
| П |
2005000000-076 |
79-87 НП |
|
| 054(02)-88 |
ББК 28.693.32
П 18 УДК 597
Автор предисловия член-корреспондент АН СССР А. П. Андрияшев
Рецензенты: член-корреспондент АН СССР М. Е. ВИНОГРАДОВ, доктор биологических наук П. А. МОИСЕЕВ Парин Н. В.
П18 Рыбы открытого океана/Отв. ред. и авт. предисл. А. П. Андрияшев.— М.: Наука, 1988.—272 с.: ил. — (Серия «Человек и окружающая среда»), ISBN 5-02-005246-9
Утверждено и печати
Редколлегией серии «Научно-популярная литература»
Академии наук СССР
Редактор издательства А. М. Гидалевич. Художник А. В. Шершуков. Художественный редактор В. с. Филатович. Технический редактор Е. Ф. Альберт. Корректоры К. П. Лосева, Р. В. Молоканова
ISBN 5-02-005246-9 © Издательство «Наука», 1988
Несколько слов следует сказать о ее авторе. Н. В. Парин начал свою работу по изучению океанских рыб более 30 лет назад скромным лаборантом на «Витязе». Затем последовала напряженная работа почти в двух десятках больших океанографических экспедициях в Тихом, Индийском и Атлантическом океанах, в которых он всегда работал как опорный ихтиолог, хотя и занимал разные должности вплоть до заведующего лабораторией и начальника рейса. Обладая незаурядным талантом исследователя-натуралиста и огромной работоспособностью, Н. В. Парин, быстро накапливая знания и опыт, приобрел широкую известность как специалист по ряду групп рыб пелагиали Мирового океана.
Обе его диссертации не были лишь необходимыми мероприятиями для приобретения очередных ученых степеней и званий, как это нередко бывает. Это были нужные исследования высокого научного уровня. Кандидатская работа «Основы системы летучих рыб» (1962) явилась первым в нашей стране серьезным исследованием обширной и малоизученной группы рыб, населяющей эпипелагиаль теплых вод всех океанов. Опубликование ее сразу принесло автору широкую известность как наиболее авторитетному знатоку семейства. Докторская диссертация «Рыбы океанской эпипелагиали», опубликованная отдельной книгой (1968), явилась значительным событием в отечественной и мировой ихтиологии благодаря полноте характеристики ихтиофауны пелагиали и широте и новизне эколого-географических обобщений. Затем последовала серия исследований по систематике и распространению многих семейств глубоководных рыб пелагиали и плодотворное участие как докладчика на международных конгрессах и как автора в ряде капитальных советских и зарубежных руководств. В результате к настоящему времени Н. В. Парин естественно выдвинулся в число выдающихся ихтиологов широкого профиля, будучи к тому же весьма активно работающим и, что очень важно, всегда доводящим до опубликования результаты своих многочисленных экспедиционных исследований.
Объективным признанием научных заслуг Н. В. Парина явилось избрание его почетным иностранным членом Американского общества ихтиологов и герпетологов (это звание одновременно могут иметь не более 10—15 наиболее авторитетных ихтиологов из всех внеамериканских стран мира) и вице-президентом Европейского ихтиологического союза.
В 1982 г. Н. В. Парин был приглашен пленарным докладчиком на IV Конгресс европейских ихтиологов в Гамбурге. Большой доклад, с которым он успешно выступил, способствовал в значительной мере обобщению всех данных по ихтиофауне открытого океана, что и составляет предмет настоящей книги. Она продолжает все расширяющуюся тематику исследований автора, включая возможно точную характеристику и ряд обобщающих классификаций не только по ихтиофауне верхней пелагиали океана, но по всей его многокилометровой толще, включая и фауну глубоководных донных рыб.
Член-корреспондент АН СССР А. П. Андрияшев
От автора
В этой книге рассказывается о рыбах, населяющих безбрежные просторы открытого океана. Некоторые из них — сайра, тунцы, палтусы, макрурусы — уже используются рыболовством, в том числе отечественным, и хорошо знакомы большинству потенциальных читателей по крайней мере в виде кулинарных полуфабрикатов или консервных изделий. Другие — летучие рыбы, меч-рыба, марлины, акулы — довольно часто упоминаются в научно-популярной литературе, описаниях морских путешествий и даже в художественной прозе (достаточно вспомнить хотя бы такие произведения, как «Моби Дик» Г. Мелвилла или «Старик и море» Э. Хемингуэя). Впрочем, доля этих общеизвестных видов невелика, и подавляющее большинство океанических рыб пока остается в сфере интересов одних только специалистов-ихтиологов, профессионально занимающихся их исследованием. К этому нужно добавить, что рыбы открытых вод Мирового океана в целом изучены несравнимо хуже, чем те, которые обитают в прибрежной зоне, а обобщающих сводок по ним до сих пор вообще не существует. Таким образом, в отличие от «нормальной» последовательности событий (сначала научное обобщение, а уж затем его популяризация) эта научно-популярная книга предшествует появлению специальной сводки, что, естественно, налагает на автора особую ответственность. Ведь совсем не просто одновременно удовлетворить запросы как широкого круга любителей природы, так и лиц, специальные интересы которых в той или иной мере соприкасаются с океанической ихтиологией, — биологов, океанологов, моряков, работников рыбной промышленности.
Наука, которой я занимаюсь, называется океанической ихтиологией. Это значит, что предметом моего исследования служат рыбы, живущие там, где берегами, как говорится, даже не пахнет. Для сбора любой информации по этим рыбам необходимо идти в открытое море, и поэтому довольно значительная часть моей жизни - в общей сложности около шести лет — прошла в дальних плаваниях. Позади остались 18 экспедиций на судах разного класса и назначения — еще в студенческие годы я первый раз вышел в океан на борту маленького логгера «Изумруд» (это было в мае 1953 г.). Потом, уже будучи сотрудником Академии наук СССР, сделал 10 рейсов на легендарном «старом» «Витязе» (в начале своей карьеры я занимал на нем должность лаборанта в постоянном плавсоставе), работал на других научно-исследовательских кораблях — «Академик Курчатов», «Дмитрий Менделеев», «Рифт», «Профессор Месяцев», «Профессор Штокман», а иногда и на промысловых судах. Мне довелось побывать в Тихом, Атлантическом и Индийском океанах, пересечь все меридианы и почти все, кроме расположенных за Полярными кругами, параллели (с или быть совсем точным, все параллели между 64° с. ш. и 62° ю. ш.). С большим трудом преодолеваю искушение перечислить здесь те далекие порты и экзотические острова, куда заносила меня благосклонная фортуна...
В этой книге речь пойдет о рыбах, населяющих только воды открытого океана, той части водной оболочки Земли, которая находится вне границ прибрежной зоны. Естественно, сразу возникает вопрос, где именно начинается этот самый «открытый океан». В современной биологической океанологии и рыбохозяйственной науке это словосочетание употребляется сейчас в двух значениях. Во-первых, так обозначается часть Мирового океана, находящаяся за пределами 200-мильных экономических зон, повсеместно введенных прибрежными государствами в 1976—1978 гг. и находящихся под их юрисдикцией, во-вторых, воды, лежащие мористее внешнего края материковой отмели (шельфа). Первое понятие восходит, таким образом, к юриспруденции, второе — к ландшафтоведению и имеет вполне определенный биотопический смысл.( Биотоп - участок среды обитания, занятый биоценозом (сообществом), т. е. совокупностью совместно встречающихся видов.) Тем не менее в приконтинентальных областях они, по существу, означают почти одно и то же, поскольку «экономические зоны» практически совпадают с шельфовыми участками. Такого соответствия нет и не может быть, однако, в отношении 200-мильных зон, окружающих венчающие подводные поднятия острова, которые практически лишены шельфа, так что по своим биологическим особенностям эти зоны бесспорно являются частью открытого океана. В то же время к открытому океану не относятся безостровные участки шельфа, даже не находящиеся под юрисдикцией какой-либо страны (уникальный пример такого рода представляет «осколок Гондваны» — огромная банка Сая-де-Малья в западной части Индийского океана).
В бентали и бентопелагиали Мирового океана три зоны — верхняя, средняя и нижняя — существуют в двух модификациях субконтинентальной и талассной, а зона верхней пелагиали даже в трех — субконтинентальной (неритической) собственно океанической и талассной (псевдонеритической). Из семи субконтинентальных биотопов к океаническим относятся, однако, только донные и придонный биотопы материкового склона (соответствующие модифинации трех биотопов верхней зоны являются шельфо-нeритическими) (табл.1).
Таблица 1
| вертикальные зоны и их примерные границы по глубине |
модификации биотопов |
| океаническая |
талласная |
субконтинентальная |
| верхняя (от поверхности до 100-200м) |
эпипелагиаль |
эпипелагиаль, эпибенталь, эпибентопелагиаль |
все биотопы заняты шельфо-неритическими сообществами |
| средняя (от 100-200 до 1000-2000 м) |
мезопелагиаль |
мезопелагиаль, мезобентопелагиаль |
мезопелагиаль, мезобентопелагиаль |
| нижняя от 1000-1500 до 3000 м |
батипелагиаль |
батибенталь, батибетопелагиаль |
батибенталь, батибетопелагиаль |
| глубинная(глубже 3000м) |
абиссопелагиаль, абиссобенталь, абиссобентопелагиаль |
не существует |
не существует |
Сообщества, населяющие биотопы открытого океана, исключительно разнообразны в видовом отношении, но в этой книге они рассматриваются на примере только одного их фрагмента — таксоцена рыб, или ихтиоцена. Таксоценом называют совокупность всех видов некоторой таксономической группы любой надвидовой категория входящих в состав одного биоценоза любого иерархического ранга. Необходимость в таком термине определяется тем обстоятельством, что изучение конкретных сообществ крайне редко проводится по полному состав слагающих его видов. В этом, действительно, не всегда есть нужда, так как многие особенности биоценозов могут быть исследованы и на примере отдельных таксоценов. В отечественную ихтиологическую литературу понятие «ихтиоцен» было впервые введено именно для обозначения совокупности рыб как части единого сообщества, объединяемого общим биотопом. Впоследствии в этом же биоценотическом смысле употреблялись такие термины, как «ихтиоценоз», «рыбная часть сообщества»; «рыбное сообщество», «рыбное население» и «ихтиокомплекс», из которых первые четыре должны рассматриваться как полные синонимы «ихтиоцена», а последний лишен строго детерминированного смысла.
Основные океанические ихтиоцены — пелагический на бентический (донный) — традиционно рассматриваются в качестве самостоятельных категорий. Этого нельзя сказать о бептопелагическом (придонно-наддонном) ихтиоцене, реальность существования которого над материковым склоном и ложем океана часто недооценивается, несмотря на его полную гомологию сообществу рыб неритической эпипелагиали.
При самом крупномасштабном районировании Мирового океана, основанном главным образом на распространении крупных систематических групп (ранга семейства и выше) выделяют несколько областей, объединяемых в три циркумглобальных биогеографических царства, или надобласти, — Аркто-Бореальное, Тропическое (Тропическо-Субтропическое) и Нотально-Антарктическое. В этом делении отражено наиболее серьезное из существующих различий региональных флор и фаун — обусловленное историческими причинами противопоставление холодноводных и тепловодных сообществ.
Районирование верхних и средних слоев пелагиали в пределах надобластей проводится главным образом на основе распределения отдельных видов планктона и нектона. Повторяющиеся видовые ареалы могут быть сгруппированы по сходству формы и географического положения в несколько основных типов или «географических элементов» фауны, что позволяет выделять участки среды того или иного биогеографического ранга. Наибольшая дробность районирования выявляется при анализе распространения планктонных организмов.
В целом биогеографическое районирование океанской эпи- и мезопелагиали подчинено широтной зональности, нарушаемой в той или иной степени только в приконтинентальных участках. В Мировом океане достаточно четко различаются следующие широтные зоны: 1) арктическая и 2) бореальная (с высоко- и низкобореальной подзонами), входящие в состав Аркто-Бореальной надобласти; 3) северная субтропическая, 4) тропическая (с северной центральной, экваториальной и южной центральной подзонами) и 5) южная субтропическая в составе Тропическо-Субтропической надобласти; 6) потальная и 7) антарктнческая (с низко- и высокоантарктической подзонами) в составе Нотально-Антарктической надобласти. В Индийском океане трех северных зон нет, и счет начинается тропической зоны (точнее говоря, с ее экваториальной подзоны). Деление бореальной зоны на подзоны отчетливо выражено только в Тихом океане, тогда как в Атлантике они значительно менее обособлены. Расчленение тропической зоны на экваториальную и центральные подзоны заметно главным образом по распространению планктонных видов, но благодаря эффекту экспатриации (выселения) границы этих подзон, как правило, оказываются сильно размытыми. Тропическая и субтропические зоны (выделение последних основано главным образом на особенностях распространения нектонных рыб и кальмаров) также разделены переходными полосами, населенными смешанной фауной. Еще более широкие «переходные зоны» расположены на периферии Тропическо-Субтропической надобласти, т. е. у ее границ с бореальной и нотальной зонами. Особенно хорошо такой участок, называемый «переходной зоной Южной субтропической конвергенции», выражен в южном полушарии. В некоторых систематических группах (в частности, среди макропланктонных рыб) здесь наблюдается высокий видовой эндемизм (более 30%), но в других таксоценах пелагического сообщества он почти отсутствует.
Границы всех широтных зон и подзон достаточно четко выражены в средних частях океанов и еще более резки в районах схождения течений у берегов. К таковым относятся, в частности, фронтальные зоны между субполярными и центральными круговоротами в западных частях океанов, соответствующие северной и южной границам Тропическо-Субтропической надобласти. На востоке эти границы, напротив, находятся в зонах расхождения течений и представляют собой обширные участки переходного типа, расположенные в Канарском, Бенгельском, Калифорнийском и Перуанском течениях. Субтропический зоны и центральные подзоны также сильно перекрываются в восточных частях океанов.
Глядя с палубы на однообразную поверхность океана, трудно вообразить, что где-то под килем проходят в толще воды незримые, но реальные зоогеографические границы. Еще труднее представить себе, что эти границы вполне равнозначны тем, которые разделяют столь резко различающиеся ландшафты суши - тундру и тайгу, степь и пустыню. Между тем в каждом участке пелагиали есть характерные рыбы, которые с первого взгляда распознаются специалистами. Моя основная мысль такова: морская зоогеография — точная и строгая наука, и знание ее принципов очень помогает в самых разных ситуациях.
Рыбы, живущие в эпипелагиали постоянно (со времени появления на свет до конца своего существования) предложено называть голоэпипелагическими и противопоставлять их видам меропипелагическим, в обязательном порядке проводящим в верхних горизонтах открытого океана лишь определенную часть своего жизненного цикла, и ксеноэпипелагическим, представляющим собой более или менее случайный элемент в поверхностном слое ( «Голос» по-гречески - весь, целый, «мерос» - часть, доля, «ксенос» - чуждый.).
Разделение эпипелагических рыб открытого океана между голо-, меро- и ксеноэпипелагической группировками, как правило, достаточно очевидно, хотя не абсолютно — как обычно, желание разложить природное разнообразие по полочкам вступает в противоречие с практикой жизни. Так, многие виды крупных акул (в частности, кархародон и тигровая), большая барракуда (Sphyraena barracuda), ауксиды, полосатый и желтоперый тунцы, ваху, большая корифена, по-видимому, в той или иной степени обычны и в неритической зоне, и в открытых водах (часто их называют неритоокеаническими). Не во всех случаях при современной степени экологической изученности можно четко разграничить мероэпипелагических и ксеноэпипелагических рыб.
В ихтиологии под хищниками традиционно понимают рыб, питающихся другими рыбами, и противопоставляют их мирным рыбам, поедающим беспозвоночных животных и растения. В гидробиологии принято другое содержание термина: хищником считается любое животное, потребляющее в пищу животный корм.
Скудность палеонтологического материала, значительные в ряде случаев отличия океанических таксонов рыб от родственных прибрежных форм, а также их сходство между собой, определяемое конвергенциями и параллелизмами, создают определенные трудности для воссоздания эволюционной истории рыб открытого океана. Тем не менее происхождение рецептной океанической ихтиофауны вполне очевидно: исходным материалом для ее формирования были шельфовые и неритические предковые формы. Заселение придонных глубин и открытой пелагиали, осуществлявшееся многими группами независимо друг от друга, представляется очень растянутым во времени. Датировка начала этого процесса, теряющаяся в далеком прошлом, невозможна без малооправданных спекуляций, а на примере эпипелагиали и мезобентали видно, что заселение основных океанских биотопов продолжается и поныне. В результате для каждого из этих биотопов характерна разновозрастная фауна.
Как известно, степень морфо-экологической специализации к условиям глубин, косвенно отражающая филогенетический возраст таксонов, была положена А. П. Андрияшевым в основу разделения глубоководных рыб на две группировки — древнеглубоководную и вторичноглубоководную. Такое разделение, несомненно реальное и очень наглядное в некоторых крайних случаях, все же довольно относительно и не дает возможности альтернативно определить принадлежность любой и каждой систематической группы рыб к одной из этих категорий. Представляется, что наиболее резко обособлены от ближайших родственников таксоны, возникшие в результате пелагизации, т. е. при переходе от бентического образа жизни к бентопелагическому, а затем к обитанию в безбрежной толще вод. Эти переходы носившие прогрессивный характер, сопровождались повышением общего уровня организации и соответствовали, таким образом, введенному академиком А. Н. Северцовым понятию «ароморфоз», тогда как постепенные адаптивные преобразования при увеличении глубины обитания не выходили за рамки частных приспособлений.
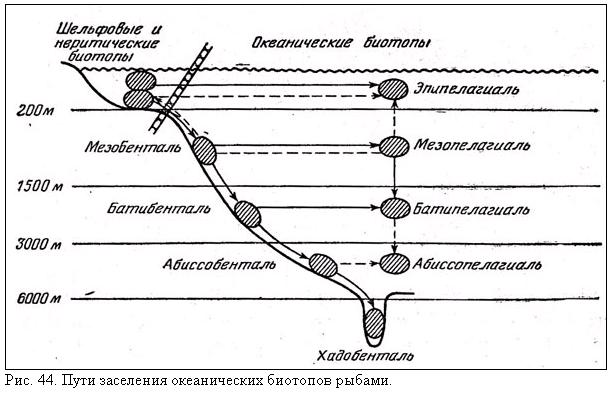
Генеральная схема заселения океанических биотопов представляется сейчас в следующем виде (рис. 44). Шельфовые предки дали начало мезобентальным рыбам, а от них последовательно произошли бати- и абиссобентальные и хадальные формы. В этом процессе очень существенная роль принадлежала вытеснению на большие глубины примитивных представителей тех или иных таксонов, не выдержавших конкуренции с более прогрессивными формами. Заселение разных горизонтов пелагиали происходило от придонных и неритических предков независимо на разных глубинах и шло главным образом по пути прогрессивной ароморфной эволюции; в формировании батипелагической фауны существенное значение имело и заселение сверху, из мезопелагиали. Картина современного биотопического распределения океанических рыб осложняется вторичным подъемом представителей некоторых групп в более поверхностные горизонты, вторичным же возвращением из открытой пелагиали к склонам.