3—6 тунцов (150—350 кг) на 100 крючков
считается вполне достаточной для
ведения рыболовства. Поэтому ярусный
промысел проводится практически на всей
акватории тропической зоны, хотя в
основном тяготеет, естественно, к ее
продуктивным районам.
Очень своеобразен и удебный лов
тунцов, применяемый для эксплуатации
поверхностных скоплений мелких тунцов,
в частности скипджека. Когда
промысловое судно приблизится к замеченному
косяку, рыбаки начинают выбрасывать в
море живую приманку — анчоусов, которых
содержат в проточных живорыбных
цистернах. Тунцы набрасываются на
приманку, жадно поедая ее, и в это время
хватают и подкинутые им крючки. Этот
промысел требует большой сноровки, так как
рывки тунцов во время клева очень
сильны. Бывают случаи, когда неопытные
ловцы вместе с удочкой оказываются за
бортом. Хороший рыбак, почувствовав
натяжение лески, должен резко откинуться
назад и, используя инерцию рывка тунца,
одним взмахом выбросить его на палубу
Обитатели бездны. Уильям Кроми 1971 г.
К скумбриевидным принадлежат наиболее быстроходные обитатели Мирового океана. Это прожорливое, чрезвычайно хищное племя состоит из 60 различных видов макрелей и тунцов; сюда входят также марлины и меч-рыба. Многие проходят огромные расстояния от мест кормежки до нерестилищ. Большинство из них обладают обтекаемой, стремительной формой, как бы воплощающей их мощь. Зачастую бока этих рыб и брюхо серебристые или радужные, а спина роскошного сине-зеленого цвета. Умеющие постоять за себя, рыбы этого рода доставят вам немало волнующих минут, если вы решите половить их или понаблюдать за ними.
Прекрасно приспособленные к быстрому и постоянному движению, макрель и тунец тонут или задыхаются, если перестанут плыть. Их небольшой плавательный пузырь не может поддержать вес их тела, если оно неподвижно; ко всему, жабры должна все время омывать вода, снабжая их достаточным количеством кислорода. Благодаря постоянному сокращению мышц температура тела тунцов на 3—4° выше температуры водной среды.
Не занятые охотой, тунцы движутся достаточно быстро, чтобы дышать и создавать соответствующую подъемную силу, сохраняя при этом нужную глубину. При запахе, виде или звуке пищи они тотчас увеличивают скорость. Желтоперый тунец развивает скорость до 80 километров в час. При такой скорости саблевидные грудные плавники для лучшей обтекаемости убираются в мелкие канавки. Глубоко вырезанный, твердый, как кость, похожий на молодой месяц хвост быстрыми ударами из стороны в сторону подталкивает тунца, в то время как остальная часть его тела остается почти неподвижной.
Эти «живые метеоры» — не только самые быстрые обитатели моря (Крупная меч-рыба, а также, по-видимому, марлины и парусники на рывках дают большую скорость, чем тунцы.), но и наиболее крупные из костистых рыб. Тунец обыкновенный (Thunnus thynnus) достигает в длину более 4 метров и веса 720 килограммов. Тунец — предмет вожделений всех рыбаков мира. Нередки случаи, когда спортсмены целых 12 часов вываживали тунца, прежде чем им удавалось его забагрить.
В течение многих лет американские тунцеловные клиперы преследовали этих тигров моря от южной части Калифорнии до районов, находящихся далеко к югу от экватора. Теперь современные сейнеры, оснащенные огромными кошельковыми тралами из нейлона, ведут промысел в основном у берегов центральной и северной части Южной Америки, набивая «кошелку» циносционами, скипджеками, длинноперыми и голубыми тунцами и похожими на тунца акулами-бонито. Американские рыбаки добывают ежегодно около 1350 тонн тунца. Это составляет 17 миллионов ящиков рыбных консервов в год. Стоят они 200 миллионов долларов — доход, уступающий лишь доходу от промысла креветок и лосося.
Рыбы северных морей СССР. А.П.Андрияшев 1954 г.
Подотряд Scombroidei
XXXIX. Сем. SCOMBRIDAE — СКУМБРИЕВЫЕ
Scombridae (частью) Regan, Ann. Mag. Nat. Hist., (8), III, 1909 : 72. — Starks, Journ. Morphol., XXI, 1910 : 79. — Солдатов и Линдберг, Обзор рыб дальневост. морей, 1930 : 101.
Scombridae Kishinouye, Journ. Coll. Agric. Univ. Tokyo, VII, 3, 1923 : 400 (моногр.). — Берг, Система рыб, 1940 : 321.
Тело умеренно удлиненное, более или менее сжатое с боков, равномерно утончается к тонкому хвостовому стеблю, который не имеет продольного бокового киля. Чешуя мелкая, в передней части тела нет своеобразного панцыря из увеличенных чешуй. Первый спинной плавник короткий, из тонких колючих лучей, отделен от II D широким промежутком. Передние лучи II D и А не увеличены. Сверху и снизу на хвостовом стебле по 4—6 дополнительных плавничков. Грудные плавники посажены высоко (на уровне глаза); брюшные I 5, под грудными. Жировое веко хорошо развито у всех представителей семейства. Верхняя челюсть не выдвижная, клювовидная; praemaxillaria короткие, плотно соединены с maxillaria. Кольцо suborbitalia полное, пронизано каналами системы боковой линии. Opisthoticum расположено на нижней поверхности черепа, не вдается между occipitale laterale и pteroticum; последнее не отделено от полости мозга. Posttemporale прикреплено к заднему концу pteroticum. Миодом открывается сзади маленькой поперечной щелью. Плавательный пузырь имеется или отсутствует. Позвонков 30—31. Известны с эоцена.
3 рода в северных частях Тихого и Атлантического океанов и в Индовестпацифической области. На севере один род.
Подотряд Thunnoidei (Plecostei)
(Эта группа рыб выделяется многими современными исследователями в самостоятельный отряд (Кишиное, Берг).)
XLI. Сем. THUNNIDAE — ТУНЦОВЫЕ
Thunninae Stаrks, Journ. Morphol., XXI, 1910 : 80 (остеология).
Thunnidae + Katsuwonidae Kishinouye, Journ. Coll. Agricult. Univ. Tokyo, VIII, 2, 1923 : 428 (моногр.).
Thunnidae Берг, Система рыб, 1940 : 333.
Близки к группе Scombroidae (именно к Cybiidae), но, как показали исследования Кишиное, отличаются мощным развитием кожной сосудистой системы, связанной с сосудистым сплетением в боковых мышцах тела; эти части бокового мускула расположены по обеим сторонам позвоночника и выделяются своим тёмнокрасным, почти черным цветом. Другое своеобразное сосудистое сплетение развито на внутренней стороне печени или в гемальном канале. В связи с описанными выше особенностями строения кровеносной системы стоит, повидимому, замечательная особенность этих рыб: температура крови тунцов выше температуры окружающей воды (разница до 9—10°).
Тело веретенообразное, не сжатое с боков. Хвостовой стебель тонкий, с костным боковым килем с каждой стороны.
Спинные плавники обычно сближены (исключая Auxius). Дополнительных плавничков 7—10. Все тело покрыто чешуей или голое (Auxidinae по Бергу = Katsuwonidae Kishin.), но в передней части его всегда имеется своеобразный панцирь из увеличенных чешуй. Жировое веко не развито. Позади prooticum глубокая ямка или костная складка, отделяющая pteroticum от полости мозга. У всех родов (кроме Auxius) на верхней стороне черепа имеется пара крупных отверстий. Миодом открывается в специальную полость парасфеноида. Плавательный пузырь имеется или отсутствует. Позвонков около 39. Известны от нижнего олигоцена.
6 родов, которые Берг относит к 2 подсемействам — Thunninae и Auxidinae. На севере как большая редкость может встретиться лишь один представитель этой группы крупных пелагических рыб.