(† Hybodontiformes Owen, 1846) Отряд Гибодонтообразные, Order † Hybodontiformes Owen, 1846 8 семейств
Материал подготовил Фактистов И. Е., 2021 г. Сайт www.fishbiosystem.ru
Гибодонтообразные (Hybodontiformes) — отряд вымерших хрящевых рыб (Chondrichthyes Huxley, 1880), относящийся к классу пластиножаберных (Elasmobranchii Bonaparte, 1838) и обладающий следующими отличительными особенностями: роговые выступы на головах самцов. Единственный отряд в инфраклассе гибодонтов (Hybodontа). Представители отряда похожи на акул и являются их ближайшими родственниками. Они жили от девона до миоцена.
Термин «гибодонт» происходит от греческих слова υβος (hybos) — «выпуклость», «горб», «горбатый» и οδοντος (odontos) — «зуб», «зубы», «зубцы». Это название было дано за форму их конических уплотнённых зубов.
Гибодонтообразные образуют группу акул, наиболее близкую к неоселахиям (Neoselachii — клада современных акул и скатов). Гибодонты различаются по конической форме зуба. Они составляли основную группу юрских акул в Европе и Северной Америке. Они сохранились до позднего мелового периода, и вымерли, возможно, из-за конкуренции со стороны других акул, хотя такие формы, как Miosynechodus, жили ещё в миоцене. Lonchidion был одним из последних гибодонтов — его характерные зубчатые тонкие шипы встречаются в пресноводных отложениях в Вайоминге вместе с окаменелостями последних динозавров, включая Тираннозавра рекса (Tyrannosaurus rex) и Трицератопса (Triceratops). Hybodontiformes идентифицируются в летописи окаменелостей преимущественно по отчетливым зубам и шипам плавников. Было известно, что они жили как в пресной, так и в солёной воде.
Гибодонтообразные классифицируются в Euselachii наряду с Xenacanthiformes, Ctenacanthiformes и Neoselachii. Род Lissodus — распространенный пример Polyacrodontidae, Hybodus — пример Hybodontidae, а Ptychodus mortoni — пример Ptychodontidae.
Впервые гибодонты были описаны в XIX веке по изолированным ископаемым зубам (Agassiz, 1837). Самые ранние останки гибодонтов происходят из карбона и включают Tristychius и других рыб из известковых песчаников в Шотландии, а также Lissodus из горных пород в Ирландии и России. Гибодонты были впервые отделены от живых акул Циттелем (1911). Хотя первые окаменелости гибодонтов относятся к каменноугольному периоду, они, вероятно, произошли от неоселахий (современных акул) в раннем девоне. Группа теперь называется гибодонтообразным включает в себя множество видов, с примерами из родов, такими как Hybodus, Acrodus, Asteracanthus, Lonchidion и Lissodus.
Образцы гибодонтообразных были извлечены из пермских отложений в Омане, что указывает на то, что гибодонты жили в океане Неотетис в течение пермского периода. Это исследование в сочетании с другими из Техаса позволяет предположить, что гибодонты были хорошо укоренившимися, а в некоторых местах и доминирующими в пермский период. В целом количество гибодонтов в перми ограничено. Первоначально предполагалось, что на разнообразие гибодонтов не повлияло значительное вымирание в конце пермского периода, вместо этого считалось, что разнообразие пермских гибодонтов уменьшилось за 50 миллионов лет до конца пермского вымирания. Однако недавние образцы, обнаруженные в Омане, предполагают, что разнообразие пермских гибодонтов распространялось до конца пермского периода, предполагая, что вымирание было более значительным, чем считалось ранее.
В окаменелостях нижнего триаса из алевролитовой пачки Вега-Фрозо (Vega-Phroso Siltstone Member) формации Серная Гора (Sulphur Mountain Formation) в Альберте (Alberta), Канада, обнаружены хорошо сохранившиеся образцы Wapitiodus aplopagus, которые пережили вымирание и были многочисленны в раннем триасе. Максимальное разнообразие гибодонтов наблюдается в триасовый период. В течение триаса и ранней юры гибодонтообразные были доминирующими селахиями, как в морской, так и в неморской среде.
При изучении окаменелостей средней юры из Англии было проанализировало 20 видов из 11 родов, предполагая, что гибодонты процветали в то время. Смена гибодонтов наблюдалась в течение средней юры, переход между четко различающимися группами, наблюдался в триасе — ранней юре и поздней юре — меловом периоде. По мере того, как неоселахии (группа современных акул) продолжали диверсифицироваться в течение поздней юры, гибодонтообразные стали менее распространёнными в открытых морских условиях, но оставались разнообразными во флювиальных и ограниченных условиях в течение мелового периода. К меловому периоду гибодонтообразные были в основном (хотя и не только) ограничены пресноводными условиями. Они оставались успешными в течение мелового периода, адаптируясь к пресноводным условиям, например, семь родов были обнаружены в пресноводных отложениях из Таиланда. Вымирание гибодонтовых акул в конце мелового периода могло быть вызвано скорее конкуренцией с другими акулами, чем ударом метеорита и вулканическими извержениями, которые были названы основной причиной этого вымирания. Большинство других акул не пострадали от вымирания в конце мелового периода, что также свидетельствует о том, что конкуренция привела к гибели гибодонтов. Тем не менее, по крайней мере, один род, Miosynechodus, по-видимому, выжил еще в конце миоцена в пресноводных отложениях Шри-Ланки.
Окаменелости зубов гибодонтообразных встречаются в отложениях от морских до речных (речные отложения). Когда они впервые появились, они населяли как морские, так и пресноводные системы. В то время как гибодонты жили в пресной воде на протяжении всего своего существования, примером гибодонтов, перемещающихся в более ограниченные условия, являются образцы средней юры, обнаруженные в лагунах и других замкнутых условиях осадконакопления. Зубы семи родов гибодонтов были описаны из пресноводной хоратской группы (Khorat Group) Таиланда, которая имеет возраст от верхней юры до нижнего мела.
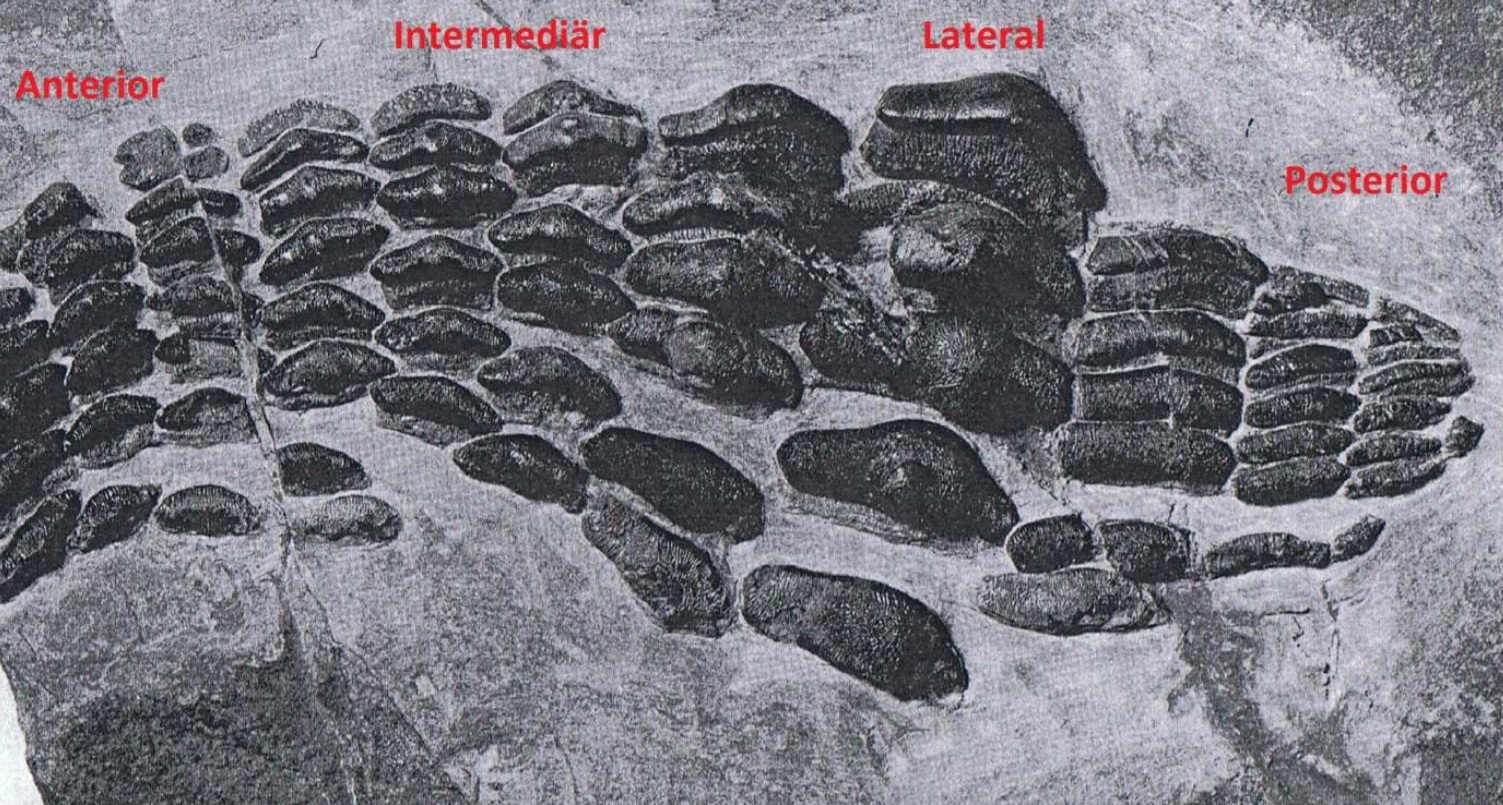
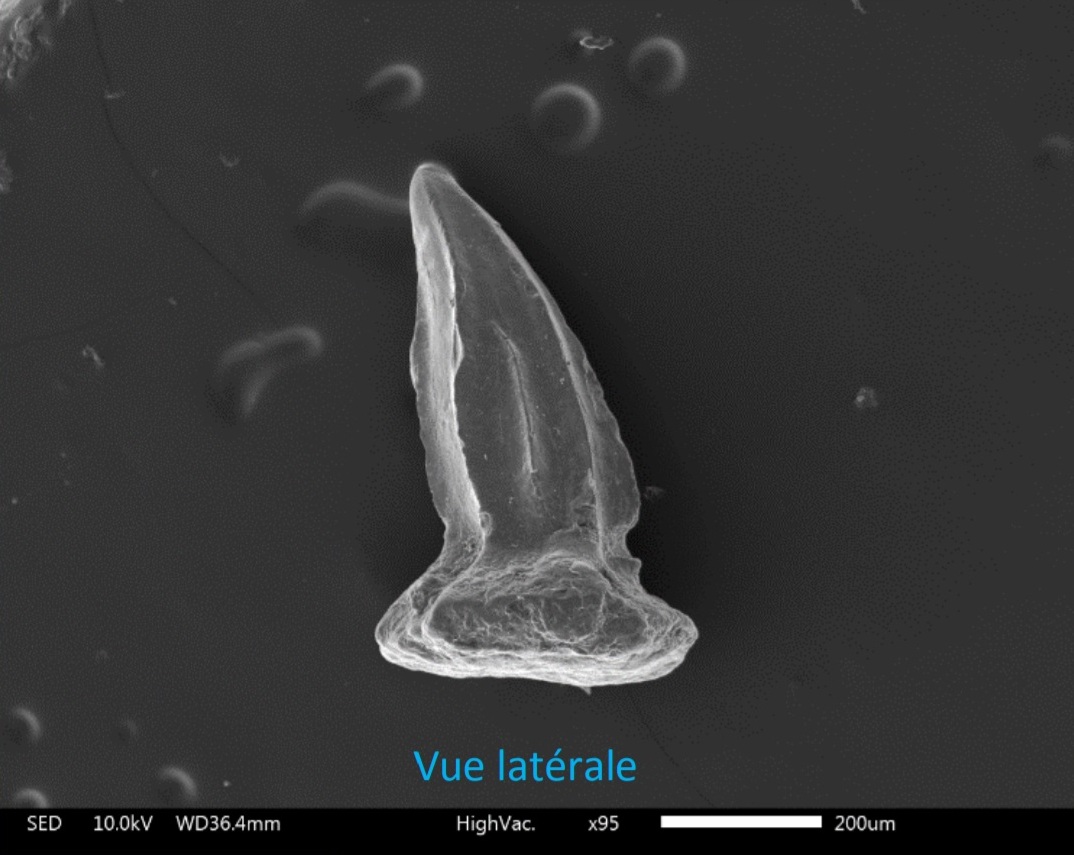
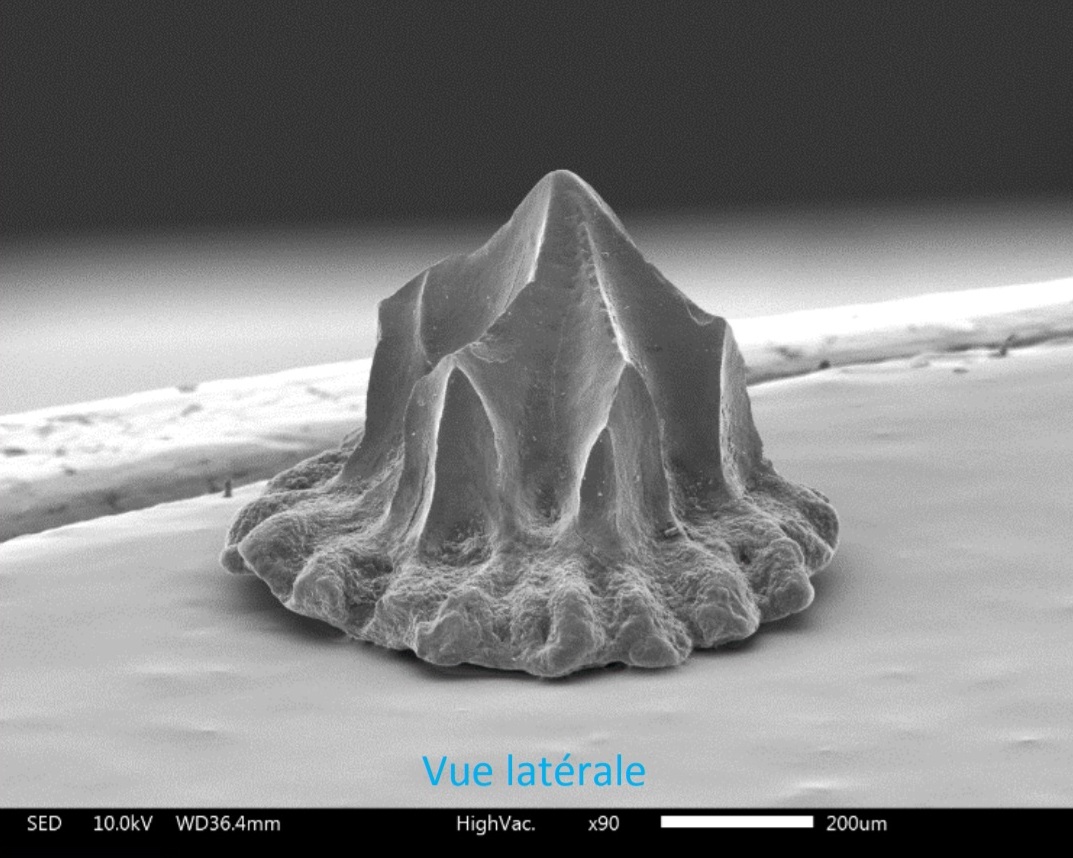
Гибодонтообразные обычно описываются и идентифицируются по размеру и форме зубов и окаменелостей шипов плавников. Считается, что у гибодонтов зубы с торчащим выступом, который выше боковых выступов. Виды гибодонтов часто определяются на основе окаменелостей их зубов, которые с большей вероятностью сохранят летопись наскальных пород, чем остатки скелета акул. В летописи окаменелостей много зубов, потому что акулы сбрасывают их на протяжении всей своей жизни, а зубы устойчивы к эрозии. Зубы гибодонтов часто сохраняются как неполные окаменелости, потому что основание зуба плохо прикреплено к коронке. Гибодонтообразные изначально были разделены на две группы в зависимости от формы зубов. В одной группе были зубы с заостренными бугорками без полости пульпы; они называются остеодонтными зубами (osteodont teeth). Другая группа имела другое расположение бугров и полость пульпы, они называются ортодонтными зубами (orthodont teeth). Например, гибодонты вида Heterophychodus steinmanni имеют зубы-остеодонты с сосудистыми каналами дентина, которые расположены вертикально параллельно друг другу, также называемые «трубчатым дентином». Коронки этих зубов-остеодонтов покрыты одним слоем эмелоида.
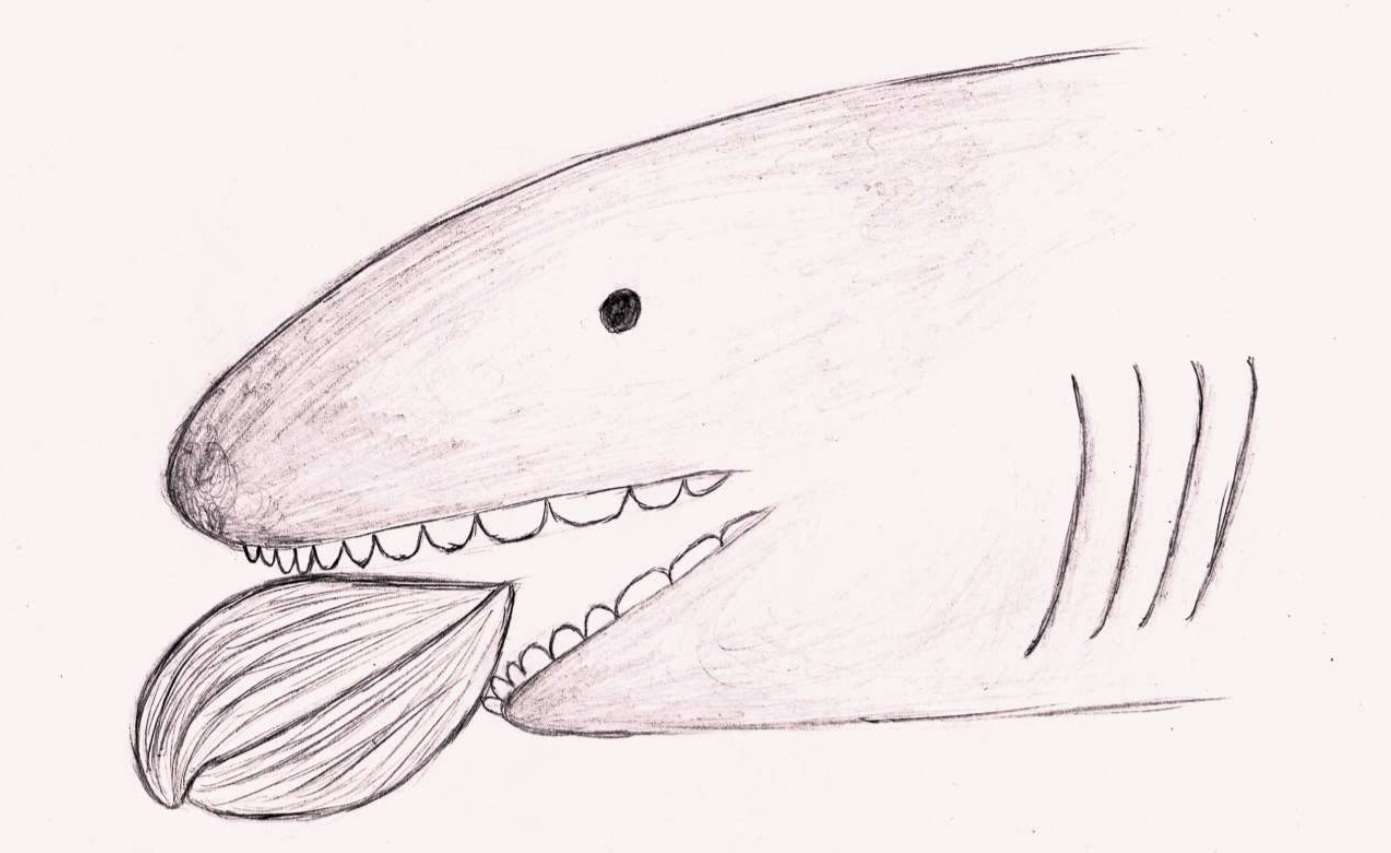
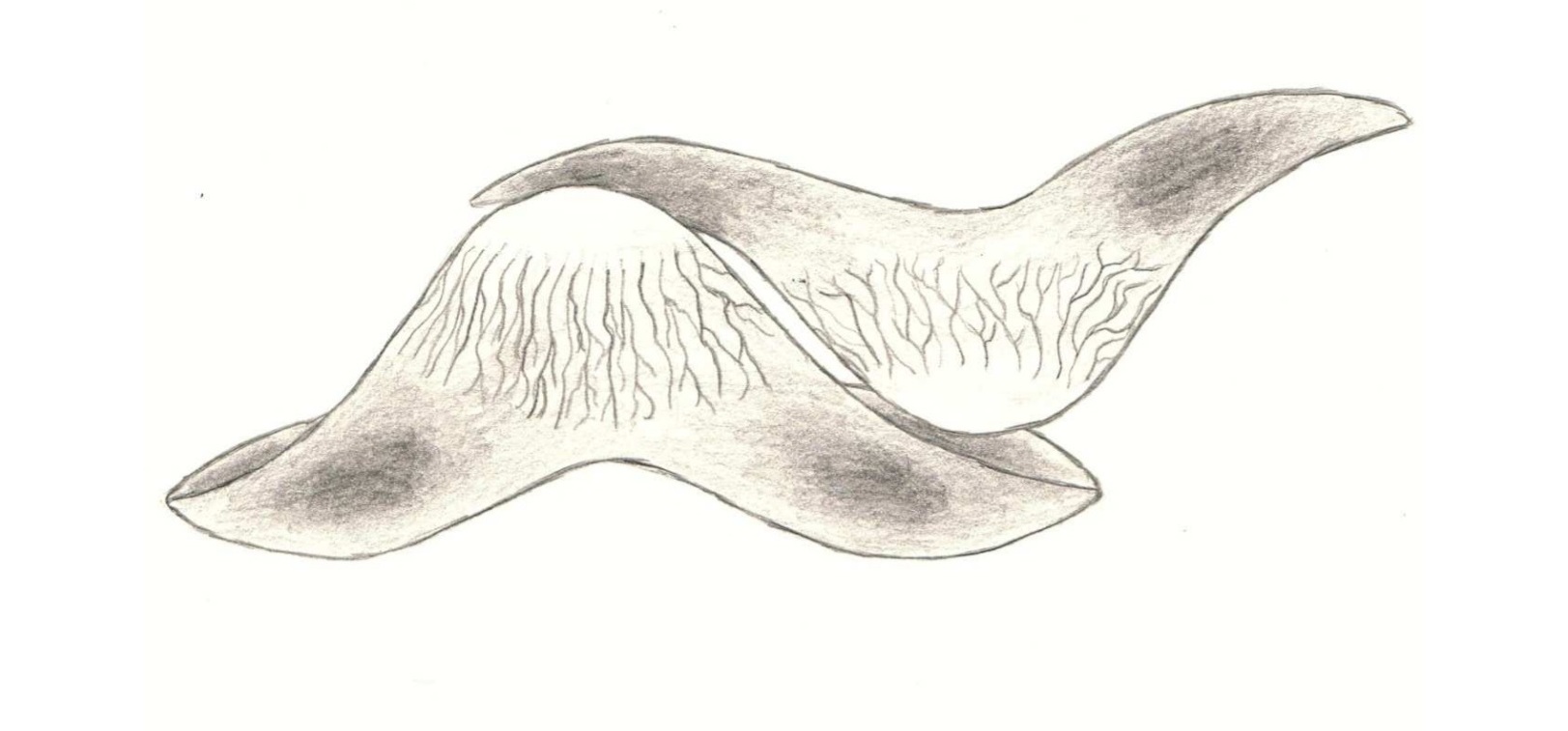
Гибодонты характеризуются наличием двух спинных плавников, каждому из которых предшествует шип определённой формы. Форма шипов плавников используется для отличия гибодонтов от других групп акул и различных видов гибодонтов. Шипы плавников имеют удлинённую форму и слегка загнуты к спине животного. У самцов гибодонтообразных были маленькие шипы на голове. У гибодонтов имелись толстые и массивные челюсти, которые различались у разных родов в зависимости от потребляемой пищи и зубов. У них были парные плавники для управления и полностью гетероцеркальный хвостовой плавник (с лопастями существенно разной длины, причём верхняя лопасть длиннее). Одна из лучших окаменелостей гибодонтов — это особь Tribodus из формации Сантана (ранний мел) на северо-востоке Бразилии. Был обнаружен трёхмерный образец, показывающий форму грудных плавников и то, как они прикрепляются к лопатке.
Гибодонты, вероятно, были медленными пловцами и использовали свои парные плавники для управления и стабилизации. Hybodus, типичный представитель гибодонтообразных, считался медлительным пловцом, но способным время от времени резко увеличивать скорость, что делало его активным хищником для быстро движущейся добычи. Гибодонты имеют самые разные формы зубов. Это разнообразие предполагает, что они использовали в своих интересах несколько источников пищи. Считается, что некоторые гибодонты, у которых были более широкие и плоские зубы, специализировались на дроблении добычи с твердым панцирем. Хорошо развитые потёртости на зубах Lissodus предполагают, что некоторые гибодонтообразные раздавливали пищу. Виды, описанные в Таиланде, имеют разную форму зубов, что свидетельствует о нескольких разных видах питания. Выпуклые зубы использовались для дробления донной добычи с твёрдым панцирем. Другие были видами, которые питались крупной мягкотелой добычей. Мало что известно о репродуктивных привычках гибодонтов. Одно исследование обнаружило множество окаменелых зубов и мешков для яиц в отложениях пресноводных озер в триасовом периоде Кыргызстана. Это место было интерпретировано как древний питомник акул на основании обилия яиц и молодых окаменелостей, а также ограниченного количества взрослых особей.
В отряд включают 8 вымерших семейств и следующие вымершие роды:
• † Семейство Acrodontidae Casier, 1959
• † Роды Acrodus Agassiz, 1838; Acrorhizodus Cappetta et al., 2006; Asteracanthus Agassiz, 1837; Bdellodus Quenstedt, 1882.
• † Семейство Distobatidae Werner, 1989
• † Роды Aegyptobatus Werner, 1989; Distobatus Werner, 1989; Glickmanodus Nessov, Mertiniene & Averianov, 1997; Reticulodus Murry & Kirby, 2002; Tribodus Brito & Ferreira, 1989.
• † Семейство Hybodontidae Agassiz, 1843
• † Роды Egertonodus Maisey, 1987; Hybodus Agassiz, 1837; Meristodonoides Underwood & Cumbaa, 2010; Pororhiza Casier, 1969; Priohybodus d'Erasmo, 1960; Thaiodus Cappetta et al., 1990.
• † Семейство Lonchidiidae Herman, 1977
• † Роды Bahariyodon Duffin, 2001; Diplolonchidion Heckert, 2004; Hylaeobatis Woodward, 1916; Isanodus Cuny et al., 2006;, Lissodus Brough, 1935; Lonchidion Estes, 1964; Parvodus Rees & Underwood 2002; Vectiselachos Rees & Underwood 2002.
• † Семейство Polyacrodontidae Glükman 1964.
• † Роды Palaeobates Meyer, 1849; Polyacrodus Jaekel, 1889; Roongodus Hairapetian & Ginter, 2009.
• † Семейство Pseudodalatiidae Reif, 1978.
• † Род Pseudodalatias Reif, 1978.
• † Семейство Steinbachodontidae Reif, 1980.
• † Род Steinbachodus Reif, 1980.
• † Семейство Tristychiidae Moy-Thomas, 1936.
• † Род Tristychius Agassiz, 1837.
Ещё два семейства могут принадлежать либо к инфраклассу гибодонтов, либо к инфраклассу неоселяхий (Neoselachii):
• † Семейство Homalodontidae Mutter et al., 2008
• † Род Homalodontus Mutter et al., 2008; Prolatodon Pla et al., 2013.
• † Семейство Ptychodontidae Jaekel, 1898.
• † Роды Heteroptychodus Yabe & Obata, 1930; Ptychodus Agassiz, 1837.
Семейство Акродонтиды — † Acrodontidae Casier, 1959, Род Акродусы — † Acrodus Agassiz, 1838 (синоним Adiapneustes Gistl, 1848; известен от пермского до палеоценового периодов): Acrodus acutus Agassiz, 1839; Acrodus alexandrae Wemple, 1906; Acrodus anningiae Agassiz, 1837; Acrodus braunii Agassiz, 1839; Acrodus cretaceus Dixon, 1850; Acrodus cuneocostatus Cuny et al., 2001; Acrodus dolloi Leriche, 1911; Acrodus emmonsi Leidy, 1872; Acrodus flemingianus de Koninck, 1863; Acrodus gaillardoti Agassiz, 1839 — типовой вид; Acrodus georgii Mutter, 1998; Acrodus hirudo Agassiz, 1839; Acrodus humilis Leidy, 1872; Acrodus illingworthi Dixon, 1850; Acrodus jaeckeli Waagen, 1895; Acrodus keuperinus Murchison & Strickland, 1837; Acrodus lateralis Agassiz, 1839; Acrodus nitidus Woodward, 1888; Acrodus nobilis Agassiz, 1838; Acrodus olsoni Johnson, 1981; Acrodus oppenheimeri Stensiö, 1921; Acrodus oreodontus Wemple, 1906; Acrodus paranense Alessandri, 1896; Acrodus scaber Stensiö, 1921; Acrodus spitzbergensis Hulke, 1873; Acrodus substriatus Schmid, 1861; Acrodus sweetlacruzensis Johnson, 1981; Acrodus undulatus Agassiz, 1839; Acrodus vermicularis Stensiö, 1921; Acrodus vermiformis Stensiö 1921, Acrodus wempliae Jordan, 1907. [L. Agassiz. 1838. Recherches Sur Les Poissons Fossiles. Tome III (livr. 11). Imprimérie de Petitpierre, Neuchatel 73-140]
Род Акродусы — † Acrodus впервые описал в 1838 году американский ихтиолог Луи Агассис (Жан Луи Родольф Агассис (Агасси) — фр. Jean Louis Rodolphe Agassiz, 1807 — 1873, американский естествоиспытатель немецкого происхождения, основоположник гляциологии (наука о природных льдах), изучал медицину в Цюрихе, Хайдельберге и Мюнхене,1831—1846 годы был профессором естествознания в университете Невшателя (University of Neuchâtel), основной из его трудов о рыбах: «Recherches sur les poissons fossiles» (5 т., Невшатель, 1833—1842, с 311 литографическими таблицами in folio), и продолжение этого труда «Monographie des poissons fossiles du vieux grès rouge du système Dévonien des Iles Britanniques» (1845), исследование о пресноводных рыбах Средней Европы остановилось на одном выпуске таблиц «Embryologie des Salmones» (1840), в 1846 году переселился в США, где полностью посвятил себя науке и обогатил коллекциями Музей сравнительной анатомии (Museum of Comparative Anatomy) при Гарвардском университете (Harvard University), в 1859 году основал (и был директором до своей смерти) Музей сравнительной зоологии (Museum of Comparative Zoology), который сейчас носит его имя, его важнейшие исследования касаются рыб, морских ежей и глетчеров, он был членом многих научных обществ, автор и соавтор описаний более 70 новых таксонов рыб, в его честь назван 21 вид рыб, а так же древнее ледниковое озеро в районе Великих озер, гора в Калифорнии, гора в штате Нью-Гемпшир, пик в Аризоне и в его родной Швейцарии, ледник в штате Монтана и на Аляске, кратер на Марсе, промонториум на Луне, мыс в Антарктиде, астероид и несколько видов животных, его сын Александр Агассис (1835—1910) — преемник отца в управлении музеем и один из выдающихся учёных своего времени).
Представители рода Acrodus — это моллюскоядные дробящезубые гибодонтные акулы. Их зубы покрыты тонким слоем эмали, сильно вытянутые, тупые, одновершинные, вдоль коронки — киль, с обеих сторон его — поперечные, иногда ветвящиеся гребешки. У этого зуба с одной стороны 3 вспомогательные вершинки, с другой — две. Акродусы существовали с позднего триаса до мела. Помимо этого наименования, их называли в своё время и адиапнестесами, и псилакантусами, и сфенонхусами, и тектодусами. Созданная модель внешнего вида была создана на основе зубов, найденных в Мушелкалке — Muschelkalk (средний триас, возраст около 237 миллионов лет), с использованием окаменелостей триаса из Монте-Сан-Джорджио — Monte San Giorgio (Швейцария) и швабских известняков (Schwäbischen Jura-Kalk) юры.
Ископаемые остатки вида Acrodus acutus известны из позднетриасовых отложений рэтского яруса Франции возрастом 205,6—201,6 млн лет. Вид был описан швейцарско-американским палеонтологом Луи Агассисом в 1839 году. Видовое название acutus с латыни означает «заострённый» и относится к зубам Acrodus acutus. Acrodus acutus был нектонным хищником, будучи, таким образом, сильным водным животным, способным противостоять течению воды. Из отложений неидентифицированного пласта исторической области Франш-Конте также известно гибодонтообразное Lissodus minimus и костные рыбы Gyrolepis albertii и Sargodon tomicus. Оттуда также описаны ископаемые остатки, вероятно принадлежащие лучепёрой рыбе Pygopterus concavus. В близлежащих отложениях были обнаружены окаменелости Acrodus acutus и Lissodus minimus, а также хрящевых ганоидов Saurichthys striatulus и Saurichthys subulatus.
Acrodus anningiae был назван Луи Агассисом в честь английского палеонтолога Мэри Эннинг (Мэри Эннинг — Mary Anning, 1799 — 1847, британский коллекционер окаменелостей и палеонтолог-любитель, известная во всем мире благодаря находкам, которые она сделала в морских окаменелостях юрского периода в скалах вдоль Ла-Манша в Лайм-Реджисе (Lyme Regis) в графстве Дорсет (Dorset) на юго-западе Англии, где она жила. Её работы способствовали коренным изменениям в научном представлении о доисторической жизни и истории Земли, которые произошли в начале XIX века).
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
%201f.jpg) |
%201p.jpg) |
%201f%20fossil.jpg) |
%202f%20fossil.jpg) |
%201f%20fossil.jpg) |
Семейство Акродонтиды — † Acrodontidae Casier, 1959, Род Акроризодусы — † Acrorhizodus Cappetta et al., 2006 (известен из нижнего мела формации Хок Круат (Khok Kruat formation) из местонахождения Кхок Пха Суам (Khok Pha Suam) близ города Шри Муанг Май (Sri Muang Mai), Таиланд): состоит из одного вида: Acrorhizodus khoratensis Cappetta et al., 2006. [H. Cappetta, E. Buffetaut, G. Cuny and V. Suteethorn. 2006. A new elasmobranch assemblage from the Lower Cretaceous of Thailand. Palaeontology 49(3):547-555]
Нижнемеловая формация Синьлун (Lower Cretaceous Xinlong Formation) в Гуанси-Чжуанском автономном районе (Guangxi Zhuang Autonomous region), Китай, которая откладывалась в неморской, речной среде, дала множество разнообразных позвоночных. Изучение зубов акул из формации Синьлун выявило присутствие зубов Acrorhizodus khoratensis, которые, по-видимому, не соответствуют положению челюсти, полученному в Таиланде. Это также дает новое понимание систематического сходства видов Гибодонтообразных.
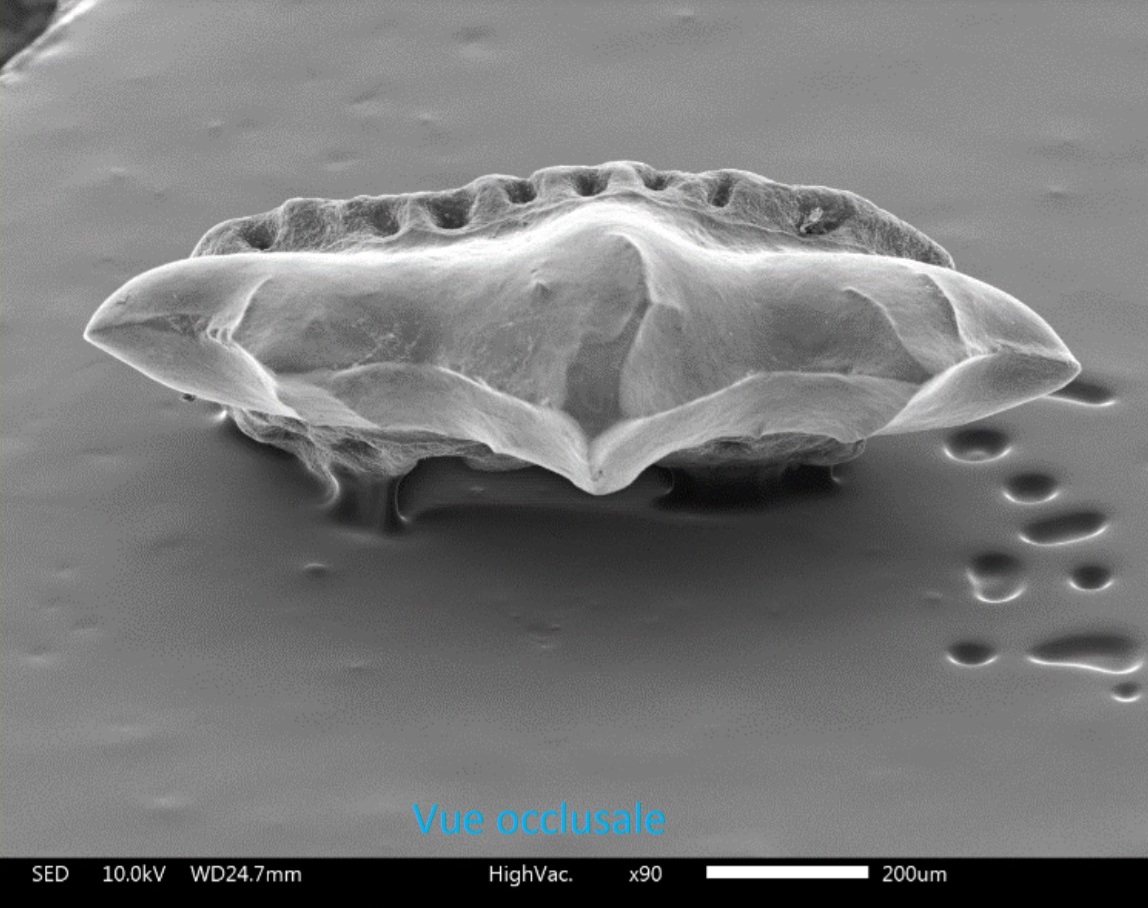
Для рода Acrorhizodus характерны мелкие зубы (общая высота менее 8 мм) с высоким квадратом короны или прямоугольной формы, с очень выпуклым губным краем, вогнутым язычным краем и прямыми боковыми краями. Чёткий гребень, параллельный губному краю зуба в окклюзионной поверхности (окклюзия — в стоматологии этот термин относится ко всякому контакту зубов верхней и нижней челюстей), разделяет её на две области, губная и язычная. С этого гребня на губной области отходят перпендикулярные складки, доходящие до края, а на язычной области — лабиолингвально (изгиб профиля в виде английской буквы S) ориентированные складки. Корень очень развитый, высокий, косой и лабиолингвально уплощенный, с большими отверстиями рядом с ним. У боковых зубов корень не такой высокий и более наклонный.
У Acrorhizodus khoratensis передние зубы симметричные, с умеренно высокой коронкой и косым и очень высоким корнем. Окклюзионный вид (рис. 8D), контур короны примерно прямоугольный. Губной край выпуклый; язычный край вогнутый и расположен параллельно губному краю. Боковые поля практически прямые. Корона имеет сильно выпуклый гребень, разграничивающий две области: язычная, косая и плоская в профиль; и удалённая (периферийная), соответствующая губным и краевым областям, которые непрерывно сливаются друг с другом. Эта периферийная область более крутая, чем язычная. Язычная область имеет орнамент, состоящий из выступающих складок в форме перевернутый V, разделенный V-образной формой, лабиолингвально ориентированной канавкой, и разделённой несколько раз спереди назад. На периферийной области имеются складки такой же конструкции, перпендикулярные посередине главному гребню, и имеющие косой край. Эти периферийные складки имеют сходную морфологию с губными складками, но более широкие, и имеющие маленькие косые гребни возникают от переднего края каждой складки. Коронка явно нависает над корнем по всему периметру зуба. Корень высокий, косой, лабиолингвально уплощенный.
Зубы Acrorhizodus khoratensis напоминают зубы Steinbachodus estheriae Reif, 1980 из Нижнего Кейпера (Lower Keuper) Германии, которые намного меньше, имеют более резцовую форму и их корона явно остроконечная на неизношенных элементах. Род Acrorhizodus может быть включен в семейство Acrodontidae, но действительные роды этого семейства (Acrodus Agassiz, 1838; Asteracanthus Agassiz, 1837 и Bdellodus Quenstedt, 1882 г.) обладают зубными рядами очень разной конструкции, с
довольно узкими передними зубами и боковыми зубами, образующими настоящие давящие и дробящие поверхности. Вполне вероятно, что род Acrorhizodus принадлежит к другому семейству, но перед его определением, необходим более качественный, сохранившийся материал.
%201f%20fossil.jpg) |
%202f%20fossil.jpg) |
%203f%20fossil.jpg) |
Семейство Акродонтиды — † Acrodontidae Casier, 1959, Род Астеракантусы — † Asteracanthus Agassiz, 1837 (известен от девона до мелового периода, от 416,0 до 70,6 миллиона лет назад): Asteracanthus acutus Agassiz, 1837; Asteracanthus aegyptiacus Stromer, 1927; Asteracanthus granulosus Egerton, 1854; Asteracanthus magnus Agassiz, 1838; Asteracanthus medius Owen, 1869; Asteracanthus minor Agassiz, 1837; Asteracanthus ornatissimus Agassiz, 1837; Asteracanthus papillosus Egerton, 1854; Asteracanthus semisulcatus Agassiz, 1837; Asteracanthus siderius Leidy, 1870; Asteracanthus somaensis Yabe, 1902; Asteracanthus tenuis Agassiz, 1838. [L. Agassiz. 1837. Recherches Sur Les Poissons Fossiles. Tome III (livr. 8-9). Imprimerie de Petitpierre, Neuchatel viii-72]
Это вымерший род акул, живший около 416—66 млн лет назад на территории США, Европы, Азии и Африки: Швейцария, Франция, Германия, Люксембург, Польша, Португалия, Британия, Япония, Нигер, Судан, Марокко, Мадагаскар и Иран. Окаменелости найдены в морских пластах от девона до мелового периода. Представители данного рода имели массивные челюсти. Зубы прямоугольные или неправильно-овальные, плоские или слабо выпуклые, без вершин, с мелкими ячейками, образованными гребешками эмали. Корни очень высокие. По-видимому, четыре серии зубов в каждой ветви челюстей и одна симфизная. Самые крупные зубы в предпоследней серии. Плавниковые шипы с многочисленными радиально-ребристыми бугорками, иногда частично сливающимися в продольные ребрышки.
%201f.jpg) |
%201f%20fossil.jpg) |
%201f%20fossil.jpg) |
%202f%20fossil.jpg) |
%203f%20fossil.jpg) |
%204f%20fossil.jpg) |
%205f%20fossil.jpg) |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
Семейство Акродонтиды — † Acrodontidae Casier, 1959, Род Бделлодусы — † Bdellodus Quenstedt, 1882 (известен из юрского периода от 183,0 до 175,6 миллиона лет назад) состоит из одного вида: Bdellodus bollensis Quenstedt, 1882. [F. A. Quenstedt. 1882. Bdellodus bollensis aus dem Posidonienschiefer von Boll. Jahreshefte des Vereins für Vaterländische Naturkunde in Württemberg 38:132-142]
Название примерно переводится как «Зуб пиявки» («Leech Tooth»). Фридрих Август фон Кенштадт (Friedrich August von Quenstadt, 1809 — 1889), немецкий геолог и палеонтолог) назвал этот род так, из-за «угольно-черного» цвета окаменелостей зубов, который он ассоциировал с пиявками. Род монотипен, содержит только вид Bdellodus bollensis, известный из тоарского времени в Германии. Он часто помещается в семейство Acrodontinae вместе с другими гибодонтами-дурофагами. Дурофагия — это пищевое поведение животных, которые потребляют организмы с твердым панцирем или экзоскелетом, такие как кораллы, моллюски с раковинами или крабы. В основном этот термин используется для описания рыб, но также используется при описании рептилий, включая ископаемых черепах.
%201f%20fossil.jpg) |
%202f%20fossil.jpg) |
Семейство Дистобатиды — † Distobatidae Werner, 1989, Род Египтобатусы — † Aegyptobatus Werner, 1989 (известен из мелового периода, ранний сеноман от 100 до 95 миллиона лет назад) состоит из одного вида: Aegyptobatus kuehnei Werner, 1989. Семейство Distobatidae при первоописании было отнесено к Myliobatiformes. Позднее переведено в Hybodontoidea.
Окаменелости были обнаружены в геологических образованиях в Формации Бахария (Bahariya Formation), Египет и датируются ранним сеноманом от 100 до 95 миллиона лет назад. Известны из разведочного бурения нефтяных скважин на большей части Западной пустыни у оазиса Бахария. Формация (четыре толщи) отложилась во время периода относительного повышения уровня моря, при этом каждое наслоение представляет собой отложения, образованные при относительном падении уровня моря. Каждая из отдельных последовательностей содержит отложения, сформировавшиеся в условиях реки, береговой линии и мелководья.
Семейство Дистобатиды — † Distobatidae Werner, 1989, Род Дистобатусы — † Distobatus Werner, 1989 (известен из мелового периода, от 99,7 до 94,3 миллиона лет назад) состоит из одного вида: Distobatus nutiae Werner, 1989.
Окружающая среда: дельтовая (1 коллекция), морская (1 коллекция), из сеномана Северной Африки в Ливии (1 коллекция), Марокко (1 коллекция, регион Менагир (Menaguir), два шестиугольных зуба, один лишь немного шире длинны, а другой намного шире длинны, слегка изогнуты, несут на своих плоских окклюзионных поверхностях ярко выпаженный орнамент). Окклюзия (стоматология) — (лат. occlusio) «всякий контакт зубов верхней и нижней челюстей». Итого: 2 коллекции, в каждой по одному экземпляру.
%201f%20fossil.jpg) |
%202f%20fossil.jpg) |
Семейство Дистобатиды — † Distobatidae Werner, 1989, Род Гликманодусы — † Glickmanodus Nessov, Mertiniene & Averianov, 1997 (известен из мелового периода, от 99,7 до 94,3 миллиона лет назад) состоит из одного типового вида — Glickmanodus rarus Nessov et al., 1997, распространённого в лиманах юго-западной окраины древней Азии позднего альба (конец раннего мела). Род назван в честь русского палеонтолога Леонида Сергеевича Гликмана (Leonid Sergeevich Glikman, 1929 – 2000) и латинского слова odus — «зуб». Название вида от латинского слова rarus — «редкий».
Местонахождение окаменелостей (боковой зуб, ЦНИГР музей, N 677/12457) — Ходжакуль (Khodzhakul), находятся в сеноманских песчаниках / аргиллитах в ходжакульской формации Каракалпакистана, Узбекистан. Glickmanodus rarus — нектонический хищник. Боковые зубы удлинённые в передне-заднем направлении, длина коронки почти в 4 раза превышает ее ширину. Корень сравнительно высокий (равен или больше высоты коронки), анаулакоризного типа, весь покрыт большим количеством беспорядочно распределённых пор (питательных отверстий) разного размера. Коронка ромбовидной формы (при взгляде сверху), уплощённая, полого приподнимающаяся в центральной части, без рельефа из эмалевых гребней на рабочей поверхности.
По строению корня и уплощенной коронке наиболее близок к Distobatus Werner, 1989 (Distobatidae) из сеномана Северной Африки (Werner, 1989) и Asiadontus Nessov, Glickman et Merteniene из апта Монголии и Киргизии (Мертинене, Несов, 1991; этот род при первоописании был отнесён к Polyacrodontidae, здесь он переводится в Distobatidae). От родов Distobatidae Glickmanodus отличается отсуствием рельефа из эмалевых гребней на рабочей поверхности коронки, от Asiadontus также сравнительно более высоким корнем. По отсутствию эмалевого рельефа сходен с Aegyptobatus Werner, 1989 из сеномана Северной Африки (Distobatidae, Werner, 1989), от которого отличается отсутствием ряда из крупных питательных отверстий на лингвальной стороне корня и относительно более низкой коронкой. От родов Acrodus Agassiz, 1837 и Asteracanthus Agassiz, 1837 (Hybodontidae, триас — мел), также имевшими уплощенные коронки, отличается отсутствием изгиба зуба в передне-задней плоскости и струйчатого эмалевого рельефа на рабочей поверхности коронки.
При первом упоминании систематическое положение рода не было определено. Позднее род был отнесен к гетеродонтоидам. Для последних характерен низкий корень (не превышающий высоту коронки) гемиаулакоризного типа, с небольшим количеством мелких питательных отверстий и одним крупным отверстием на лингвальной стороне. Glickmanodus, имеющий корень анаулакоризного типа, не может относиться к Heterodontoidea. Корень анаулакоризного типа имеется у Hybodontoidea и Hexanchiformes. Зуб Glickmanodus rarus до некоторой степени напоминает комиссурные зубы Hexanchidae. От последних отличается сравнительно более высоким корнем, коронкой, выпуклой в центральной части (не плоской) и имеющей более правильные очертания, отсутствием в основании одной из сторон коронки ряда из мелких столбиков эмали, разделенных глубокими бороздками.
Семейство Дистобатиды — † Distobatidae Werner, 1989, Род Ретикулодусы — † Reticulodus Murry & Kirby, 2002 (известен из триасового периода, от 205,6 до 201,6 миллиона лет назад) состоит из одного типового вида — Reticulodus synergus Murry & Kirby, 2002, окаменелости которого обнаружены в горизонте канала Ретия (Rhaetian channel horizon) в формации Чинл (Chinle Formation) в Аризоне, США. [P. A. Murry and R. E. Kirby. 2002. A new hybodont shark from the Chinle and Bull Canyon formations, Arizona, Utah and New Mexico. Upper Triassic Stratigraphy and Paleontology, New Mexico Museum of Natural History and Science Bulletin 21:87-106]
Reticulodus synergus — нектонический хищник. Единичные зубы вымершего Reticulodus synergus известны из слоев верхнего триаса ревуэльского возраста в Аризоне, Юте и Нью-Мексико, США. Взрослые зубы имеют длину 1—10 мм, их апикальные поверхности украшены многочисленными сетчатыми образованиями, зубцами и ямками, а в апикальном виде они выглядят частично округлыми, прямоугольными или шестиугольными — эти зубы, вероятно, позволяли этому виду быть дюрофагом. Дурофагия — это пищевое поведение животных, которые потребляют организмы с твердым панцирем или экзоскелетом, такие как кораллы, моллюски с раковинами или крабы. В основном этот термин используется для описания рыб, но также используется при описании рептилий, включая ископаемых черепах.
Ювенильные зубы имеют длину примерно 0,7—2 мм, имеют большую вариацию формы коронки, чем взрослые зубы, и обладают зубными рядами сцепляющего типа, которые облегчают захват и потребление более мягких организмов. Таким образом, зубной ряд перешел от сцепляющего типа у молодых особей к дробящему у взрослых, что позволило бы взрослым и молодым особям занимать одну и ту же среду обитания и подавило внутривидовую конкуренцию между двумя возрастными группами.
%201f%20fossil.jpg) |
%202f%20fossil.jpg) |
%203f%20fossil.jpg) |
%204f%20fossil.jpg) |
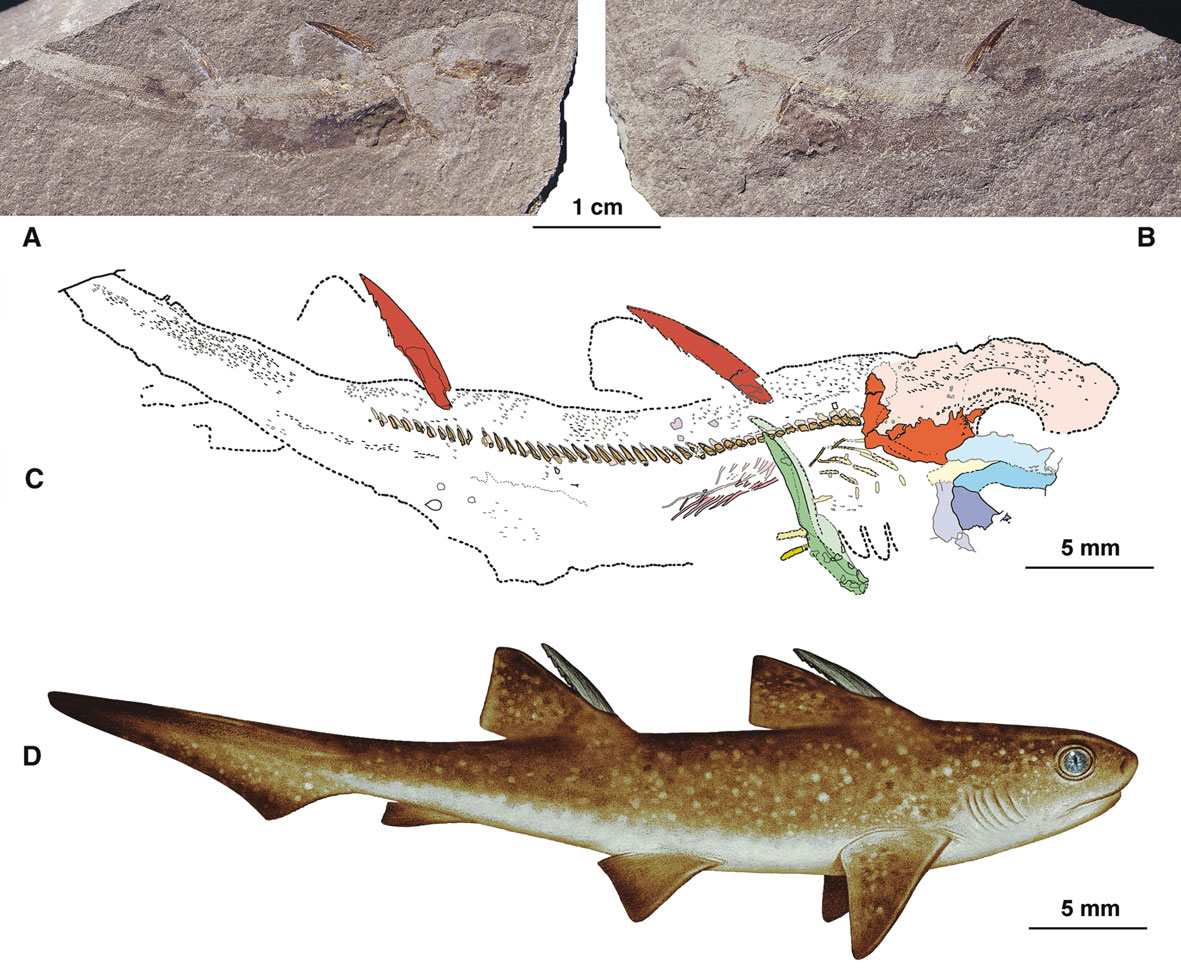
Семейство Дистобатиды — † Distobatidae Werner, 1989, Род Трибодусы — † Tribodus Brito & Ferreira, 1989 (известен из мелового периода, от 112,6 до 94,3 миллиона лет назад) состоит из одного типового вида — Tribodus limae Brito & Ferreira, 1989, частичный скелет (полная рыба без хвостового плавника) которого обнаружены в Чапада-ду-Арарипе (Chapada do Araripe), пачка Ромуальдо (Romualdo Member), которая находится в лагуне / ограниченном мелководном сублиторальном сланце в формации Сантана (Santana Formation) в Бразилии. [P. M. Brito and P. L. N. Ferreira. 1989. The first hybodont shark, Tribodus limae n.g., n.sp., from the Lower Cretaceous of Chapada do Araripe (North-East Brasil). Anais da Academia Brasileira de Ciencias 61(1):53-57]
Трибодус — вымерший род гибодонтовой акулы из формации Ромуальдо на северо-востоке Бразилии. Он жил в раннемеловом периоде (от апта до альба ). Этот род известен по расчленённым и, в некоторой степени, законченным экземплярам из местности, известной необычайной сохранностью найденных ископаемых останков. Образцы Tribodus были сохранены в трёхмерном виде, а методы кислотной обработки позволили раскрыть большую часть анатомии этой акулы. У трибодуса есть шипы на спинном плавнике и шипы на голове, но только у самцов. Tribodus limae — нектонический хищник. [Lane, Jennifer A.; Maisey, John G. (2012). "The Visceral Skeleton and Jaw Suspension In the Durophagous Hybodontid Shark Tribodus limae from the Lower Cretaceous of Brazil". Journal of Paleontology. 86 (5): 886–905.]
%201f%20fossil.jpg) |
%202f%20fossil.jpg) |
%203f.jpg) |
%204f%20skeleton.jpg) |
Семейство Гибодонтиды — † Hybodontidae Agassiz, 1843, Род Эгертонодусы — † Egertonodus Maisey, 1987 (известен из юрского и мелового периодов, от 161,2 до 99,7 миллиона лет назад): Egertonodus basanus Egerton, 1845; Egertonodus duffini Rees & Underwood, 2008; Egertonodus fraasi Brown, 1900.
Основываясь на черепных элементах, Maisey (1987) описал род Egertonodus. [Maisey, J.G., 1987. Cranial anatomy of the Lower Jurassic shark Hybodus reticulatus (Chondrichthyes: Elasmobranchii), with comments on hybodontid systematics: American Museum Novitates, n. 2878, p. 1-39.]
Зубы этого рода были в 2008 году описаны Рисом (Rees) и Андервудом (Underwood); они отметили бугорок с уплощенной губной гранью, сильным режущим краем, слабыми складками нижней поверхности коронки и множественными тонкими выступами. Они предоставили этому роду временной диапазон от батского до барремского / аптского в Англии. [Rees, J. and Underwood, C., 2008. Hybodont sharks from the English Bathonian and Callovian (Middle Jurassic): Palaeontology, v. 51, p. 117-147.]
Бурдон (Bourdon) и другие (2011) сообщили об одном зубе из сантона в Нью-Мексико, который, как они утверждали, лучше сравнивается с образцами Egertonodus, чем обычный гибодонтид из песчаника Пойнт-Лукаут — Meristodonoides. Поскольку существует только один экземпляр, и наличие этого зубного рисунка значительно расширило бы географический и временной диапазон рода, он был включен как Egertonodus. [Bourdon, J., Wright, K., Lucas, S.G., Spielmann, J.A. and Pence, R., 2011. Selachians from the Upper Cretaceous (Santonian) Hosta Tongue of the Point Lookout Sandstone, central New Mexico. New Mex. Mus. Nat. His. and Sc., Bulletin 52; 54pp.]
%201f%20fossil.jpg) |
%202f%20fossil.jpg) |
%203f%20fossil.jpg) |
%204f%20fossil.jpg) |
%201f%20fossil.jpg) |
%201f.jpg) |
 |
Семейство Гибодонтиды — † Hybodontidae Agassiz, 1843, Род Гибодусы — † Hybodus Agassiz, 1837 (известен из пермского, триасового, юрского и мелового периодов, от 295,0 до 66,0 миллиона лет назад): Hybodus acutus Agassiz, 1837; Hybodus aequitridentatus Cuny et al., 2008; Hybodus angulatus Munster, 1841; Hybodus angustus Agassiz, 1843; Hybodus antingensis Liu, 1962; Hybodus apicalis Agassiz, 1837; Hybodus austiensis Davis ,1881; Hybodus brabanticus Leriche, 1930; Hybodus bugarensis Pla et al., 2013; Hybodus butleri Thurmond, 1971; Hybodus carinatus Agassiz, 1837; Hybodus cloacinus Quenstedt, 1858; Hybodus copei Hay, 1899; Hybodus crassispinus Agassiz, 1837; Hybodus crassus Agassiz, 1837; Hybodus curtus Agassiz, 1837; Hybodus cuspidatus Agassiz, 1843; Hybodus delabechei Charlesworth, 1839; Hybodus dimidiatus Agassiz, 1837; Hybodus dorsalis Agassiz, 1837; Hybodus dubius Agassiz, 1843; Hybodus ensatus Agassiz, 1837; Hybodus ensensis Woodward, 1916; Hybodus formosus Agassiz, 1837; Hybodus grewingki Dalinkevicius, 1935; Hybodus hauffianus Fraas, 1895; Hybodus hexagonus Munster, 1841; Hybodus houtienensis Young, 1940; Hybodus huangnidanensis Wang, 1977; Hybodus inflatus Agassiz, 1843; Hybodus lawsoni Duff, 1842; Hybodus leptodus Agassiz, 1837; Hybodus levis Woodward, 1889; Hybodus leviusculus Agassiz, 1837; Hybodus longiconus Agassiz, 1843; Hybodus mackrothi Geinitz, 1861; Hybodus major Agassiz, 1837; Hybodus marginalis Agassiz, 1837; Hybodus medius Agassiz, 1843; Hybodus microdus Stensio, 1921; Hybodus mougeoti Agassiz, 1843; Hybodus multiconus Jaekel, 1889; Hybodus multiplicatus Jaekel, 1889; Hybodus nevadensis Wemple, 1906; Hybodus novus Henry, 1876; Hybodus obtusus Agassiz, 1843; Hybodus pinii Bassani, 1886; Hybodus pleiodus Agassiz, 1837; Hybodus plicatilis Agassiz, 1843; Hybodus punctatus Davis, 1881; Hybodus rapax Stensio, 1921; Hybodus raricostatus Agassiz, 1843; Hybodus reticulatus Agassiz, 1837; Hybodus sasseniensis Stensio, 1921; Hybodus shastensis Wemple, 1906; Hybodus storeri Case, 1978; Hybodus striatulus Agassiz, 1837; Hybodus subcarinatus Agassiz, 1837; Hybodus sublaevis Agassiz, 1843; Hybodus sulcatus Agassiz, 1837; Hybodus tenuis Agassiz, 1837; Hybodus undulatus Agassiz, 1843; Hybodus yohi Yang et al., 1984; Hybodus zuodengensis Yang et al., 1984. [L. Agassiz. 1837. Recherches Sur Les Poissons Fossiles. Tome III (livr. 8-9). Imprimérie de Petitpierre, Neuchatel viii-72]
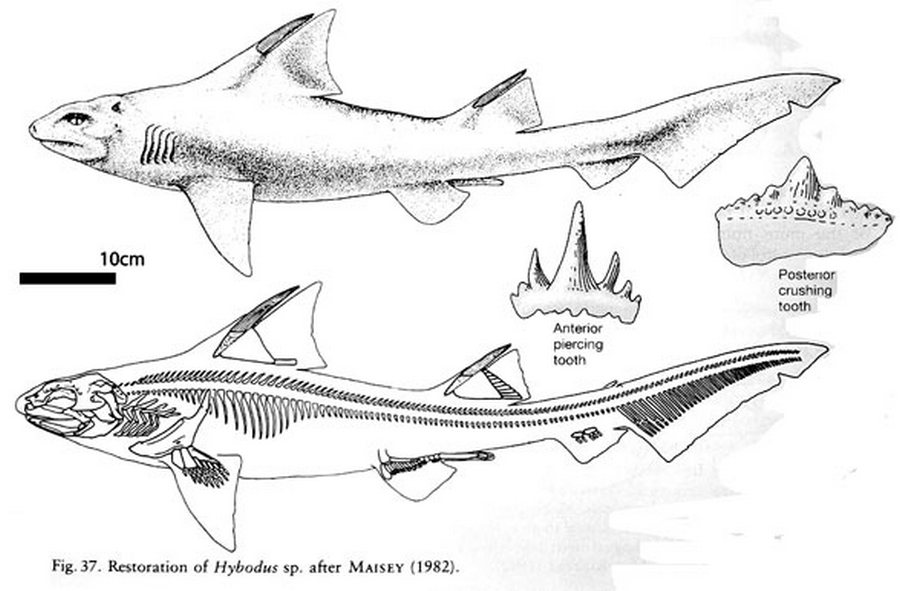
Название рода Hybodus происходит от древнегреческих слов υβος (hybos) — «выпуклость», «горб», «горбатый» и οδους (odous) — «зуб», «зубы» и указывает на форму зубов этих рыб («горбатый зуб»). Впервые появившись к концу пермского периода и исчезнув в позднем меловом периоде, в триасовый, юрский и меловой периоды гибодонты были особенно успешными и их можно было найти в мелководных морях по всему миру. По причинам, которые до конца не изучены, гибодонты вымерли ближе к концу позднего мелового периода. Виды Hybodus выростали примерно до 2 метров в длину и, как полагают, были нектоническими хищниками-оппортунистами, ловко приспосабливающимися к изменяющимся обстоятельствам.
Скелет всех гибодонтообразных состоит из хряща, поэтому плохо поддаётся окаменению. Палеонтологи, однако, обнаружили несколько хорошо сохранившихся скелетов гибодусов. У них было небольшое тело классической торпедообразной формы, и два спинных плавника, помогавших рыбам менять направление движения. Передний колючий спинной плавник мог служить рыбе средством защиты: если её собирался проглотить более крупный хищник, она поднимала спинной плавник и его острый шип вонзался в нёбо врага. Особой быстротой плавания гибодусы, по-видимому, не отличались, но их добычу составляли, в основном, мелкие и медленно передвигающиеся животные. Небольшой рот не предназначался для умерщвления крупных жертв; скорее всего, пищу гибодусов составляли мелкие животные. В челюстях сидели зубы двух типов: острые, которыми гибодусы схватывали скользких рыб, и сильно уплощённые — ими они раздавливали раковины моллюсков и панцири морских ежей. У самцов также были класперы (claspers), специализированные органы, которые непосредственно вводят сперму в самку и которые до сих пор присутствуют у современных акул.
Самые молодые окаменелости гибодусов происходят из формации «Парк динозавров» («Dinosaur Park» Formation). Они датируются периодом от 68,6 до 66 миллионов лет назад. Первые окаменелые зубы были найдены в Англии примерно в 1845 году. С тех пор зубы (и спинные шипы) были обнаружены по всему миру. Ископаемые остатки найдены в Азии, Европе, Северной Америке. Учитывая большое количество мест, где были найдены зубы, и спинные шипы гибодуса, эти рыбы были обитателями всех океанов Земли на протяжении многих десятков миллионов лет (с пермского по меловой период).
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
%201f%20fossil.jpg) |
%201f%20fossil.jpg) |
%202f%20fossil.jpg) |
 |
 |
 |
%201f%20skeleton.jpg) |
Семейство Гибодонтиды — † Hybodontidae Agassiz, 1843, Род Меристодоноидесы — † Meristodonoides Underwood & Cumbaa, 2010 (известен из мелового периода): Meristodonoides butleri (Thurmond, 1971) — Аптиан-альбский ярус Техас; Meristodonoides montanensis (Case, 1978) — Кампанский ярус из Монтаны; Meristodonoides novojerseyensis (Case & Cappetta, 2004) — Маастрихтский ярус из Нью-Джерси; Meristodonoides rajkovichi (Case, 2001) — Сеноманский ярус из Миннесоты.
Типовой вид — Meristodonoides rajkovichi из сеномана, позднего мела Миннесоты, США, который изначально был видом рода Hybodus. Этот вид, наряду с другими видами Hybodus, был передан в 2010 г роду Meristodonoides Чарли Дж. Андервудом и Стивеном Л. Кумба, которые проводили ревизионное исследование гибодонтид.
Род Meristodonoides хорошо представлен единичными зубами из Западного Внутреннего морского пути (Western Interior Seaway), а также из Казахстана, Северной Европы и других регионов Северной Америки. Они связаны с относительно мелководными морскими отложениями. Андервуд и Кумба (2010) отмечают, что диагностические характеристики включают в себя: прямой главный бугорок с небольшим наклоном в более боковых положениях; основной бугорок овальный, лабиолингвально слабо сжат, с полным, но слабым режущим краем; боковые выступы маленькие или отсутствуют, хорошо отделены от главного бугорка; хорошо дифференцированные губные складки; и низкий, хорошо васкуляризованный корень, много отверстий присутствует ниже коронки (лабиально).
 |
 |
 |
 |
 |
 |
Семейство Дистобатиды — † Distobatidae Werner, 1989, Род Пороризы — † Pororhiza Casier, 1969 (известен из мелового периода, от 113,0 до 100,5 миллиона лет назад) состоит из одного типового вида — Pororhiza molimbaensis Casier, 1969, окаменелости которого относятся к Альбианскому периоду в последовательности Болангу (Bolangu Series), Молимба (Molimba), Демократическая Республика Конго (Democratic Republic of the Congo), в 1971—1997 годах — Республика Заир (Zaire).
Среди гибодонтообразных могут быть идентифицированы два основных вида режущих зубов: с высокой тульей (Priohybodus) и с низкой тульей (Mukdahanodus, Thaiodus и Pororhiza), причем последний зубной ряд имел место только у видов, проводящих хотя бы часть своего жизненного цикла в пресных водах. Pororhiza molimbaensis имел зубной ряд режущего типа с высокими зубьями, лабиолингвально сплющенными, узко суженными в одном месте, режущие кромки слегка зазубрены, а высокий корень пересечен лабиолингвальными протоками открывающимися большими отверстиями. Наличие крупных отверстий в корне всех изученных таксонов интересны, и можно предположить значение своеобразной васкуляризации (обеспеченность кровеносными сосудами). Дело в том, что это встречается у пресноводных гибодонтов и может указывать на особую, конвергентную адаптацию к
пресноводной среде или плезиоморфный характер для это группа пресноводных гибодонтов. Действительно, почти наверняка эти таксоны обитали в пресной воде, возможно, в дельте, но не в полностью морской среде. Возможно, что развитие крупных протоков через корень — это приспособление к жизни в пресной воде.
Среди этих различных зубных рядов самыми необычными, безусловно, являются зубчатые режущие, поскольку этот вид зубных рядов, хотя и распространен у современных неоселахиевых акул, чрезвычайно редок среди гибодонтов. До сих пор было известно только три вида гибодонтов, которые развили режущие зубные ряды с зазубренными краями: Priohybodus arambourgi d 'Erasmo, 1960, известный из поздней юры — раннего мела Африки, Аравийского полуострова и Южной Америки, Pororhiza molimbaensis Casier, 1969, ограниченный альбским регионом Демократической Республики Конго (экс-Заир), и Thaiodus ruchae Cappetta et al., 1990, известным из аптского периода Таиланда и Тибета. Эти зубные ряды у гибодонтовых акул были ограничены коротким периодом времени, от поздней юры до альба, и коррелировались с высокой толерантностью к жизни в пресной воде.
Семейство Дистобатиды — † Distobatidae Werner, 1989, Род Приогибодусы — † Priohybodus d'Erasmo, 1960 (известен из юрского и мелового периода, от 150,8 до 99,7 миллиона лет назад) состоит из одного типового вида — Priohybodus arambourgi d'Erasmo, 1960.
Priohybodus arambourgi описан на основе почти 200 зубов и шипа спинного плавника из песчаника Чикла (от апта до альба, ранний мел) около Налута (Nalut) на северо-западе Ливии, в Сахаре и на Аравийском полуострове. Геологические данные не ограничиваются поздней юрой и ранним меловым периодом Африки, окаменелости также обнаружены в пачке Батови (Batoví Member) формации Такуарембо (Tacuarembó Formation), Уругвай. Материал (22 отдельных зуба и шип спинного плавника) был обнаружен в тонком костном горизонте, связанном с обильными костными фрагментами, чешуёй и зубами рыб из отряда † Semionotiformes Arambourg & Bertini 1958 и тероподных динозавров. Среди специализированных, мультикуспидальных, зубчатых и сжатых с боков коронок зубов есть патологическая коронка со вторым боковым выступом большего размера, чем первый, морфологический признак, не описанный ранее. Общее количество боковых выступов и угол между основным выступом и первой парой боковых выступов достигает более высоких значений, чем сообщалось ранее. До сих пор возраст формации Такуарембо было трудно определить, поскольку в её составе ископаемых остатков отсутствовали полезные биостратиграфические индикаторы. Нахождение Priohybodus arambourgi в этой единице значительно расширяет палеобиогеографический диапазон вида и позволяет нам предложить позднеюрский — раннемеловой возраст формации Такуарембо.
%201f%20fossil.jpg) |
%202f.jpg) |
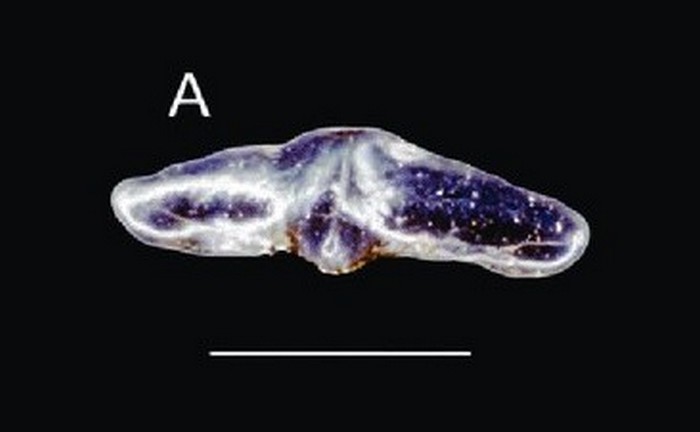
Семейство Дистобатиды — † Distobatidae Werner, 1989, Род Тайодусы — † Thaiodus Cappetta et al., 1990 (известен из мелового периода, от 125,45 до 99,7 миллиона лет назад) состоит из одного типового вида — Thaiodus ruchae Cappetta et al., 1990.
Новый род был описан из раннемеловой Хок-Круатской формации, Таиланд. Полевые работы проводились тайско-французской палеонтологической лабораторией. Экспедиция собрала дополнительный материал. Кроме описанного Thaiodus ruchae Cappetta, Buffetaut & Suteethorn, 1990, были собраны зубы двух других таксонов. Эти акулы демонстрируют широкий диапазон питания, и многие из них, по-видимому, ограничены пресноводной средой и, следовательно, вероятно, являются эндемиками плато Корат (Khorat Plateau). Тем не менее, Thaiodus и Heteroptychodus также встречаются в дельтовых и / или морских средах за пределами Таиланда, но, тем не менее, ограничены азиатским континентом.
Формация Хок Круат довольно хорошо датирована на основе позвоночных и палинологических данных. Гибодонтная акула Thaiodus в настоящее время известна только из формация Хок-Круат (Khok Kruat Formation) и формации Такена (Takena Formation) в Тибете, которая датируется апт-альбским периодом. Среда осадконакопления формации Хок-Круат считается в основном речной. Следовательно, селахи, описанные в этой статье, были, по всей вероятности, пресноводными или, в лучшем случае, эвригалинными.
Был найден зуб Thaiodus ruchae, сломанный с одной стороны, но, несмотря на это, сохранность очень хорошая. Зуб, вероятно, был симметричный и не очень широкий. Его средняя часть поднимается, образуя очень тупой куспид. Незначительно передняя кромка спускается по острие и дифференцируется на низкие, тупые и все более широкие вторично зазубренные зубцы. Рядом с краем, возникает четко очерченный, острый зубчатый бугорок с более тупым бугорком у его основания. Главный бугорок довольно шаровидный на вершине и несёт часть язычной, а иногда и губной, радиальной складки. В профиль верхняя часть губной стороны довольно прямая, но вогнутая в основании, а лингвиальная сторона выпуклая (см. рис. 3). Особенно интересен корень, более узкий, чем коронка. На лингвальной стороне (рис. 2E) он отделён от макушки глубокой поперечной бороздкой; его лингвиальное лицо довольно высокое, слегка вогнутое и наклонное при виде в профиль (рис. 3). В его основании — шесть больших круглых и выровненных отверстий. Базальный лицо (рис. 2D) плоское, очень слабо лабиолингвиально развитое, с параллельными и прямыми губными и язычными краями. Губная поверхность корня хорошо развита и наклонена; его основание перфорировано шестью отверстиями, которые соединяются с язычной стороной. На сломанной части зуба виден канал, соединяющий губное и язычное отверстия (рис. 3). Лабиально коронка нависает и полностью закрывает корень, от которого он отделён небольшой поперечной выемкой (см. рис. 2С). На языковой стороне (рис. 2E) основание коронки демонстрирует очень неровную поверхность, несущую мелкие и нерегулярно расположенные отверстия.
Материал, доступный для первоначального описания (Cappetta et al. 1990), был довольно богат, но, к сожалению, ни один из зубов не был полным, особенно на корневом уровне; поэтому было трудно определить, какая часть зуба была губной и язычной. Настоящий материал состоит из одного сломанного зуба, показывающего очень хорошо сохранившийся корень. Важные морфологические особенности корня — очень уменьшенная базальная поверхность и большие отверстия, открывающиеся губно и лингвиально. В зубных рядах зубы были тесно связаны от одного ряда к другому, острая базальная часть на губной стороне (рис. 3), сочленяется в поперечном направлении, бороздка зуба расположена чуть лабиально в ряду (Рис. 4). Хорошая сохранность корня позволяет по новому интерпретировать ориентацию зубов. Тот факт, что корень более развит лингвиально у пластиножаберных рыб, похоже, что то, что мы считали губной стороной (в Cappetta et al., 1990) становится лингвиальной и наоборот. Интересно отметить, что род гибодонтоидов (даже если его отнесение к Hybodontidae s.s. сомнительно) из пресных вод нижнего мела месторождения Демократической Республики Конго (Заир), Pororhiza Casier, 1969, также показывает корень с большим отверстиями. Эту же особенность можно наблюдать у других гибодонтов из Хок Пха Суам: Hybodus sp., Heteroptychodus aff. steimanni и Acrorhizodus khoratensis gen. et sp. nov. Возможно, что развитие крупных протоков через корень — это приспособление к жизни в пресной воде.
%201f%20fossil.jpg) |
%202f.jpg) |
%203f.jpg) |
%204f%20fossil.jpg) |
Семейство Лонхидииды — † Lonchidiidae Herman, 1977, Род Бахарииодоны — † Bahariyodon Duffin, 2001 (известен из мелового периода) состоит из одного типового вида — Bahariyodon bartheli Werner, 1989.
Одиночные зубы Bahariyodon bartheli (Werner 1989, Duffin 2001) из верхнего сеномана (поздний мел) Египта были обнаружены в местонахождениях Дуйра (Douira) и Дар-эль-Кариб (Dar el Karib) в формации Дуйра (Douira Formation) из осадочного комплекса группы Кем Кем (Kem Kem Group), Северная Африка.
Семейство Лонхидииды — † Lonchidiidae Herman, 1977, Род Диплолонхидионы — † Diplolonchidion Heckert, 2004 (известен из триасового периода, от 221,5 до 205,6 миллиона лет назад) состоит из одного типового вида — Diplolonchidion murryi Heckert, 2004. [A. B. Heckert. 2004. Late Triassic microvertebrates from the lower Chinle Group (Otischalkian-Adamanian: Carnian), southwestern U.S.A. New Mexico Museum of Natural History and Science Bulletin 27:1-170] По други данным род † Diplolonchidion Heckert, 2004 отнесён к семейству † Polyacrodontidae Glükman, 1964.
Типовой образец обнаружен в песчанике / конгломерате Норианского канала (Norian channel) в формации Тековас (Tecovas Formation) в Техасе, США. Diplolonchidion murryi — нектонический хищник.
Семейство Лонхидииды — † Lonchidiidae Herman, 1977, Род Хилаеобатисы — † Hylaeobatis Woodward, 1916 (известен из мелового периода, от 112,6 до 99,7 миллиона лет назад) состоит из одного типового вида — Hylaeobatis problematica Woodward, 1916. [A. S. Woodward. 1916. The fossil fishes of the English Wealden and Purbeck Formations. Palaeontographical Society, London 1915: 1-48]
Окаменелости обнаружены в английских формаций Wealden и Purbeck. Зуб типового образца хранится в Йоркском музее (York Museum). Орнамент на зубах Hylaeobatis состоит из довольно сильной сетчатой структуры. Зубы Hylaeobatis также могут быть отделены от зубов Lissodus по отсутствию окклюзионного гребня и губного выступа в боковых зубах. Зубы Hylaeobatis problematica поперечно удлинённые, от овального до прямоугольного в окклюзионном виде; окклюзионная поверхность украшен сетчатыми складками, несколько вздутый, но без дифференцированных бугорков; губной бугорок почти отсутствует на всех зубах; корневой массив с хорошо развитыми отверстиями на лингвиальной стороне.
%201f%20fossil.jpg) |
%201f%20fossil.jpg) |
%202f%20fossil.jpg) |
Семейство Лонхидииды — † Lonchidiidae Herman, 1977, Род Исанодусы — † Isanodus Cuny et al., 2006 (известен из мелового периода, от 130,0 до 122,46 миллиона лет назад) состоит из одного типового вида — Isanodus paladeji Cuny et al., 2006. [G. Cuny, V. Suteethorn, S. Kamha, E. Buffeteut, and M. Philippe. 2006. A new hybodont shark assemblage from the Lower Cretaceous of Thailand. Historical Biology 18(1):21-31]
Название рода Isanodus происходит от названия северо-восточной части Таиланда Isan — Исан и греческого слова οδους (odous) — «зуб», «зубы». Вид назван в честь Паладея Срисука (Paladej Srisuk) «за его неустанный труд по сбору окаменелостей в Таиланде, и который пожертвовал часть типовой серии Исследовательскому центру динозавров Сахсакхана (Sahatsakhan Dinosaur Research Centre)». Зуб типового образца обнаружен в местечке Пху Фан Тонг (Phu Phan Tong), которое находится в барремских / аптских пойменных песчаниках / известняках формации Сао Хуа (Sao Khua Formation) в Таиланде. Isanodus paladeji — нектонический хищник, обитающий в «пойменных» и морских средах.
Диагноз: Сильная, не градиентная моногнатическая гетеродонтия, аналогичная у Lissodus nodosus с четырьмя типами зубов: передними, переднебоковыми, заднебоковыми и задними; наличие лабиального штифта; главный бугорок пирамидальный с треугольным основанием, очень низкий у предполагаемых заднебоковых зубов; орнамент, состоящий из продольного гребня с одним мезио-дистальным V-образным губным гребнем и до четырёх мезио-дистальных V-образных язычных гребней; наличие гранул на некоторых предполагаемых передних и переднебоковых зубах; короткие лабиолингвиально ориентированные гребни в некоторых предполагаемых переднебоковых и заднебоковых зубах, берущие начало от мезиодистальных гребней; на мезиальной и дистальной частях переднебоковых и заднебоковых зубов до четырех хорошо развитых лабиальных узлов. Средние размеры зубов (в мм): 2,50 x 2,00.
В 2016 году находки из места Фу Фан Тонг (Phu Phan Thong) нижнего мела, провинция Нонг Буа Лам Фу (Nong Bua Lam Phu Province), северо-восток Таиланде, дали десять морфотипов зубов пресноводных акул рода Isanodus, которые позволяют провести ревизию этого рода. Два новых морфотипа, относящиеся к переднебоковым и заднебоковым зубам, обладают общими признаками с передними и задними зубами Isanodus paladeji, тогда как его бывшие переднебоковые и заднебоковые зубы, по-видимому, принадлежат новому виду, Isanodus nongbualamphuensis sp. nov. Рисунок орнамента на некоторых зубах Isanodus paladeji и Isanodus nongbualamphuensis очень похож на тот, который наблюдается у рода Heteroptychodus, что предполагает близкое родство этих двух родов.
%201f%20fossil.jpg) |
%202f%20fossil.jpg) |
Семейство Лонхидииды — † Lonchidiidae Herman, 1977, Род Лиссодусы — † Lissodus Brough, 1935 (известен от каменноугольного периода до мелового периода, от 345,3 до 66,0 миллиона лет назад): типовой вид — Lissodus africanus Broom, 1909; Lissodus angulatus Stensiö, 1921; Lissodus cassangensis Teixeira, 1956; Lissodus cristatus Delsate & Duffin, 1999; Lissodus guenneguesi Delsate, 2003; Lissodus hasleensis Rees, 1998; Lissodus johnsonorum Milner & Kirkland, 2006; Lissodus leiodus Woodward, 1887; Lissodus leiopleurus Agassiz, 1839; Lissodus lepagei Duffin, 1993; Lissodus levis Woodward, 1887; Lissodus minimus Agassiz, 1839; Lissodus wardi Duffin, 1985; Lissodus xiushuiensis Wang et al., 2007.
Lissodus — род акул, очень похожий на представителей рода Hybodus, хотя и имеет немало отличий от него. Как и в случае с Hybodus, Lissodus имел округлую морду, четыре рога, шипы на спинных плавниках и хвост с очень длинной вершиной. Но он отличался от Hybodus тем, что его брюшные плавники были бескостными, а его зубы были очень плоскими и не имели зазубренного края, что указывало на то, что их наверняка использовали для раздавливания твердой добычи, такой как моллюски. Удлинённая форма его тела говорит нам, что он, должно быть, жил на рифах и в очень мелководных морях, даже в устьях рек. Считается, что это животное могло жить и в пресной воде. Представитель рода Lissodus был около 15 сантиметров в длину и имел плоские зубы, которые он использовал для поедания моллюсков.
Типовой вид — Hybodus africanus Broom, 1909 из скифа, раннего триаса Беккеровского Краала (Bekker's Kraal), Южная Африка. Челюсти глубокие, нижняя челюсть сужается кпереди; передние зубы с центральный бугорок, окклюзионный гребень и губной бугор от умеренного до хорошо развитого; окклюзионная поверхность губного бугорка плавно наклонена к основанию коронки; форма коронки почти треугольной формы; боковые зубы ниже, крупнее, мезио-дистально
расширенны; бугорки, окклюзионный гребень, губной бугор развиты слабо; корень лингвально наклонный, ниже макушки, не такой объемный; одиночный, строго горизонтальный ряд мелких круглых отверстий около соединения коронки и корня; базальная пластинка головных шипов «Т-образная» с окончательно расширенными долями.
В род Lissodus включены виды Lissodus angulatus (Stensiö, 1921) из скифа Шпицбергена; Lissodus africanus (Broom, 1909) из скифа Южной Африки; Lissodus cassangensis (Teixeira, 1956) из скифа Анголы; Lissodus cristatus Delsate & Duffin, 1999 из анисовского острова Люксембурга; Lissodus hasleensis Rees, 1998 из г. Плиенсбахский, Дания; Lissodus leiodus (Woodward, 1887) из бата Англии; Lissodus leiopleurus (Woodward, 1889) из бата Англии и Шотландия; Lissodus lepagei Duffin, 1993 из Люксембурга; Lissodus levis (Woodward, 1887) из альба Англии; Lissodus minimus (Agassiz, 1839) из ретиана Англии, Германии и Бельгии; Lissodus nodosus (Seilacher, 1943) из среднего и позднего триаса Германии; Lissodus wardi Duffin, 1985 из бата Англии. Кроме того, плохо сохранившийся фигурный зуб из кимериджа северной Франции, по-видимому, является переднебоковым зубом Lissodus.
Зубы Lissodus легко поддаются общей морфологии и отделимы от родов Lonchidion и Parvodus из-за низких и широких зубов, так как в отличие от более высоких и изящных зубов у двух последних родов. В пределах Lonchidiidae, зубы Lissodus больше напоминают зубы Hylaeobatis и Vectiselachos. Однако наличие хорошо разграниченных бугров и боковых выступов у Lissodus отделяет зубы этого рода от двух других. Орнамент короны тоже отличается. Зубы Lissodus обычно имеют слабые складки, покрывающие большая часть коронки, а зубы Vectiselachos украшены сильными складками и гранулами. Орнамент на зубах Hylaeobatis состоит из довольно сильной сетчатой структуры. Зубы Hylaeobatis также можно отделить от зубов Lissodus по отсутствию
окклюзионного гребня и губного выступа в боковых зубах.
%201f%20fossil.jpg) |
%201f%20fossil.jpg) |
%201f%20fossil.jpg) |
%202f%20fossil.jpg) |
 |
 |
 |
 |
 |
 |
Семейство Лонхидииды — † Lonchidiidae Herman, 1977, Род Лонхидионы — † Lonchidion Estes, 1964 — синоним — Priscavolucris Gomez Pallerola, 1979 (известен от триасового периода до мелового периода, от 221,5 до 66,0 миллиона лет назад): Lonchidion anitae Thurmond, 1971; Lonchidion babulskii Cappetta & Case, 1975; Lonchidion breve Patterson, 1966; Lonchidion crenulatum Patterson, 1966; Lonchidion delsatei Guennegues & Biddle, 1989; Lonchidion derenzii Manzanares et al., 2017; Lonchidion griffisi Case, 1987; Lonchidion humblei Murry, 1981; Lonchidion indicus Yadagiri, 1986; Lonchidion inflexum Underwood & Rees, 2002; Lonchidion khoratensis Cuny et al., 2006; Lonchidion marocensis Duffin & Sigogneau-Russell, 1993; Lonchidion microselachos Estes & Sanchíz, 1982; Lonchidion monteschi Gomez Pallerola, 1979; Lonchidion selachas Estes 1964; типовой вид — Lonchidion selachos Estes, 1964; Lonchidion striatum Patterson, 1966; Lonchidion weltoni Duffin, 1985. [R. Estes. 1964. Fossil vertebrates from the Late Cretaceous Lance Formation, eastern Wyoming. University of California Publications in Geological Sciences 49:1-187]
Второй род гибодонтов с мелкими зубами, Lonchidion, был описан Эстесом в 1964году на основе отдельных зубов и связанных с ними шипов плавников и головных шипов Lonchidion selachos Estes, 1964, из формации Maastrichtian Lance верхнего мела в Вайоминге, США. Паттерн гетеродонности Lonchidion selachos не был полностью понят Эстесом, которые включили орректобидные зубы (см. Herman, 1977) в качестве симфизов. Lonchidion был широко признан как оправданный род, и несколько новых видов были описан между 1964 и 1985 гг. (Patterson, 1966; Thurmond, 1971; Cappetta and Case, 1975; Murry, 1981; Estes и Sanchíz, 1982). О останках скелетов Lonchidion не известно, за исключением нескольких неполных скелетов из нижнего мела Испания, описываемая как «Lissodus palustris» Gomez Pallerola, 1992 и, возможно, принадлежащий Lonchidion.
Типовой вид — Lonchidion selachos Estes, 1964 из Маастрихтской формации (Maastrichtian Lance
Formation), образованой в позднем меловом периоде восточного Вайоминга, США. Зубы у этого вида изящные, лабиолингвиально узкие; основной куспид низкий, но маркированные, до двух пар зубцов; губной бугорок узкий, часто параллельный,
двусторонний, сильно развитый; дистальные части коронки заострены, часто образуют дистальные
пара боковых выступов; корень обычно шире самой нижней части коронки; губная сторона корня сильно вогнута; небольшие круглые отверстия, расположенные неравномерно рядом с коронкой; базальная пластинка головных шипов с ярко выраженной «стрелкообразной» формой.
Позже были включены виды — Lonchidion anitae Thurmond, 1971 из апт—альбского периода
Техаса, США; Lonchidion babulskii Cappetta and Case, 1975 из кампана Нью Джерси, США; Lonchidion breve Patterson, 1966 из валанжинско-барремского периода Англии; Lonchidion crenulatum (Patterson, 1966) из берриас-валанжинского периода Англии; Lonchidion delsatei (Guennegues and Biddle, 1989) из тоаров Франции; Lonchidion griffisi (Case, 1987) из кампана штата Вайоминг, США; Lonchidion humblei Murry, 1981 из карнийского периода Техас, США; Lonchidion inflexum Underwood and Rees, в печати из берриаса
Англия; Lonchidion marocensis (Duffin, Sigogneau-Russell, 1993) из берриаса Марокко; Lonchidion microselachos Estes and Sanchiz, 1982 из баррем-апта Испания; Lonchidion selachos Estes, 1964 из маастрихта штата Вайоминг, США; Lonchidion striatum Patterson, 1966 год из готеривско-барремского периода Англия; Lonchidion weltoni (Duffin, 1985) из сеномана штата Орегон, США.
Так как зубы Lonchidion palustris Gomez Pallerola, 1992 крайне слабо изучены, этот вид в настоящее время не представлен. Кроме того, два безымянных виды "Lissodus" были обозначены Велтоном и Фаришем (1993) и один — Каппеттой и Касе (1999), хотя они могут быть синонимами. Эти позднемеловые находки судя по всему, это Lonchidion, но от каждого вида были обнаружены только один или два зуба. Есть также запись о зубах Лонхидиона из Muschelkalk (ладинского периода) Крайльсхайм в Германии (Паттерсон, 1966:331). Ряд зубов карнийского происхождения Вирджиния, США (Johansson, 1992), имеют морфологию, очень близкую к типовому виду Lonchidion.
Изящный внешний вид четко разделяет зубы Лонхидиона от таковых у родов Lissodus, Vectiselachos и Hylaeobatis. Зубы Парводуса такие же изящные (тонкие), как зубы Lonchidion, но они всегда имеют довольно высокие боковые выступы, как в отличие от мельчайших у Лонхидиона. Выступ короны крупный у Lonchidion, но у Parvodus почти отсутствует, и у последнего имеется губная выпуклость.
Зубная морфология Lonchidion неожиданно немного изменилась в течение длительного периода времени. Некоторые виды имеют морфологию, близкую к морфологии типового вида и различаются в первую очередь количеством орнаментов, а иногда и гетеродонтией. (Гетеродонтизм (или гетеродонтность, также гетеродонтия) — наличие у животного зубов, различающихся по форме и функциональному назначению.) У самого раннего вида, Lonchidion humblei, боковые зубы более удлинённые и асимметричны, чем у любых более поздних видов этого рода, но в остальном похожи. Связанные между собой головные шипы имеют морфологию, очень похожую на шипы «приговорённой стрелковидной формы (convict arrow shaped)» (Antunes et al. 1990) у типового вида. Это только два вида, у которых обнаружены колючки.
Единственный юрский представитель рода — Lonchidion delsatei известен только из двух плохо изученных и плохо сохранившихся зубов. Однако форма губного выступа и общая форма,
в том числе изящный внешний вид, делает эти зубы типичными для Lonchidion. Два вида из раннемелового периода, Lonchidion breve и Lonchidion microselachos, имеют морфологию, очень похожую на Lonchidion selachos. Вертикальная штриховка хорошо развита у Lonchidion marocensis и Lonchidion striatum, и в меньшей степени у Lonchidion inflexum. У Lonchidion marocensis зубы немного шире, приближающиеся по морфологии к определённым видам, отнесеным к роду Polyacrodus. В дистальной и мезиальной части у Lonchidion inflexum коронки наклонены лингвально, зубы имеют V-образную форму на окклюзионной проекции. Окклюзионный гребень на зубах Lonchidion crenulatum зубчатый, а зубы несколько более остроконечные, чем у типового вида. Наличие вупуклости также характеризует зубы Lonchidion anitae; они также обладают крайним губным бугорком. Боковые зубы Lonchidion weltoni имеют язычную и губную выпуклости, а передние зубы больше похожи на типовые. Зубы Lonchidion babulskii редко имеют хорошую сохранность, они также тонкие. Другой позднемеловой вид — Lonchidion griffisi, характеризуется зубчатым окклюзионным гребнем, но во всех других аспектах аналогичен типовому виду. Изношенные зубные коронки, похоже, имели лонхидионоподобную морфология, основанную на форме губного выступа. Зубы Lonchidion могут не иметь остеодентина, как показано у Lonchidion breve. Однако в лабиолингвиально узких зубах Lonchidion, может просто не хватить места, чтобы включить сердцевину остеодентина и это не может быть таксономически полезным для мелкозубых акул.
Известная стратиграфическая летопись Lonchidion относится к ладинскому периоду от среднего триаса до маастрихта, позднего мела. Точное стратиграфическое положение зубов из Крейлсхейма (Patterson, 1966:331) неизвестно. Акулы этого рода были особенно распространены в ранний меловой период и особенно в неморской среде. Отсутствие находок Lonchidion через большую часть юрского периода может быть артефактом, и исследование подходящих горизонтов могло бы, вероятно, расширить наши знания о роде. Акулы этого рода имеют способность переносить широкий диапазон солёности. Однако за время жизни рода акулы были более разнообразны в неморской среде, и только в поздний меловой периодпоздний меловой период, что они встречаются в полностью морских условиях.
%201f.jpg) |
%202f%20fossil.jpg) |
 |
 |
 |
 |
 |
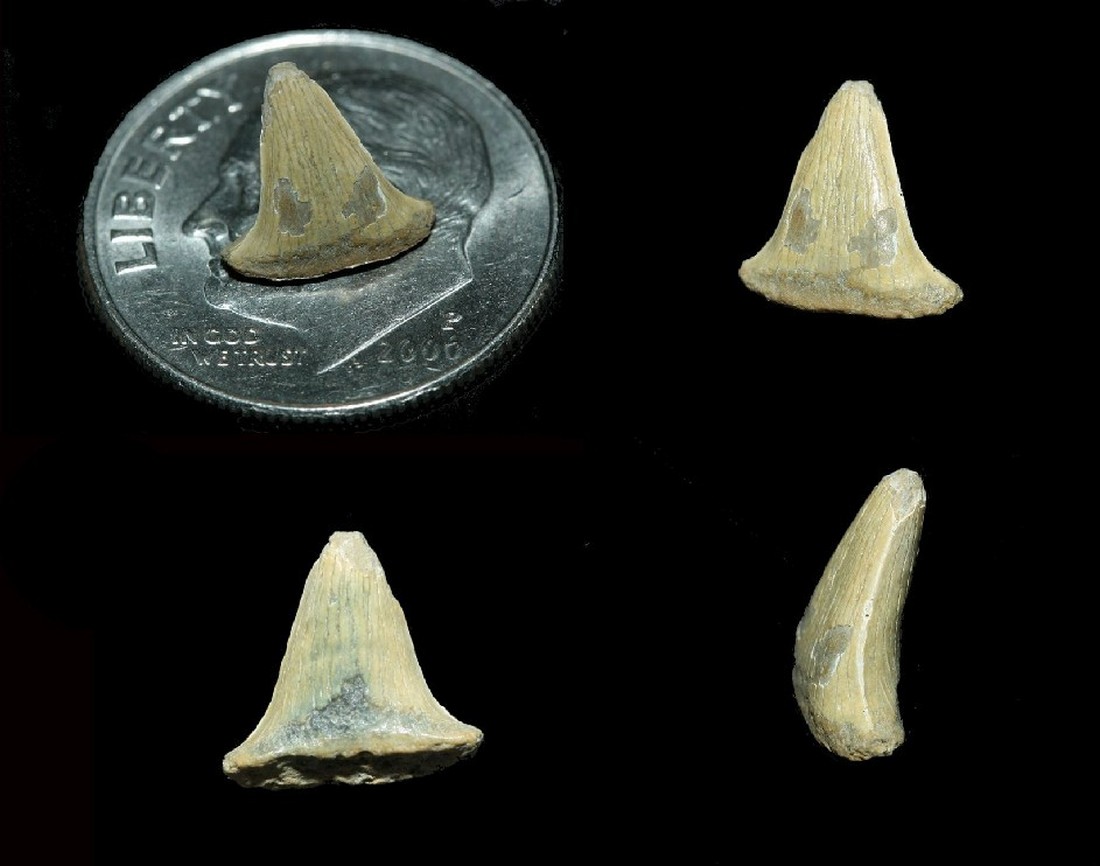
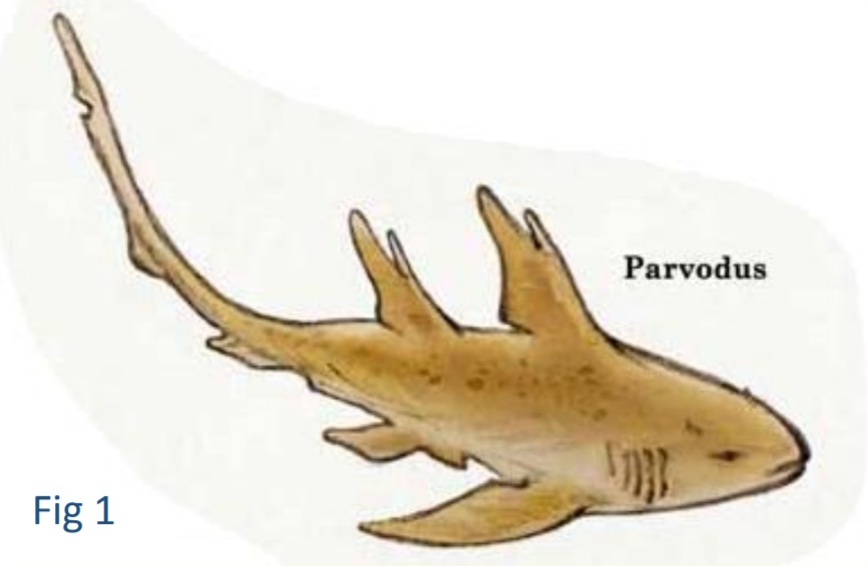
Семейство Лонхидииды — † Lonchidiidae Herman, 1977, Род Парводусы — † Parvodus Rees & Underwood 2002 (известен от триасового периода до мелового периода, от 247,2 до 112,6 миллиона лет назад): Parvodus curvidens (Duffin & Thies, 1997); Parvodus pattersoni Duffin, 1985; Parvodus rugianus Ansorge, 1990. [J. Rees and C. Underwood. 2002. The status of the shark genus Lissodus Brough, 1935, and the position of nominal Lissodus species within the Hybodontoidea (Selachii). Journal of Vertebrate Paleontology 22(3):471-479]
Название рода происходит от латинского слова parvus — «небольшой», «маленький» и греческого слова οδους (odous) — «зуб», «зубы». Типовой вид Lissodus rugianus Ansorge, 1990 из раннего мела г. Рюген, северная Германия.
Эти лонхидиидные акулы с мелкими зубами; передние зубы с двух сторон симметричные; бугорки передних зубов умеренно высокие, низкие, но хорошо разграничены в боковых частях; губной бугорок хорошо развит и округлён, часто поддерживается слабым лабиальным корневым упором; зубы очень изящные с окклюзией (Окклюзия — «всякий контакт зубов верхней и нижней челюстей».); корень довольно
низкий, наклонен к языку.
В род Parvodus включены виды — Parvodus curvidens (Duffin, Thies, 1997) из кимериджского периода Германии; Parvodus pattersoni (Duffin, 1985) из бата Англии и Шотландии; Parvodus rugianus (Ansorge, 1990) из берриаса-валанжина Германия, Англия, Дания и Швеция. Не исключено, что гетеродон «Лонхидион» Patterson, 1966 из валанжина Англии также следует отнести к этому роду, но этот вид малоизвестен, и только голотип и несколько других зубов двух паратипов могут быть включены и представлять другие виды (Underwood and Rees). Delsate и Duffin (1993) описали Lissodus cf. pattersoni из синемюриана Бельгии на основе одного неполного зуба, который также может быть отнесен к Parvodus.
Зубы Parvodus лабиолингвиально узкие и тонкие. Это отделяет их от зубов Lissodus, Vectiselachos и Hylaeobatis. Они могут быть отделены от зубов Lonchidion хорошо развитыми выступами и бугорками и более слабым губным бугорком. Зубы Парводуса внешне похожи на зубы некоторых видов Polyacrodus, но легко отделяются от зубов вида Polyacrodus polycyphus (Agassiz, 1837) небольшими размерами и изящным внешним видом. Степень разнородности у Parvodus также ниже, чем у Polyacrodus. Морфология корней зубов Polyacrodus также различна, неправильных отверстий относительно меньше, а корень лишен небольших круглых отверстий рядом с соединением коронка-корень.
Межвидовые различия между разными видами Parvodus. Передние зубы Parvodus curvidens с окклюзией изогнуты и
имеют более выраженные бугорки и выступы, но в остальном похожи на таковые у типового вида. Зубы Parvodus rugianus немного более массивные и более крупные, орнаментированы больше, чем у других видов рода. Известный стратиграфический диапазон этого рода — батский, от средней юры до валанжина, ранний мел, хотя возможно возникновение в синемурийском периоде. Вполне вероятно, что зубы этого рода в прошлом игнорировались из-за их небольшого размера и поверхностного сходство с ювенильными зубами других таксонов гибодонтов.
 |
 |
 |
 |
 |
 |
 |
Семейство Лонхидииды — † Lonchidiidae Herman, 1977, Род Вектиселахосы — † Vectiselachos Rees & Underwood, 2002 (известен из мелового периода) состоит из одного типового вида — Vectiselachos ornatus Woodward, 1889 из раннего мела Остров Уайт, Англия. [J. Rees and C. Underwood. 2002. The status of the shark genus Lissodus Brough, 1935, and the position of nominal Lissodus species within the Hybodontoidea (Selachii). Journal of Vertebrate Paleontology 22(3):471-479]
Название рода происходит от названия слоя Vectis, образования в южной Англии, где были обнаружены окаменелости типового вида, и греческого слова σελαχος (selachos) — «акула», «хрящевая рыба» .
Эти лонхидиидные акулы с выраженным зубным рядом дробящего типа; передние зубы объёмные, с хорошо обозначенными бугорками и губными выступами; слабо орнаментированы, преимущественно с бороздками и реже с гранулами; боковые зубы низкие, сильнее орнаментированы, всегда с гранулами; губные бугорки на боковых сторонах слабо развиты или отсутствует; корень заметно меньше коронки и сравнительно тонкий.
Передние зубы Vectiselachos несколько похожи на зубы Lonchidion, но всегда более луковичные и украшены гранулами, небольшими неправильные возвышения эмелоида, этот признак является исключительным для этого рода. Густая скульптура бороздок и гранул, а также форма губной выпуклости отделяет зубы Vectiselachos от зубов Lissodus и Hylaeobatis.
Зубы нового вида гибодонтовой акулы из рода Vectiselachos (Chondrichthyes: Lonchidiidae) описаны из позднего апта (раннего мела) на юге Англии. Vectiselachos gosslingi имеет очень характерные грубые бороздки, которые образуют выступы на окклюзионных поверхностях коронки.
%201f%20fossil.jpg) |
%201f%20fossil.jpg) |
Семейство Полиакродонтиды — † Polyacrodontidae Glükman, 1964, Род Палаеобатэсы — † Palaeobates Meyer, 1849 (известен из триасового и юрского периодов, от 247,2 до 155,7 миллиона лет назад): Palaeobates angustissimus Agassiz, 1838; Palaeobates polaris Stensiö, 1921; Palaeobates reticulatus Duffin, 1998; Palaeobates shastensis Bryant, 1914. По други данным род † Palaeobates Meyer, 1849 отнесён к семейству † Acrodontidae Casier, 1959.
Палеобаты имели зубцы давящего типа, которые они использовали для раздавливания добычи с твердым панцирем. Зубы имеют ортодонтическую гистологию.
В 2010 году был представлен новый хорошо сохранившийся экземпляр Palaeobates polaris из смитийского «рыбного горизонта» Шпицбергена (архипелаг Шпицберген, Арктическая Норвегия). Находка более полная, чем типовой материал Palaeobates polaris, и содержит, среди прочего, левую нижнечелюстную ветвь с соответствующими зубными рядами и губным хрящом, а также элементы подъязычной дуги и часть переднего спинного плавника. Palaeobates polaris имеет ту же гистологию зубов, что и другие виды рода Palaeobates (ортодонтические зубы с полостью пульпы), но орнамент зубов отличается. Несколько исследований показали, что установить филогенетические отношения между гибодонтиформными акулами с помощью гистологии зубов проблематично. Более того, ортодонтические зубы с полостью пульпы, вероятно, плезиоморфны Hybodontiformes. На основании новой находки Palaeobates polaris, три других признака считаются апоморфными для палеобатов в целом: (1) нижний край зубной борозды меккелевского хряща проходит ближе к вентральной части, чем к дорсальному краю нижней челюсти у Palaeobates polaris, но медиальнее этих краев нижняя челюсть в коронной группе гибодонтоидов Acrodus; (2) количество и расположение зубов Palaeobates polaris хорошо отличимы от таковых у Acrodus и Asteracanthus; и (3) шипы спинных плавников Palaeobates polaris, а также Palaeobates angustissimus имеют тенденцию быть более тонкими по сравнению с таковыми у Hybodus и Acrodus. Тем не менее, требуется больше материала, чтобы убедиться, что эти черты полезны для отделения палеобатов от других гибодонтиформ.
%201f%20fossil.jpg) |
 |
 |
 |
Семейство Полиакродонтиды — † Polyacrodontidae Glükman, 1964, Род Полиакродусы — † Polyacrodus Jaekel, 1889 (известен от пермского периода до мелового периода, от 279,5 до 70,6 миллиона лет назад): Polyacrodus claveringensis Stensiö, 1932; Polyacrodus heterodonPatterson, 1966; Polyacrodus jiangxiensis Wang et al., 2007; Polyacrodus krafti Seilacher, 1943; Polyacrodus lapalomensis Johnson, 1981; Polyacrodus minimus Agassiz, 1939; Polyacrodus parvidens Woodward, 1916; Polyacrodus polycyphus Agassiz, 1843; Polyacrodus pyramidalis Stensiö, 1921; Polyacrodus ritchiei Johnson, 1981; Polyacrodus tiandongensis Wang et al., 2001; Polyacrodus tregoi Rieppel et al., 1996; Polyacrodus wichitaensis Johnson, 1981; Polyacrodus zideki Johnson, 1981. [O. Jaekel. 1889. Die Selachier aus dem oberen Muschelkalk lothringens. Abhandlungen zur Geologischen Specialkarte von Elsass-Lothringen, Band 3 4:273-33]
Зубы умеренно большие, низкие и широкие, с преобладанием большого стержневидного корня, более высокого, чем коронка. Слабо развитый главный бугорок присутствует в средней точке коронки, с последующей серией очень слабо развитых дополнительных бугорков по длине режущей кромки. Отчётливый орнамент из мелких гребней присутствует на коронке, с гребнями, сходящимися на вершине бугорков.
Три несвязанных фрагмента зубов гибодонтовой акулы Polyacrodus sp. Были обнаружены в карьере формации Кайова (нижний мел, Альб), округ Макферсон, штат Канзас, в июне 2003 года. Зубы этой акулы представляют собой новый рекорд для малоизученной морской палеофауны позвоночных Кайова. Обломки были обнаружены вместе с зубами другого гибодонта, ската и рыбы-пикнодонта. Зубы являются первым признанным появлением «Polyacrodus» в Канзасе и еще раз подтверждают разнообразие прибрежной палеофауны в Западном внутреннем море в раннем меловом периоде.
 |
 |
 |
 |
Семейство Псевдодалатииды — † Pseudodalatiidae Reif, 1978, Род Псевдодалатиасы — † Pseudodalatias Reif, 1978 (известен из триасового периода, от 242,0 до 201,6 миллиона лет назад): Pseudodalatias barnstonensis Sykes, 1971; Pseudodalatias henarejensis Botella et al., 2009. [W. E. Reif. 1978. Tooth enameloid as a taxonomic criterion 2. Is "Dalatias" barnstonensis Sykes, 1971 (Triassic, England) a squalomorphic shark?. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 1978(1):42-58]
Псевдодалатииды, семейство хондрихтидов с неопределённым филогенетическим сходством, до сих пор были известны исключительно по зубным видам Pseudodalatias barnstonensis (Sykes, 1971), стратиграфический диапазон которых ограничен верхним триасом Европы (Англия, Италия). Псевдодалатиасы имеют характерный зубной ряд, который позволяет им удерживать и резать свою добычу, демонстрируя неоселахический дизайн, но лишённый трехслойной эмелахической микроструктуры неоселахических зубов. Открытие Pseudodalatias henarejensis nov. sp. в Ладинском регионе Испании расширяется стратиграфический диапазон и палеогеографическое распространение этого семейства. Этот новый вид также демонстрирует, что в семействе Pseudodalatiidae прогрессивно эволюционировал зубной ряд с режущим механизмом.
Все, что известно об этих животных, — это несколько окаменелых зубов, которые находят изолированными или небольшими полными сериями по одиннадцать зубов в каждом. Наборы зубов небольшие, достигают в длину максимум трех сантиметров. Полная серия состоит из центрального (симфизарного) зуба с симметричным боковым профилем и пяти пар боковых зубов с асимметричным профилем. Зубцы зазубрены и имеют от четырёх до шести зубцов с каждой стороны. Внешний вид животного совершенно загадочен; вероятно, это была крошечная хищная акула.
Pseudodalatias известен исключительно благодаря зубам, которые встречаются в некоторых европейских отложениях. Отдельные зубы вида, Pseudodalatias barnstonensis, встречаются в английском bonebeds (Костное ложе — это любой геологический пласт или месторождение, содержащее кости любого типа.) и во Франции, в то время как некоторые полные серии были найдены в Италии, особенно в провинции Бергамо. В Испании окаменелости другого вида, Pseudodalatias henarejensis, были обнаружены на более ранних уровнях (средний триас, около 240 миллионов лет назад), что показывает, как зубы этих животных специализировались на протяжении миллионов лет к функции резки.
Неясно, какова была классификация псевдодалатий; безусловно, морфология зубов очень отличается от современной. Род был отнесён к группе Euselachii в отдельное семейство (Pseudodalatidiidae). Pseudodalatias имеет характерные зубные ряды, напоминающие neoselaci, но не имеет трехуровневой пластинчатой микроструктуры, типичной для этой группы. Вероятно, что Pseudodalatias был примитивным представителем batoids или neoselacians, хотя кажется неправдоподобным, что это была аберрантная (отклоняющийся от нормального строения) акула-гибодон.
Отложения, в которых были найдены окаменелости этого животного, указывают на то, что Pseudodalatias обитали в поверхностных водах замкнутых тропических морских бассейнов, дно которых характеризовалось крайней нехваткой или недостатком кислорода. Зубы Pseudodalatias позволяли этому животному удерживать и разрезать добычу на куски.
%201f%20fossil.jpg) |
%202f%20fossil.jpg) |
%203f%20fossil.jpg) |
%201f%20fossil.jpg) |
%202f%20fossil.jpg) |
%203f%20fossil.jpg) |
Семейство Штайнбаходонтиды — † Steinbachodontidae Reif, 1980, Род Штайнбаходусы — † Steinbachodus Reif, 1980 (известен из триасового периода, от 235,0 до 221,5 миллиона лет назад) состоит из одного типового вида — Steinbachodus estheriae Reif, 1980 из Австрии (1) и Германии (6). [W. Reif. 1980. Tooth enameloid as a taxonomic criterion: 3. A new primitive shark family from the lower Keuper. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 160:61-72]
Название рода Steinbachodus происходит от названия города Штайнбах (нем. Steinbach) в Германии, в земле Гессен и греческого слова οδους (odous) — «зуб», «зубы».
Зубы Acrorhizodus напоминают зубы Steinbachodus estheriae Reif, 1980 из Нижнего Кейпера (Lower Keuper) Германии, которые намного меньше, имеют более резцовую форму и их корона явно остроконечная на неизношенных элементах. Орнамент кроны состоит из вертикальных лабиальных и язычных гребней, а не поперечных, как у Acrorhizodus. Кроме того, зубы Steinbachodus имеют менее развитый корень, лишённый крупных протоков. Таким образом, мы рассматриваем что сходство между Acrorhizodus и Steinbachodus является результатом морфологической конвергенции.
Семейство Тристихииды — † Tristychiidae Moy-Thomas, 1936, Род Тристихиусы — † Tristychius Agassiz, 1837 (известен из каменноугольного периода, от 336,0 до 326,4 миллиона лет назад) состоит из одного типового вида — Tristychius arcuatus Agassiz, 1837. [L. Agassiz. 1837. Recherches Sur Les Poissons Fossiles. Tome III (livr. 8-9). Imprimérie de Petitpierre, Neuchatel viii-72]
Окаменелости Tristychius arcuatus, включая шипы плавников, были найдены в Шотландии. Впервые вид был обнаружен швейцарским биологом Луи Агассисом в 1837 году, а позже подробно описан Джоном Диком, бывшим одноклассником Коутса, в 1978 году. Тристихиус и другие акулы девонского периода, подобные ему, найдены в конкрециях железных пород вдоль берегов залива Ферт-оф-Форт недалеко от Эдинбурга, Шотландия. Тристихиус был маленькой акулой, около 60 сантиметров в длину. У него был хорошо развитый хвостовой плавник, похожий на таковой у многих современных акул. Физически он мог напоминать современную морскую рыбу. У тристихиуса также были шипы, прикрепленные к основанию его спинных плавников, вероятно, для защиты от хищников.
У Tristychius был круглый рот, выдвинутый вперед на конце морды, как у современной акулы-няньки. Его способность контролировать это отверстие дало ему доступ к ранее неиспользованным пищевым ресурсам, таким как добыча, укрывающаяся в неглубоких норах, или трудные для отлова стаи креветок или молоди рыб.
Исследователи из Чикагского университета впервые применили инструменты, разработанные для изучения трехмерных движений и механики современных рыбьих челюстей, для анализа ископаемых рыб. Технология компьютерной томографии, способной получать изображения окаменелости, пока она все ещё заключена в каменную оболочку, показывает, что у акулы возрастом 335 миллионов лет были сложные челюсти, способные к всасывающему питанию, обычному для костистых рыб, таких как окунь, карп, а также современные акулы-няньки.
Примечательно, что эти древние челюсти акулы примерно на 50 миллионов лет старше, чем самые ранние свидетельства того, что аналогичные челюсти приспособлены для всасывающего кормления костистых рыб. Это показывает как эволюционную универсальность акул, так и то, как акулы быстро отреагировали на новые экологические возможности после одного из пяти крупных вымираний в истории Земли.
%201f.jpg) |
%202f.jpg) |
%203f.jpg) |
%204f.jpg) |
%205f.jpg) |
%208f.jpg) |
%206f%20skeleton.jpg) |
%207f%20skeleton.jpg) |
Семейство Хомалодонтиды — † Homalodontidae Mutter et al., 2008, Род Хомалодонтусы — † Homalodontus Mutter et al., 2008 (известен из триасового периода, от 251,3 до 247,2 миллиона лет назад): типовой вид — Homalodontus aplopagus Mutter et al., 2007; Homalodontus homalorhizo Mutter et al., 2007. [R. J. Mutter, A. G. Neuman, and K. Blanger. 2008. Homalodontus nom. nov., a replacement name for Wapitiodus Mutter, de Blanger and Neuman, 2007 (Homalodontidae nom. nov., ? Hybodontoidea), preoccupied by Wapitiodus Orchard, 2005. Zoological Journal of the Linnean Society 154:419-420]
Название рода Homalodontus происходит от греческих слов ομαλος (homalos) — «гладкий», «ровный», «плоский» и οδους (odous) — «зуб», «зубы».
Mutter, de Blanger & Neuman (2007) предложили название Wapitiodus (Wapitiodidae) для новой раннетриасовой гибодонтообразной акулы из группы Vega-Phroso Siltsone формации Sulphur Mountain. Они обратили внимание на то, что это название связано с родовым названием конодонта Wapitiodus Orchard (2005: 98–99). Согласно статье 52 4-го издания Международного кодекса зоологической номенклатуры (МКЗН) родовое название акулы является младшим омонимом родового названия конодонта. Это название необходимо изменить, и предложили новое название Homalodontus для рода акул. Чтобы предотвратить возможную путаницу в будущем, и следуя Принципу координации, применяемому к названиям групп семейств (ICZN изд. 4, статья 39), также предложили новое название семейства Homalodontidae для Wapitiodidae Mutter et al ., 2007.
%201f%20fossil.jpg) |
 |
Семейство Птиходонтиды — † Ptychodontidae Jaekel, 1898, Род Гетероптиходусы — † Heteroptychodus Yabe & Obata, 1930 (известен из мелового периода, от 140.2 до 122,46 миллиона лет назад): Heteroptychodus kokutensis Cuny et al., 2010, типовой вид — Heteroptychodus steinmanni Yabe & Obata, 1930. [R. J. Mutter, A. G. Neuman, and K. Blanger. 2008. Homalodontus nom. nov., a replacement name for Wapitiodus Mutter, de Blanger and Neuman, 2007 (Homalodontidae nom. nov., ? Hybodontoidea), preoccupied by Wapitiodus Orchard, 2005. Zoological Journal of the Linnean Society 154:419-420]
Ptychodontidae имели очень необычные для хрящевых рыб зубы: у них нет лезвия, а их рабочая поверхность широкая и часто почти плоская. Их эмаль покрыта рёбрами (обычно параллельными, но иногда радиально расходящимися из центра). Эти зубы покрывали челюсти, плотно состыковываясь друг с другом. Таким образом, это типичный дробящий зубной аппарат, которым рыбы дробили раковины и панцири своей добычи. Подобное устройство зубного аппарата — большая редкость для хрящевых рыб (среди современных родов дробящие зубы имеют обыкновенные куньи, вислоносые и рогатые акулы).
Обзор фауны пресноводных акул из формация Пху Крадунг (Phu Kradung Formation) на северо-востоке Таиланда позволяет признать новый вид рода Acrodus. Остальная часть фауны акул включает зубы Hybodus, дермальные зубчики гибодонтид, Jiaodontus, Lonchidion, Heteroptychodus. Присутствие зубчиков Heteroptychodus kokutensis и шипы спинного плавника предполагает раннемеловой возраст вершины этой формации. Однако возраст Пху Крадунг все ещё неопределён, с противоречивыми сигналами из палинологии, термохронологии детритового циркона и палеонтологии позвоночных. В любом случае оказывается, что это старейшее появление рода Heteroptychodus, и предполагает тайское происхождение этого рода, который может быть заменил род Acrodus в тайских пресноводных палеоэкосистемах.
Самый большой зуб Heteroptychodus kokutensis размером 4,5 мм мезио-дистально (мезиально или дистально
отсутствует), 1,5 мм в лабио-лингвальной области, высота 2 мм, включая корень. Коронки удлинённые в мезио-дистальном направлении, плоские или слегка выпуклые, без бугорка, орнаментированные набором гребней, проходящих мезио-дистально и параллельно друг другу. Есть два-четыре гребня. Только самый губной достигает
мезиальные и дистальные конечности коронки. Лингвально, они становятся короче. Маленькие короткие гребни берут начало в обоих языковых и лабиальной стороне от самого губного гребня и перпендикулярны к последнему. Языковая часть апикальной грани коронки орнаментирована грубыми неровными гребнями, которые иногда образуют узлы на верхняя часть языковой стороны. Губная часть выпуклая мезиально или дистально с короткими выступами в базальной части, которые, однако, не достигают основания коронки.
%201f%20fossil.jpg) |
 |
 |
 |
Семейство Птиходонтиды — † Ptychodontidae Jaekel, 1898, Род Птиходусы — † Ptychodus Agassiz, 1837 (известен из мелового периода, по некоторым данным, есть находки из нижнего палеогена, от 94,3 миллиона лет назад): Ptychodus altior Agassiz, 1839; Ptychodus anonymus Williston, 1900; Ptychodus arcuatus Agassiz, 1837; Ptychodus articulatus Agassiz, 1837; Ptychodus belluccii Bonarelli, 1899; Ptychodus concentricus Agassiz, 1839; Ptychodus decurrens Agassiz, 1839; Ptychodus depressus Dixon, 1850; Ptychodus elevatus Leriche, 1929; Ptychodus gibberulus Leriche, 1929; Ptychodus latissimus Agassiz, 1843; Ptychodus mahakalensis Chiplonkar & Ghare, 1977; типовой вид — Ptychodus mammillaris Agassiz, 1839; Ptychodus marginalis Agassiz, 1839; Ptychodus martini Williston, 1900; Ptychodus mortoni Agassiz, 1843; Ptychodus multistriatus Woodward, 1889; Ptychodus occidentalis Leidy, 1868; Ptychodus oweni Dixon, 1850; Ptychodus papillosus Cope, 1875; Ptychodus parvulus Whiteaves, 1889; Ptychodus paucisulcatus Dixon, 1850; Ptychodus polygyrus Agassiz, 1839; Ptychodus rugosus Dixon, 1850; Ptychodus spectabilis Agassiz, 1837; Ptychodus whipplei Marcou, 1858. [L. Agassiz. 1837. Recherches Sur Les Poissons Fossiles. Tome III (livr. 8-9). Imprimerie de Petitpierre, Neuchatel viii-72]
Ptychodus — род акул-дурофагов (сокрушающих панцири) из позднего мела. Их зубы собирали на всех континентах, включая Австралию, но чаще всего встречаются в отложениях, отложенных в Западном внутреннем море Северной Америки. Они вымерли во время сантона, около 85 миллионов лет назад. Птиходусы были морскими донными рыбами, внешне похожими на представителей отряда воббегонгообразных. В основном обитали в морях Северного полушария, где были самыми крупными рыбами. Наиболее крупные виды — Ptychodus mortoni и Ptychodus rugosus. Первый достигал в длину 10 метров, второй — 14,5 метров, что делало их самыми крупными рыбами мелового периода. Так как Ptychodus принадлежат к хрящевым рыбам, в ископаемом состоянии от них обычно сохраняются только зубы. Это типичные дробящие зубы: тупые, плоские и широкие, их жевательная поверхность покрыта рёбрами. Такие зубы характерны для животных, питающихся твёрдой пищей (моллюсками и другими животными, защищёнными раковиной или панцирем).
Ptychodus mortoni жили в коньякском и сантонском веках 85,8—66,0 млн лет назад верхнемеловой эпохи. Впервые зубы этого вида нашёл американский конхиолог и палеонтолог Тимоти Аббот Конрад в 1833 году в штате Алабама. Много ископаемых остатков было найдено и в Канзасе. Здесь проходил Западный центральный морской путь — море, которое в середине и конце мелового периода покрывало большие площади в центральной части Северной Америки. Зубы Ptychodus mortoni отличаются от зубов почти всех остальных видов птиходусов тем, что на них рёбра расходятся радиально от вершины. По общей форме зубы этого вида больше всего похожи на зубы Ptychodus whipplei, но имеют менее выступающую вершину.
Как выяснилось, акула, названная исследователями Ptychodus mortoni, использовала свою потрясающую челюсть не только для укусов своих жертв, но и для разгрома жилищ морских жителей — раковин древних моллюсков и ракообразных, кораллов и даже подземных нор некоторых видов рыб. Сначала палеонтологи решили, что птиходус питался гигантским планктоном, который также обитал в древних морях в юрском и меловом периоде, но позже установили, что главной пищей для акулы служили рыбы, моллюски и древние раки. По словам ученых, колоссальная рыбина, предпочитавшая, как и современные предки, живых обитателей морей, просто растирала в порошок убежища рачков и даже ветви кораллов.
Еще одним шоком стало то, что потрясающий своими размерами и мощью птиходус был, в общем-то, никудышным пловцом. Доисторическая рыбина со сверхсильными челюстями перемещалась преимущественно в нижних слоях океана, шаркая брюхом об дно. Подниматься выше акуле не было необходимости — её основные жертвы жили в кораллах, подземных норах, и поэтому охотиться, выслеживая добычу, как это делают современные акулы, ей было попросту не нужно. Высокой скоростью первобытная акула тоже не обладала. А зачем, если из норы или раковины уже никуда не сбежать? Потрясающие челюсти и огромная сила позволили гигантской рыбине стать настоящей царицей придонных вод и гарантировали полное отсутствие врагов. Даже древние морские ящеры, размеры которых могли превышать масштабы Ptychodus mortoni не могли противостоять хищной рыбине из-за колоссальных челюстей. Однако и неторопливость, и плохие способности к маневренности и плаванию — скверные свойства для естественного отбора беспощадной природы. В ходе своего развития эволюция миллионы лет отбирала лучших из лучших, и птиходус, к сожалению, оказался за бортом. 65 миллионов лет назад неторопливые акулы с уникальным "зубным плато" вместо обычных зубов вымерли вместе с другими исполинами древнего мира — динозаврами.
Новый род и вид Paraptychodus washitaensis n. gen. et n. sp. описан на основе 13 зубов из среднеальбской формации Дак Крик группы Вашита на севере центрального Техаса, США. Этот материал имеет важное значение, так как демонстрирует промежуточную форму зубов между Lonchidiidae и Ptychodus в отношении окклюзионного орнамента коронки и морфологии корня зуба; новый таксон представляет собой самого раннего представителя семейства Ptychodontidae в Северной Америке. Анализ дентальных признаков и стратиграфических проявлений внутри семейства позволяет предположить, что Paraptychodus washitaensis является базальным таксоном и самым недавним предком Ptychodus. Морфологические тенденции производных видов Ptychodus демонстрируют постоянную специализацию в морфологии коронок зубов. Диагноз Paraptychodus washitaensis из верхнего альба Техаса позволяет скорректировать стратиграфическое распространение рода Ptychodus от нижнего сеномана до нижнего кампана.
%201f.jpg) |
%202f.jpg) |
%203f.jpg) |
%204f.jpg) |
%205f%20fossil.jpg) |
%206f%20fossil.jpg) |
%207f%20fossil.jpg) |
%201f%20fossil.jpg) |
%202f%20fossil.jpg) |
 |
 |
 |
 |
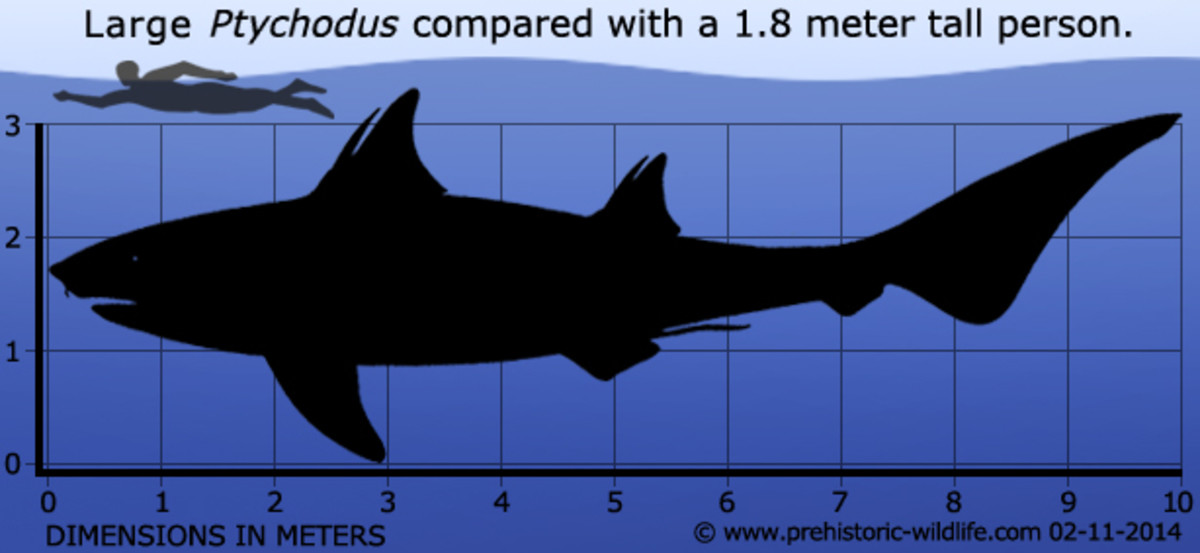 |
 |
 |