Anarhichas denticulatus Krøyer, 1845
Синяя зубатка, синюха, вдовица, баренцовоморская синяя зубатка (Rus),
Northern wolffish, Blue sea-cat, Jelly-cat
(Eng)
Синонимы:
Anarhichas denticulatus Krøyer, 1845
Anarhichas latifrons Steenstrup & Hallgrimsson, 1876 Синяя зубатка (Rus),
Anarrhichas fortidens Gill, 1911
Lycichthys denticulatus (Krøyer, 1845)
Lycichthys fortidens Gill, 1911
Lycichthys latifrons Gill, 1911
Lycichthys parvodens Lühmann, 1954
Lycichthys paucidens Gill, 1905
Рыбы открытого океана. Н.В.Парин 1998 г.
Наиболее глубоководный вид - синяя зубатка (Anarhichas denticulatus), достигающая 1,4 м в длину, отмечена даже на глубине более 900 м. Все зубатки откладывают очень крупную донную икру, но их личинки поднимаются в толщу воды, где проходят весь мальковый период жизни (его продолжительность у разных видов сильно разнится).
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Синяя зубатка (Anarhichas latifrons)
отличается самым слабым развитием бугорковидных зубов; сошниковый ряд
зубов у нее много короче нёбных рядов,
тогда как у всех других видов зубаток, после
перехода их к жизни у дна, он длиннее
нёбных.
Имеются точные данные о поимках в
трал синих зубаток длиной до 138 см при
весе 32 кг, но, по некоторым сведениям,
на яруса вылавливались и двухметровые
синие зубатки.
Окраска синей зубатки
однотонно-темная, пятна неяркие и нечеткие,
группировка их в полосы обычно совсем
незаметна.
Ловится синяя зубатка в тех же
районах, что и пятнистая, но у острова Ян-Майен встречен пока что только этот вид.
Мертвый экземпляр синей зубатки был
найден на берегу острова Принс-Патрик,
далеко к западу от ныне известных
районов обитания атлантических зубаток.
Предпочитает наибольшие глубины (до
936 м) и совершает большие
передвижения, чем все другие виды зубаток.
В созревающих самках синей зубатки
находят 23—29 тыс. икринок (при длине
рыб 112—124 см) диаметром 6—7 мм; эти
икринки выметываются весной, летом или
осенью. Кладки синей зубатки пока еще
никто не находил. До сего времени
описаны всего 2 малька длиной 25 и 26 мм,
пойманных в мае над материковым свалом
между Лофотенскими островами и
островом Медвежьим. Самки со зрелей икрой
обычно ловятся поблизости от свала, а
восточнее, в Баренцевом море,
встречаются только незрелые; отсюда происходит
еще одно, поморское, название синей
зубатки — вдовица.
Переход молоди к жизни у дна очень
сильно запаздывает: синяя зубатка
появляется в траловых уловах обычно лишь
при длине 60—70 см.
Питается синяя зубатка в основном
гребневиками, медузами и рыбой, а
иглокожих, ракообразных и особенно моллюсков потребляет гораздо меньше, чем
другие виды зубаток. Зубы у нее иногда
совсем не стираются, но тем не менее
меняются ежегодно, как и у других
зубаток.
Мясо синей зубатки жидкое, водянистое,
а поэтому синюю зубатку до недавнего
времени не заготовляли, выбрасывали за
борт или резали на куски и наживляли
ими яруса, особенно при ловле палтуса.
В мясе много витамина А, возможно, в
связи с быстрым ростом; вообще, чем больше
глубины, на которых предпочитают
держаться атлантические зубатки, тем
быстрее они растут: например, в возрасте 7 лет
беломорская зубатка достигает в среднем
37 см, баренцевоморская полосатая 54 см,
пятнистая 63 см и синяя 92 см.
Фауна СССР. Рыбы. Том V, вып. 5. Семейство Зубаток (Anarhichadidae). В.В.Барсуков 1959 г.
2. Anarhichas latifrons Steenstrup et Hallgrimsson — Синяя зубатка, синюха, вдовица.
(Latus — широкий, frons — лоб.)
Blaasstenbid, hakjorring stenbid (норвежцы); blа stenbit (шведы); bredpandet havkat (датчане); blue sea-cat, jelly-cat (англичане); Wasserkatze, Blauer Katfisch (немцы); blagoma (исландцы); leveäotsainen merikissa (финны).
Anarrhichas latifrons Steenstrup, Förh. Scand. Naturf., 3 Möte (1842), 1843 : 647 (Исландия; nomen nudum). — Steenstrup et Hallgrimsson in: Steenstrup, Vidensk. Meddel. Naturh. Foren., (1876), 1877 : 177—201, tab. III, fig. 3 (Гренландия, Исландия; морфология, питание). — Collett, Christ. Vidensk.- Selsk. Forhand., (1879), 1, 1880 : 46, pl. (fig.) II (Финмаркен, сравнение с другими видами) — Bean, Proc. U. S. Nat. Mus., II (1879), 1880 : 217 (синонимия, измерения). — Collett, Nyt. Mag. Natur., 29, 2, 1885 : 67—68 (измерения, питание). — Lilljeborg, Sverig. Norg. Fisk., 1, 1891 : 546—547 (измерения, описание скелета). — Smitt, Scand. Fish., I, 1893 : 237, pl. XIII, fig. 2 (цветн. рис.; питание). — Good a. Bean, Ocean. Ichth., 1895 : 301, fig. 271 (измерения, число позвонков и лучей в плавниках; 42°50'20" N, 50°51' W, глуб. 471 морск. саж.). — Книпович, Определитель, 1926 : 94, фиг. 60 (Баренцево море, до 128 см).
Anarrhichas denticulatus Krøyer, Overs. Dansk. Vidensk. Selsk. Forh., 1844 : 139 (Гренландия). — Gaimard, Voy. Scand. Lapp. Spitzb. Feroe, Paris, 1845 (?) : pl. 12 (цветн. рис.). — Günther, Cat. fish., III, 1861 : 211 (Гренландия, D 80). — Collett, Rep. Norw. mar. fish, invest., II, 3, 1905 : 114 (синонимия; Фарерские острова, хребет Уайвиля Томпсона).
Anarhichas latifrons Cornish, Rept. Dept. Mar. Fisher. Canada (1902—1905), 39, 1907 : 87 (Нов. Шотландия, на глубине 50 морск. сажен). — Вrеdеr, Fieldbook mar. fish. Labr. Texas, 1929 : 275, 276 (определительная таблица; Нов. Шотландия). — Saemundsson, Zool. Iceland, IV, 72, 1949 : 31—34 (Исландия, синонимия; на глубине до 850 м; длиной менее 70 см не встречается). — Есипов в изд.: Пром. рыбы СССР, 1949 : 616, атлас, табл. 189 (сводка, цветн. рис.; молодь 11—13.5 см у о. Медвежьего). — Барсуков, Докл. АН СССР, XCV, 4, 1954 : 897—899 (смена зубов).
Anarhichas denticulatus Jensen, Spolia Zool. Mus. Haun., IX, 1948 : 121—123 (Гренландия, синонимия; на глубине до 520—737 морск. сажен; питание) — Walters, Amer. Mus. Novitates, No. 1643, 1953 : 3—4 (о. Принца Патрика). — Walters, Bull. Amer. Nat. Hist., 106, 5, 1955 : 306 (боковой линии нет?).
Lycichthys latifrons Gill, Proc. U. S. Nat. Mus., XXXIX, 1911 : 167 (Исландия и западное побережье Гренландии, по Стенструпу). — Jordan, Evermann a. Clark, Check list fish. N. a. M. Amer., 1930 : 472 (синонимия).
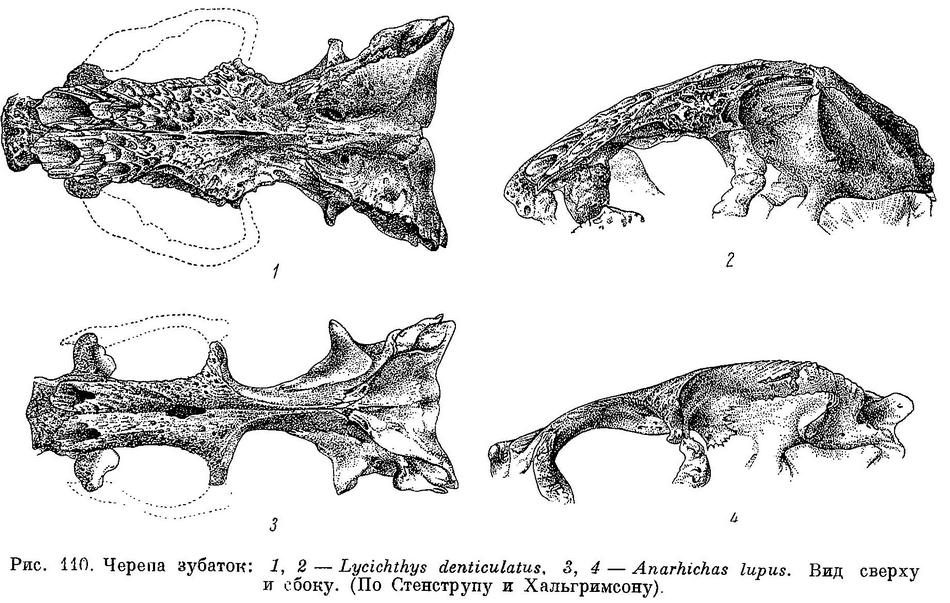
Lycichthys denticulatus Gill, Proc. U. S. Nat. Mus., XXXIX, 1911 : 167 (западное побережье Гренландии, по Кройеру). — Андрияшев, Рыбы сев. морей СССР, 1954 : 220—222, фиг. 110, 1, 2, 111, 1, 112.
Lycichthys fortidens Gill, Proc. U. S. Nat. Mus., XXXIX, 1911 : 166—167, fig. 8 (побережье штата Мэн и Нов. Шотландии).
Lycichthys paucidens Gill, Bull. Biol. Soc. Wash., 18, 1905 : 251 (побережье штата Мэн и Нов. Шотландии). — Gill. Proc. U. S. Nat. Mus., XXXIX, 1911 : 167, fig. 9—10, pl. 18 (расположение зубов).
Lycichthys latifrons + Lycichthys denticulatus + Lycichthys fortidens + Lycichthys paucidens + Lycichthys parvodens Lühmann, Zeitschr. Zellforsch. mikrosk. Anat., 40, 5, 1954 : 470—509 (смена зубов, гистология).
Распространение, биология. Giinther, Rep. deep, sea fish., XXII, 1887 : 71 (42°27' N, 64°20' W). — Good, Fish. a. fish, industr. U. S., I, 1884 : 248 (Ново-Шотландские банки; на глубинах 200—400 морск. сажен). — Книпович, Эксп. научно-пром. исслед. Мурмана, I, 1902 : 55 и сл., фиг. 7 (многочисленные местонахождения у берегов Мурмана; фотогр.). — Брейтфус, там же, II, 1903 (местонахождения в Баренцевом море). — Книпович, там же, II, 1, 1904 (местонахождения в Баренцевом море). — Gill, Trans. Nat. Hist. Soc. North., 1, 1904 (1907) : 499— 500 (у берегов Великобритании; цит. по: Bigelow. a. Schroeder, 1935 : 17). — Gill, Rep. Dove Mar. labor., Cullercoats, 1906 : 47 (цит. по: Bigelow a. Schroeder, 1935 : 17). — Брейтфус, Эксп. научно-пром. исслед. Мурмана, II, 1906; II, 1915 (местонахождения в Баренцевом море). — Солдатов, в сб. «Естеств. произв. силы России», VI, 3 (2), 1920 : 18 (использование). — Thilemann, Wiss. Meeresunters. (Helgoland), XIII, 2, 1921 : 206 (Мурман; питание). — Wоllеbаеk, Norg. fisk., 1924 : 212, fig. 236 (северное побережье Норвегии, на глубинах 300—600 м). — Jenkins, Fish. Brit. isles, 1925 : 109 (у берегов Великобритании). — Jensen, Rap. Proc.-Verb., XXXIX, 1926 : 99 (Зап. Гренландия). — Saemundsson, Synops. fish. Iceland, 1927 : 18 (вокруг Исландии). — Броцкая, Докл. 1-й сесс. Гос. океаногр. инст., 4, 1931 : 50—51, рис. 6, 28, 1 (питание). — Lundbeck, Wissenschaftl. Meeresunters., Helgol., XVIII, 3, 1932 : 13—16, 18 (Баренцево море). — Смарагдова, Тр. Гос. океаногр. инст., III, 2, 1933 (Баренцево море, уловы). — Bigelow a. Schroeder, Proc. Bost. Soc. Nat. Hist., XLI, 2, 1935 : 15, pl. 3 (Ново-Шотландские банки). — Merriman, Copeia, 3, 1935 : 143 (Ново-Шотландские банки; фотогр.). — Ehrenbaum, Handb. Seefischer. N. Europas, 1936 : 141 (сводка). — Lundbeсk, Ber. Deutsch. wiss. Komm., N. F., VIII, 2, 1936 : 100, tab. 4 (низкое качество мяса). — Есипов, Пром. рыбы Баренц. моря, 1937 : 51, фиг.17 (промысел). — Nybelin, in: Andersson, Fiskar. Fiske Norden, 1, 1942 : 104 (м. Скаген, о. Мосешер). — Mаслов, Тр. Полярн. научно-исслед. инст. морск. рыбн. хоз. и океаногр., VIII, 1944 : 154 (Баренцево море; темп роста, плодовитость, промысел). — Травин, в сб. «Пром. рыбы Баренц. и Бел. морей», 1952 : 170 (питание). — Bigelow a. Schroeder, Fish. Gulf Maine, 1953 : 502 (конкретных сведений о находках в заливе Мэн нет.). — Rae a. Wilson, Scott. Nat., 65, 3, 1953 : 150 (банка Викинг, в июне; Фарерские о-ва). — Полянский, Тр. Зоолог. инст., XIX, 1955 : 29, 50, 59, 61, 64, 70, 78, 83, 90, 94, 97, 102, 109, 116 (паразиты). — Барсуков, Вопр. ихт., 8, 1957 : 42—44, рис. 9, 10, 11, 3, 12, 3, 13, 3 (распределение в Баренцевом море). — Kotthaus u. Krefft, Ber. Deutsch. wiss. Komm. Meeresforsch., XIV, 3, 1957 : 177—178 (места поимки у побережья Гренландии; экземпляр длиной 6 см из желудка трески, без описания).
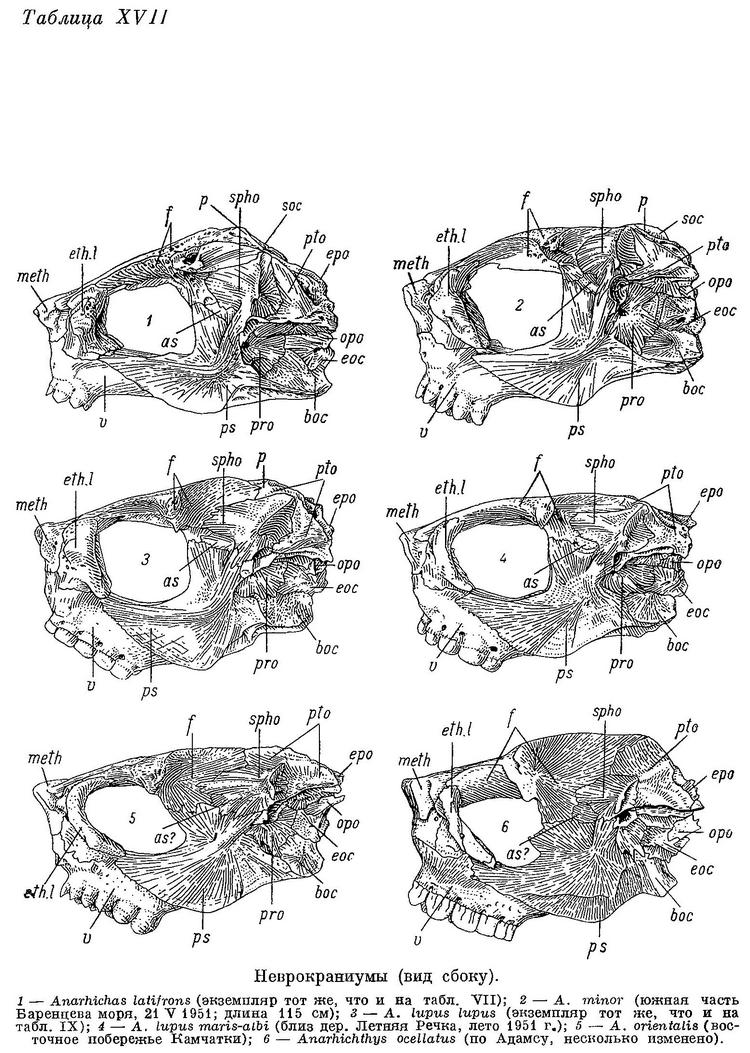
D 76—81, А (45) 46—50, Р 19—21 (22), С 19—22, позвонков 78—82 (туловищных 27—29, хвостовых 51—54), лучей жаберной перепонки 7, окологлазничных косточек (с praeorbitale) 6—8. Наибольшая высота тела у взрослых 3.6—4.2 раза в его длине. Антеанальное расстояние 49.1—55.2%, антедорсальное 14.9—18.2%, длина головы от конца рыла до заднего угла operculum 16.8—18.0% длины тела. Расстояние от начала спинного плавника до верхнего края жаберной щели более 1/2 длины головы. Ширина основания грудного плавника составляет менее 40% высоты тела у анального плавника. Основания спинного и анального плавников утолщены, граница между ними и телом слабо заметна. Хвостовой плавник усеченный. Нижняя поверхность верхней губы перед клыками покрыта сосочками, ее толщина здесь у взрослых значительно превосходит толщину нижней губы.
Зубы мельче и более заострены, чем у других зубаток. Сошниковый ряд зубов очень короткий, его длина составляет лишь 0.5—0.7 длины нёбного ряда зубов. Нёбные ряды зубов простираются назад значительно дальше сошникового ряда. Боковые зубы внешнего ряда на нижнечелюстных костях и зубы на нёбных у взрослых конические, у молоди длиной 60—70 см — с загнутыми вершинами (клыкообразные). Наибольшие бугорковидные зубы на нижней челюсти (у молоди они конические или тупоконические) расположены на площадке клыков или на пережиме. Внешний ряд зубов на нижней челюсти не образует перерыва в области пережима нижнечелюстной кости. Жевательные мышцы развиты слабо, составляют всего лишь 5.7—6.5% от веса головы с плечевым поясом и около 1% от веса тела. Сухожилие, идущее от переднего края порции к передней части maxillare, обычно имеется. Frontale позади заглазничных отростков не сужена в киль, ширина ее здесь несколько превосходит ширину между орбитами, реже равна последней. Sphenoticum не опущено: его верхний край прикрывает сбоку сочленение frontale с pteroticum. Верхний край гребней на sphenoticum расположен позади вертикали переднего края parietalia. Крыша черепа куполообразно выпуклая (табл. VII, XVII). Задний угол глазницы (между крыльями и телом парасфеноида) находится примерно на вертикали заглазничных отростков frontale. Бугорки на opisthoticum не сдвинуты вверх. Ширина хрящевой пластинки, разделяющей scapula и coracoideum, превышает ее длину не более чем в 1.5 раза. Все кости непрочные, легко ломаются, имеют пористую структуру. Обонятельные нервы по толщине примерно равны зрительным. Селезенка округлая. В мышцах содержится более 90% воды.
Тело серовато-бурое. Интенсивность бурых тонов наиболее изменчива: от легкого бурого оттенка до густого шоколадного цвета. В последнем случае маскируется светло-фиолетовый оттенок на теле и неясные темные пятна, обычно заметные на спине и боках.
Описание дано по 16 экз. из Баренцева моря, из них позвонки и лучи в плавниках подсчитаны у 6—10 экз., а описание черепа и жевательных мышц дано по 3—4 экз.
Длина в Баренцевом море до 138 см, вес до 32 кг, в уловах обычна от 70 до 120 см, преобладающие размеры 90—110 см при весе 10—20 кг. Средняя длина в 1946—1951 гг.: в южной части Баренцева моря 96 см, в районе Центральной возвышенности 104 см, у о. Медвежьего 101 см, у западного побережья Шпицбергена 78 см. Максимальная длина у берегов Исландии и Гренландии 120 см (Saemundsson, 1949 : 34; Iensen, 1948 : 122). Есть указания на длину до 2 м (Nybelin, in Andersson, 1942 : 104).
Гилл (Gill, 1911 : 162—163) выделяет Anarhichas latifrons в особый род Lycichthys на основании ряда признаков, сведенных им в таблицу. Привожу свои замечания относительно этих признаков, придерживаясь таблицы Гилла.




Из таблицы видно, что довольно многие из приведенных Гиллом различий (см. число лучей жаберной перепонки, ось черепа, постфронтальная область, sphenoticum и parietale, suprascapula) в действительности не существуют или имеют иной характер, нежели приписываемый им. Следует отметить, что и в характеристике семейства, данной Гиллом, обнаруживаются неточности: Гилл пишет, что у зубаток нет миодома и что межчелюстные кости соединены швом с этмоидом (сустав praemaxillare-mesethmoideum был правильно описан еще Андре — Andre, 1784 : 276).
Различия в заостренности, разобщенности и числе зубов в значительной мере обусловлены сезонной, индивидуальной и возрастной изменчивостью, не учитываемой Гиллом. Возрастные изменения этих признаков не прекращаются у зубаток и поеле созревания, хотя темп их, конечно, снижается. Тем не менее эти различия, как и все остальные, между Anarhichas latifrons и Anarhichas lupus по большей части выражены резко, но в каждом из них между указанными крайними видами становится переходный — Anarhichas minor. Сюда можно добавить целый ряд других признаков, не указанных Гиллом, а также упомянутую им по Стенструпу и Коллетту форму лучей спинного плавника. По большинству этих признаков Anarhichas minor стоит ближе к Anarhichas lupus, нежели к Anarhichas latifrons, но другие, наоборот, сближают Anarhichas latifrons и Anarhichas minor, обособляя Anarhichas lupus. Есть и такие признаки, которые сближают Anarhichas latifrons с Anarhichas lupus и отделяют от них Anarhichas minor. Так, у Anarhichas minor, в отличие от других зубаток, сильно развита выемка в задней части спинного плавника, край praeoperculum в области нижнего отверстия для канала боковой линии не сужен, а расширен, вертикальный отросток межчелюстной кости более высок, верхнечелюстная кость дальше заходит за короноидной отросток (Последнее наблюдается также у Anarhichas orientalis.) и т. д.
Последовательность в построении системы семейства требует скорее выделения в особый род Anarhichas orientalis, нежели Anarhichas latifrons. В самом деле, основным признаком для деления зубаток на роды служит резкое различие между Anarhichthys и Anarhichas в числе позвонков, а также лучей в спинном и анальном плавниках. По всем этим признакам Anarhichas orientalis без переходов отличается от атлантических видов, связанных переходами.
Впрочем, выделение любых систематических групп в принципе следовало бы производить, лишь выяснив родственные связи между изучаемыми организмами и различия в древности их происхождения — степень важности тех или иных признаков можно достойно оценить только с этой точки зрения. Поскольку схема эволюции семейства еще недостаточно обоснована, лучше всего воздержаться от введения в систему излишних подразделений, тем более, что практически нужды в этом нет.
Вследствие недоучета сезонной, индивидуальной и возрастной изменчивости в форме, расположении и числе зубов Гилл описал по единичным экземплярам одного и того же вида — Anarhichas latifrons — целых 4 вида.
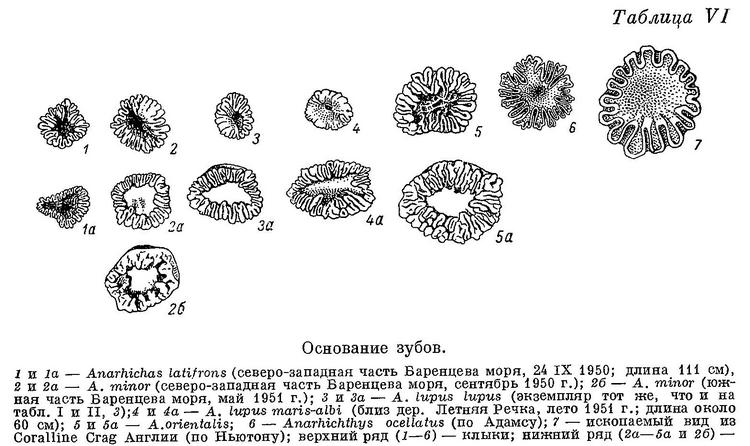
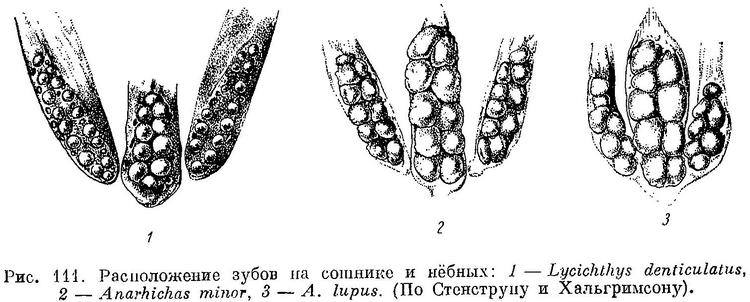
Недостаточный учет возрастной изменчивости явно выступает на примере Lycichthys paucidens: в определительной таблице, Lycichthys paucidens приписывается лишь один ряд зубов на нижней челюсти, тогда как на рис. 10, где изображены зубы этого «вида», ясно виден второй ряд зубов. Этот ряд короток, простирается назад лишь немного далее пережима, что характерно для молоди и других видов зубаток. Едва ли может возникнуть сомнение в том, что Lycichthys paucidens Гилла является всего лишь молодым экземпляром Anarhichas latifrons. Изменчивость формы, числа и расположения зубов, которую мне пришлось наблюдать у разных особей Anarhichas latifrons из Баренцева моря, полностью вмещает в себя признаки всех трех остальных описанных Гиллом видов рода Lycichthys. Вопреки необоснованному предположению Гилла, указанная изменчивость внутри вида Anarhichas latifrons оказалась ничуть не меньшей, чем аналогичная изменчивость внутри видов Anarhichas minor и Anarhichas lupus.
Вид Anarhichas latifrons Steenstrup et Hallgrimsson по правилам приоритета должен называться Anarhichas denticulatus Krøyer. Однако, учитывая то обстоятельство, что название Anarhichas latifrons гораздо более принято, особенно в отечественной литературе, его следует сохранить. К тому же Стенструп, по-видимому, имел достаточные основания для предварительного сообщения о данном виде, опубликованном на 2 года ранее описания Кройера, так как в дальнейшем описал его гораздо полнее, чем Кройер, дал лучшие рисунки зубов, а также отличные рисунки черепа.
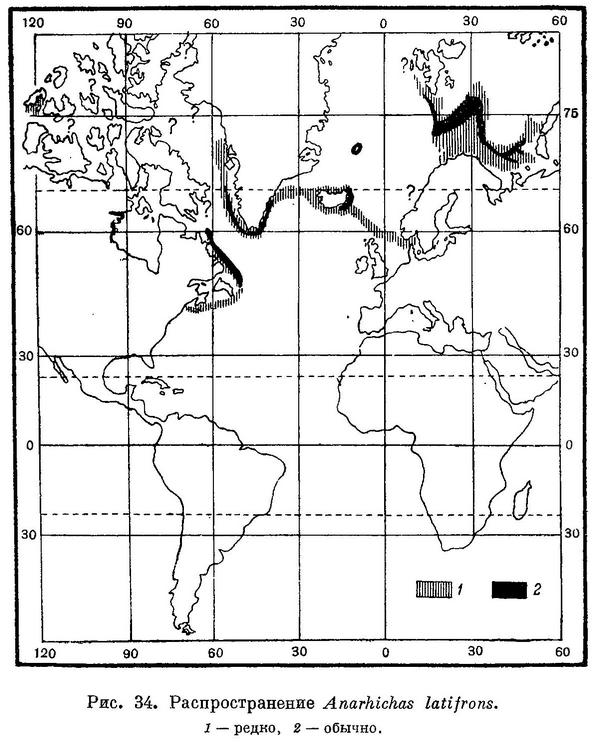
Распространение. Северная часть Атлантического океана (рис. 34). В Баренцовом море на север до возвышенности Персея, на северо-восток и восток до губы Крестовой на Новой Земле и южной части Новоземельского мелководья. В Кольском и Мотовском заливах синяя зубатка не встречается, но отмечена в Варангер-фиорде. В Белом море отсутствует. Вдоль материкового свала встречается по крайней мере от о. Принца Карла на севере до Лофотенских островов на юге, однако известна и южнее — на глубинах у Фарерских и Шетландских островов и на подводном хребте между Фарерскими островами и Исландией. Отмечены случаи поимки на банке Викинг (Rae a. Wilson, 1953 : 150), а также в проливе Скагеррак — 6 VI 1934 на глубине 270 м у мыса Скаген и 16 V 1941 на глубине 120 м у о. Мосешер (Nybelin, in Andersson, 1942 : 104). Однажды поймана у берегов Великобритании (Gill, 1904 : 499—500, цит. по: Bigelow a. Schroeder, 1935 : 17). Вокруг Исландии обычна, чаще встречается на восточном побережье. На западном побережье Гренландии отмечена к северу пока что до фиорда Упернавик, ловилась и далеко от берегов в Дэвисовом проливе. Экземпляр длиной 116 см был найден на о. Принца-Патрика (северное побережье Канады), на берегу: очевидно, был выкинут волной после гибели (Walters, 1953 : 3). На восточном побережье Гренландии найдена пока что до Ангмагссалик. Датский пролив (Kotthaus u. Krelft, 1957 : 177). Известна в районе Ново-Шотландских банок и южнее, почти до мыса Код. В залив Мэн, возможно, не заходит: точных сведений о находках ее там не имеется.
Биология. В Баренцевом море встречается на глубинах от 65 до 495 м. У островов Анде, Вестеролена и Фарерских известна на глубинах 300—600 м, у побережья Исландии — от глубин менее 100 м и до 850 м, у берегов Северной Америки от 90 до 850 м, у побережья Гренландии — от 70 м до 936—1326 м (Jensen, 1948 : 122; Kotthaus u. Krefft, 1957 : 177). У берегов Северной Америки чаще ловится на внешних склонах банок, на глубине 270—720 м (Good, 1884 : 248; Bigelow a. Schroeder, 1935 : 17), у берегов Исландии — на глубинах 150—200 м, в Баренцевом море — на 150—400 м, причем на наибольших глубинах держится здесь зимой (Барсуков, 1957 : 42). В сентябре—октябре 1955 г. у побережья Гренландии наибольшие уловы тралом (10—15 экз. за полчаса траления) получены на глубинах 90, 110—160, 120 и 370—425 м. Наименьший экземпляр длиной 52 см пойман на глубине 370—425 м (Kotthaus u. Krefft, 1957 : 177). Предпочитает илистые грунты и высокие солености. ЭНПИМ не встретила ее при солености менее 34.51‰. Встречается при температурах от —1.4° (Книпович, 1902) до 5—6°. В Баренцевом море предпочитает температуры от 2 до 4°. У побережья Гренландии в июне—октябре 1955 г. ловилась при температуре от 1.3 до 5.9° (Kotthaus u. Krefft, 1957 : 177).
Миграции, места и сроки нереста не изучены. Судя по распространению Anarhichas latifrons в Баренцевом море, миграции ее, по-видимому, сходны с миграциями пятнистой зубатки (см. Anarhichas minor), но могут иметь большую протяженность. Заходы Anarhichas latifrons в районы, довольно удаленные от основных мест обитания (мыс Скаген и о. Мосешер, о. Принца-Патрика), также могут свидетельствовать о наибольшей подвижности этого вида по сравнению с остальными зубатками.
Весьма немногочисленные наблюдения над состоянием половых продуктов у баренцевоморских особей дают повод к двум различным предположениям: 1) нерест очень сильно растянут и может происходить у разных особей по крайней мере с апреля по октябрь; 2) нерест происходит примерно в августе, но оставшиеся после него единичные крупные икринки могут сохраняться в яичниках очень долго. Трудно сказать, какое из этих предположений больше отвечает действительности. 7 VIII 1939 добыта самка IV стадии (экспонат из музея ПИНРО). В конце октября 1951 г. в районе Медвежинской банки все исследованные самки (несколько экземпляров) имели VI—II стадии, некоторые с крупными икринками, оставшимися после нереста (по сообщению М. И. Кичангина). Самка VI стадии с такими икринками добыта 4 IV 1947 на южном склоне Медвежинской банки. 10—13 V 1951 одна самка длиной 109 см была во II—III стадии (вес гонад 1.92% от веса тела), одна длиной 117 см — в III (вес гонад 2.8%), две (длиной по 119 см) — в VI стадии с оставшимися после нереста крупными икринками (вес гонад 1.57 и 1.20%); вес гонад у самцов от 0.02 до 0.22%, в среднем 0.09% (73°10' с. ш., 30—33° в. д.; собственные наблюдения).
Самки созревают, по Г. И. Милинскому (1944), при длине не менее 80 см. Диаметр зрелых икринок 7.25—8.0 мм (Расс, 1941 : 31). Плодовитость, по П. З. Сахно (Маслов, 1944 : 156), у самки длиной 112 см — 23 485, длиной 124 см — 29 290, 134 см — 28 380 икринок (Баренцево море). Кладки икры нигде еще не найдены.
Данному виду приписывается одна личинка длиной 24 мм, найденная у Вардё (Collett, 1879 : 58) и 2 длиной 21 и 28 мм, найденные у входа в залив Св. Лаврентия (первая у поверхности, при температуре +11.5° м солености 27.83‰, вторая на глубине 25—125 м, — Dannevig, 1919 : 16), но достаточных оснований для этого определения не имеется. Все признаки, приведенные в описании Колетта, свойственный личинкам Anarhichas lupus, за исключением однообразной темно-коричневой окраски, которую, однако, и последние принимают на свету (Mcintosh, 1890 : 288). Единственным основанием для определения личинок из залива Св. Лаврентия послужило расположение зубов сошника, которое, однако, не описывается; Данневиг, очевидно, полагал, что расположение зубов у личинок зубаток и взрослых особей сходно, но в действительности это не так, насколько можно, судить по личинкам Anarhichas lupus lupus. Большие размеры личинок также ни о чем не говорят: как выяснено, при выходе из икры личинки Anarhichas lupus могут достигать длины 25 мм. Таким образом, видовая принадлежность указанных личинок неясна. Судя по месту их находок, они, вероятнее всего, принадлежат Anarhichas lupus.
Молодь Anarhichas latifrons длиной менее 70 см редко встречается в уловах во всех частях ареала, а у берегов Исландии вообще неизвестна (Saemundsson, 1949 : 34). За пределами Баренцева моря не было отмечено ни одного экземпляра длиной менее 52 см, если не считать экземпляр длиной 41.3 см из Дэвисова пролива, описанный Пичманом (Pietschmann, 1932 : 54) — судя по изменениям и однообразной бледно-коричневой окраске в спирте, это Anarhichas lupus lupus — и экземпляр длиной 6 см из желудка трески (Kotthaus u. Krefft, 1957 : 177), описание которого не дается. В Баренцевом море, а также у о. Медвежьего и западного побережья Шпицбергена с 1941 по 1950 г. сотрудниками ПИНРО было обнаружено 55 особей Anarhichas latifrons длиной 25—65 см (3.5 % от общего числа выловленных). Единственный случай поимки мальков длиной 11—13.5 см отмечен Есиповым (1949 : 616) в июле у о. Медвежьего. К сожалению, ни одной особи Anarhichas latifrons длиной менее 65 см в коллекциях ПИНРО не сохранилось, описаний их не имеется и проверить определение невозможно. Anarhichas latifrons нерестится, по всей вероятности, на больших глубинах материкового свала, как это и предполагает Г. И. Милинский. Поэтому ежегодность нереста маловероятна, так как в течение круглого года наблюдаются значительные для этого вида скопления в таких отдаленных от материкового свала районах, как Гусиная банка. Основная масса молоди, по-видимому, также держится на больших глубинах материкового свала. К тому же молодь, судя по слабой приспособленности ее зубов и губ к питанию донной фауной по сравнению со взрослыми особями, по всей вероятности, гораздо чаще последних держится в толще воды, не будучи связана с дном. Очевидно, в силу указанных причин она редко попадает в тралы.
В кишечниках синей зубатки в Баренцевом море найдены офиуры — Ophiura sarsi, Ophiopholis aculeata, морские звезды — Asterias linki, Ctenodiscus crispatus, Solaster endeca, Solaster gracilis, морские ежи — Strongylocentrotus droebachiensis, голотурии, крабы — Hyas sp., раки-отшельники — Eupagurus pubescens, Eupagurus bernhardus, креветки — Pandalus, брюхоногие моллюски — Buccinum sp., двустворчатые — Cardium sp., крылоногие моллюски — Clione limacina, осьминоги — Rossia, сцифомедузы — Cyanea arctica, гребневики — Beroe cucumis, черви — Nereis, Aphrodite, актинии, гидроиды, губки, рыба — мойва, молодь тресковых, камбала-ерш, морской окунь, пинагор и отбросы с траулеров. Преобладают иглокожие; ракообразные и моллюски встречаются гораздо реже. Обычна рыба, но неизвестно, какую часть рыбы Anarhichas latifrons ловит, а какую подбирает из выброшенной с траулеров вместе с другими отбросами (кишечники, позвоночники, головы трески и пикши и т. д.), поедаемыми ею. Нередко кишечники синих зубаток бывают набиты гребневиками и медузами (Thilemann, 1921 : 206; Ehrenbaum, 1936 : 141; Есипов, 1949 : 614; данные ПИНРО; собственные наблюдения), что резко отличает Anarhichas latifrons от других зубаток. У берегов Гренландии в желудках немногих исследованных экземпляров найдены морские звезды, морские ежи и крабы; на Ново-Шотландских банках, у 1 экз. — морские ежи (Cornish, 1907 :87).
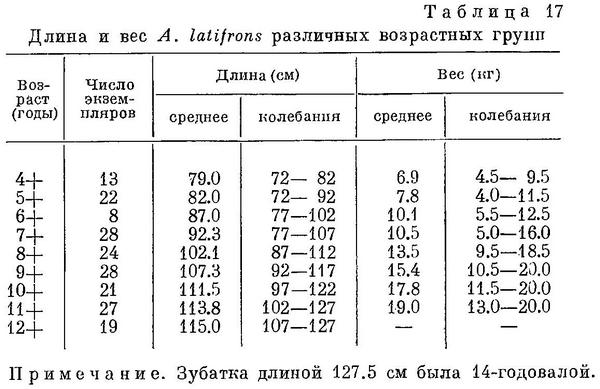
Темп роста Anarhichas latifrons в Баренцевом море, по данным П. З. Сахно (Маслов, 1944 : 155), наибольший по сравнению с другими видами зубаток, особенно в первые 4 года жизни (см. табл. 17). Впрочем, не исключена вероятность, что в действительности прирост за первые 4 года у основной массы особей меньше, чем это вытекает из данных табл. 17: вполне возможно, что среди молодых особей в первую очередь вылавливаются наиболее быстро растущие экземпляры, тогда как тугорослые дольше держатся в тех местах, что и остальная молодь размером до 60—70 см и, таким образом, дольше избегают вылова. Если это так, то средние длины, указанные в табл. 17, приближаются к максимальным длинам основной массы особей данной возрастной группы тем больше, чем моложе группа. Колебания численности Anarhichas latifrons, по-видимому, сходны с таковыми Anarhichas minor.
Синяя зубатка была встречена мной в кишечнике полярной акулы (Somniosus microcephalus). Экземпляр длиной 6 см (описание его не дается) найден в желудке трески (Kotthaus u. Krefft, 1957 : 177).
Ю. И. Полянский (1955) обнаружил у баренцевоморских Anarhichas latifrons (3 экз.) следующих паразитов: Myxoproteus elongatus (мочевой пузырь), Podocotyle reflexa (кишечник), Neophasis lageniformis (кишечник), Steringophorus furciger (кишечник), Diphterostomum microacetabuum (кишечник), Lepidophyllum steenstrupii (мочевой пузырь), Lecithaster gibbosus (кишечник), Derogenes varicus (желудок), Scolex polymorphus (кишечник), Pyramicocephalus phocarum (полость тела), Contracoecum aduncum (кишечник), Anisakis sp. larvae (поверхность тела, мезентерии, кишечник), Echinorhynchus gadi (кишечник), Clavellodes rugosa(жабры), Sphyrion lumpi (кожа). Fellodistomum fellis, весьма обычный у Anarhichas minor и Anarhichas lupus, не обнаружен, возможно, не только в силу малого числа наблюдений, но и вследствие того, что Anarhichas latifrons редко посещает прибрежную полосу, сравнительно неширокую, в которой создается контакт между первым (Nucula tenuis) и вторым (Ophiura sarsi) промежуточными хозяевами (Полянский, 1958 : 85).
Хозяйственное значение гораздо меньше, чем у пятнистой зубатки, не только в силу меньших уловов, но и в связи с низким качеством мяса (водянистое). Мясо синей зубатки содержит 90—92% влаги, всего лишь 5—7% белка и 0.7—1.8% жира (Травин, 1952 : 175). Содержание витамина А у синей зубатки, по данным ПИНРО, выше, чем у пятнистой зубатки и трески, в среднем 2.57. Следует, однако, заметить, что содержание витамина А у рыб сильно колеблется по временам года (Суворов, 1948 : 97).
У Anarhichas latifrons, в отличие от других зубаток, нижняя челюсть несколько укорочена. Когда рот закрыт, передние большие клыки нижней челюсти не вклиниваются между такими же клыками верхней, а располагаются позади них (табл. VII).
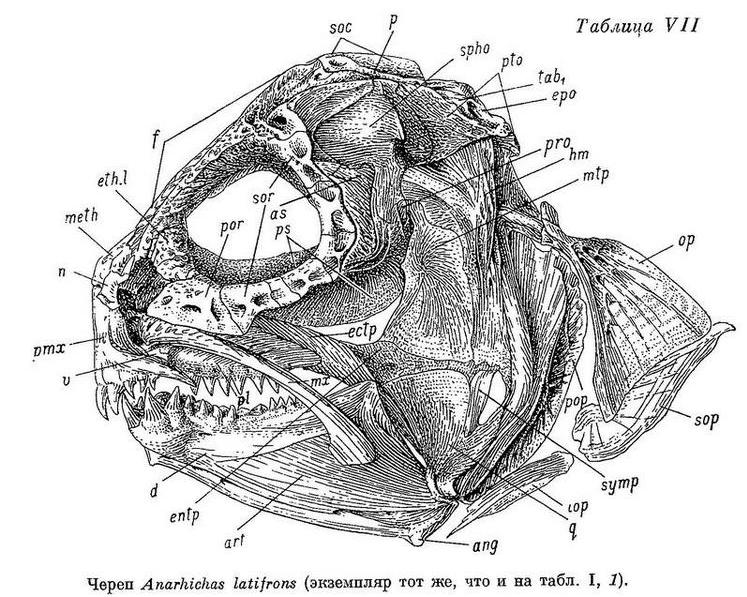
Все клыки мельче, острее, чем у других видов, почти не стираются; разница между большими и малыми клыками на обеих челюстях меньше. Очевидно, клыки синей зубатки мало приспособлены к отрыванию организмов от субстрата и служат главным образом для удержания более подвижной плавающей добычи. Большая часть зубов на нёбных костях (а иногда и все) высокие, конические. Из таких же конических зубов составлен внешний ряд зубов на нижнечелюстных костях (за площадкой), а также часть внутреннего ряда зубов, расположенная кзади от внешнего. Все эти зубы работают в паре с зубами нёбных костей. Внешний ряд зубов нёбных костей расположен внутрь от внешнего ряда зубов нижнечелюстных. Зуб одного из этих рядов помещается напротив зуба другого так, что их боковые поверхности при закрытом рте оказываются рядом друг с другом (обычно у переднего зуба нёбных и заднего клыка нижнечелюстных они почти соприкасаются, по направлению же назад дистанции увеличиваются). Передние зубы внутреннего ряда нёбных располагаются между внешним и внутренним рядами зубов нижнечелюстных. В целом образуются своего рода «зубчатые ножницы», отрывающие и разрывающие лучи морских звезд и офиур. Все конические зубы мельче и острее, стираются меньше, чем у других зубаток, а поэтому могут лучше служить и для схватывания более подвижных животных, лишенных панциря. Особенно малы нижнечелюстные давящие зубы по сравнению с этими зубами у других видов. Складчатость их оснований, как и у других зубов, выражена слабее, а полость пульпы развита сильнее (табл. VI, 1): скрепление зубов с цоколями менее прочное.
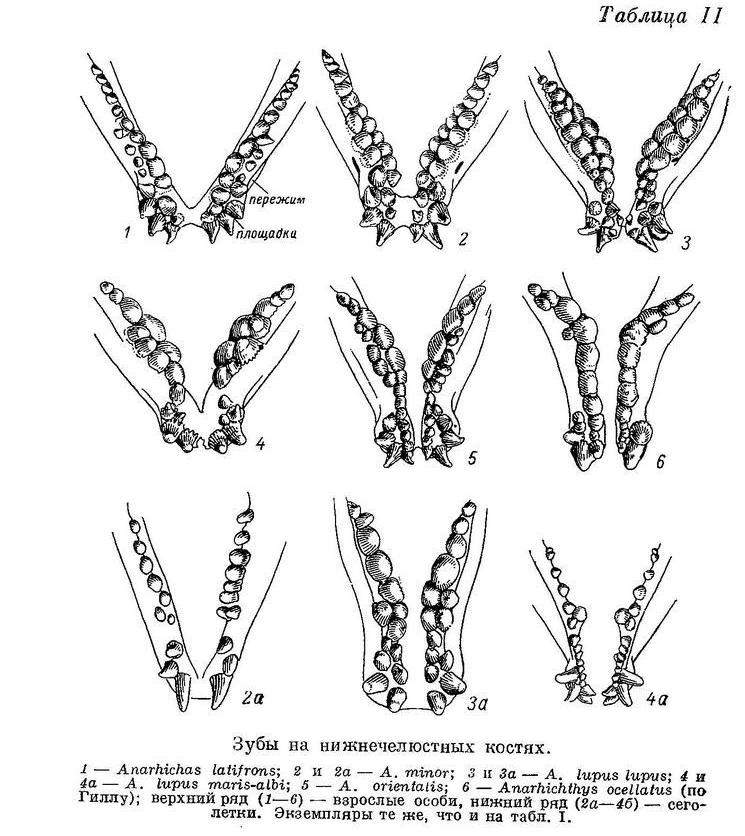
На нижней челюсти наибольшие давящие зубы расположены, как и у других видов зубаток, в средней части внутреннего ряда зубов, но сдвинуты вперед, ближе к клыкам, помещаясь обычно на самой площадке и непосредственно за нею (табл. II, 1).
Соответственно работающие с ними в паре зубы сошника образуют лишь короткие ряды. Эти ряды значительно короче нёбных рядов (табл. I, 1), поскольку зубы на нёбных костях простираются назад почти столь же далеко, как и конические зубы на нижнечелюстных костях, образующие в паре с зубами на нёбных «зубчатые ножницы».
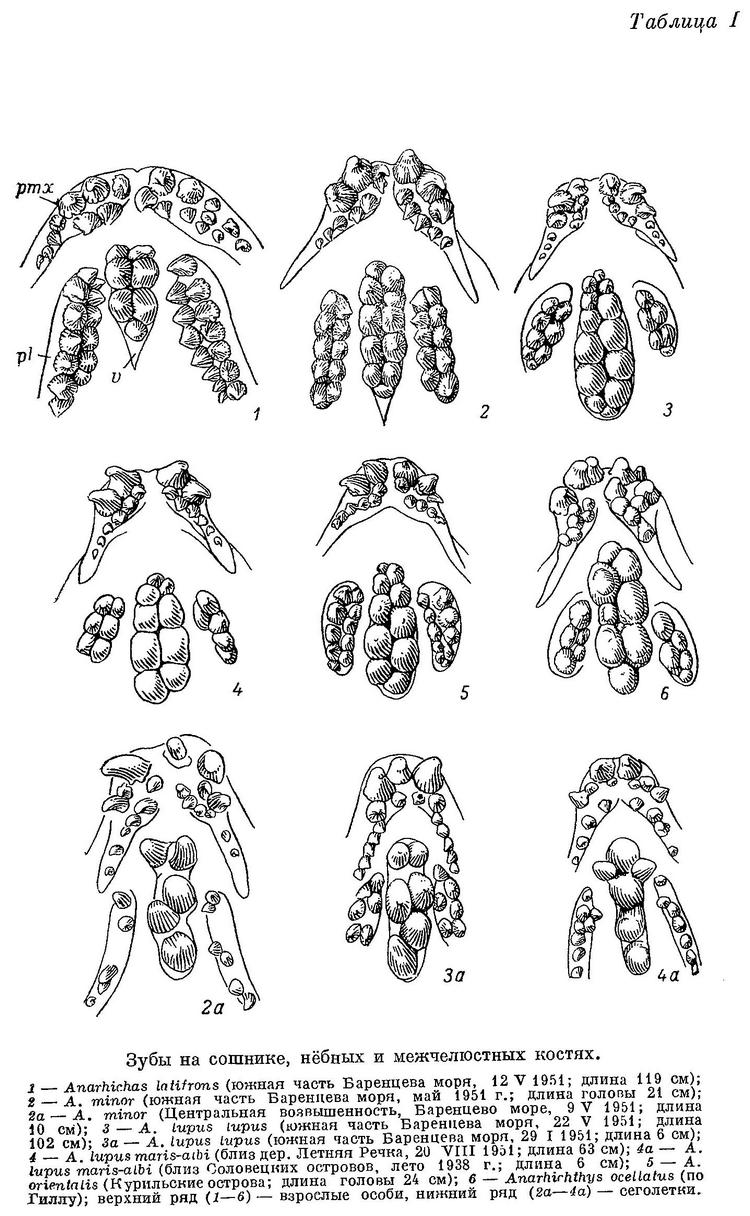
Задние зубы внутреннего ряда на нёбных костях нередко тупоконические, ниже передних и располагаются напротив или даже чуть внутрь от задних давящих нижней челюсти, образуя функционально как бы продолжение назад зубов сошника; однако не у всех особей это выражено одинаково хорошо, а у некоторых и совсем не выражено. Наибольшие давящие зубы сошника обычно расположены ближе к его переднему концу, чем к заднему (табл. I, 1); эти зубы такие же слабые, как и на нижнечелюстных костях. Пережим на нижнечелюстных костях развит мало (табл. II, 1), опускание вниз нижнечелюстных давящих зубов и наклон их внутрь почти не выражены. По-видимому, Anarhichas latifrons может раздавливать морских ежей и крабов лишь со сравнительно слабым панцирем, а моллюсков лишь мелких, с тонкой раковиной.
Не менее интересна возрастная изменчивость в строении зубной системы.
К сожалению, в моем распоряжении не было Anarhichas latifrons длиной менее 60 см. (Здесь и всюду далее, где упоминается длина рыбы, речь идет об абсолютной ее длине.) У молодых особей этого вида длиной 60—70 см малые клыки по величине лишь немного отличаются от больших. Описание дано по 3 экз. из Баренцева моря. Lycichthys paucidens (Gill, 1911 : 167, fig. 9, 10, pl. 18) также представляет собой, по моему мнению, молодой экземпляр Anarhichas latifrons, так как имеет строение зубов, очень сходное с молодью последнего вида. Различие состоит лишь в том, что зубов, расположенных между двумя обычными рядами, у него нет: по-видимому, этот ювенильный признак был утрачен им раньше, чем другие. Все зубы внешнего и следующие за ними зубы внутреннего ряда нижнечелюстных, а также многие зубы нёбных имеют форму клыков, вершины их не прямые, а загнуты внутрь и назад (табл. V, 1).
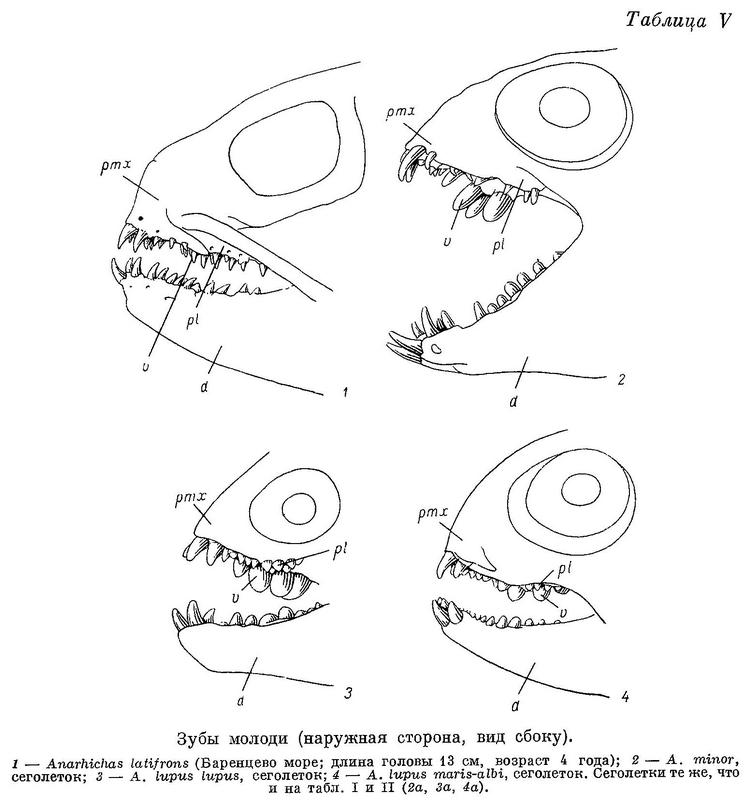
Зубы внутреннего ряда на нижнечелюстных костях, расположенные рядом с зубами внешнего, и зубы на сошнике не округлые, а высокие, конические. Ряд зубов на сошнике имеет форму треугольника, обращенного основанием вперед. Ряды зубов на всех костях менее правильны, чем у взрослых особей. Имеются зубы в промежутках между двумя крайними рядами, что редко бывает у взрослых. Вершины зубов нёбных костей при закрытом рте слабо заходят за вершины зубов нижнечелюстных. Боковые поверхности этих зубов, более или менее параллельные у взрослых, расположены у молоди под большим углом друг к другу. Следовательно, «зубчатые ножницы» образованы не внешним и задней частью внутреннего ряда зубов нижнечелюстных и зубами нёбных костей, как у взрослых, а зубами сошника и передней частью внутреннего ряда зубов нижнечелюстных костей, т. е. зубами, которые у взрослых принимают на себя давящую функцию. Судя по строению зубов, у молоди Anarhichas latifrons основную роль в питании играют подвижные организмы, лишенные панциря, что, вероятно, еще резче выражено у молоди меньших размеров. Переход к питанию крупными иглокожими и мелкими моллюсками, очевидно, происходит поздно, при длине не менее 70—80 см, т. е. в возрасте не менее 4 лет.

Примечания. 1) В этой и следующих таблицах цифры, выделенные курсивом, указывают количество экземпляров. 2) Для Anarhichas latifrons измерялись особи длиной более 100 см.
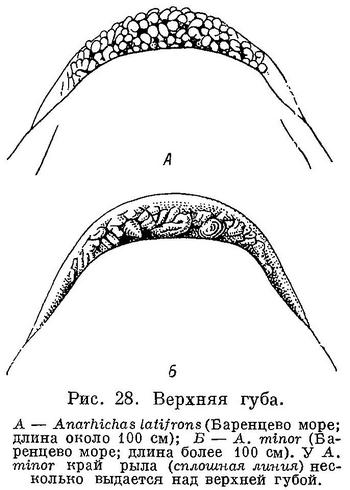
У всех зубаток губы устроены довольно сходно, но у Anarhichas latifrons задняя часть нижней губы выдается вверх в виде клапана, которому соответствует небольшое углубление на верхней губе. Возможно, этот клапан препятствует выскальзыванию изо рта гребневиков и медуз, составляющих обычный объект питания синей зубатки. У всех зубаток поверхность губ спереди, перед клыками, покрыта складками, похожими на бахрому, но у Anarhichas latifrons вместо бахромок развиты хорошо выраженные сосочки, а сами губы гораздо толще (табл. 2).
По-видимому, эта особенность в строении губ вызвана большей их ролью в отыскании пищи. Последнее может быть вызвано уменьшением роли зрения в связи с обитанием на наибольших глубинах по сравнению с другими зубатками. Нижняя губа у Anarhichas latifrons значительно тоньше верхней, притом нижняя челюсть и без того несколько укорочена. Укорочение нижней челюсти и преимущественное развитие верхней губы свидетельствуют о том, что взрослые особи Anarhichas latifrons берут пищу преимущественно со дна. У молоди нижняя челюсть не столь укорочена, губы тоньше, верхняя губа по толщине меньше превышает нижнюю, сосочки развиты слабее — и все это несмотря на большие глубины обитания, чем у взрослых; таким образом, доставляется еще одно доказательство меньшей роли донной фауны в питании молоди. Не исключена возможность, что молодь Anarhichas latifrons ведет батипелагический образ жизни и у дна встречается реже, чем взрослые особи.
Окраска
У Anarhichas latifrons общий фон серый со светло-фиолетовым оттенком или с разной интенсивности бурыми тонами вплоть до шоколадного. Бока, спина, иногда верх головы и спинной плавник покрыты очень расплывчатыми пятнами, обычно слабо отличающимися от общего фона. По Есипову (1937 : 49, 52), у некоторых особей эти пятна расположены в виде слабо выраженных поперечных полос. На плавниках пятна обычно незаметны.
Радужина чаще всего сплошь темно-бурая, лишь изредка с небольшими желтыми пятнами или крапинками; таким образом, ее окраска значительно меньше зависит от освещения, чем у предыдущих видов. Судя по всему, это объясняется обитанием Anarhichas latifrons на больших глубинах. Последнее, а возможно, также и предпочтение к илистым грунтам обусловливают однотонность общей окраски данного вида.
Дополнение к разделу «Anarhichas latifrons». На Нью-Фаундлендских банках, по данным ПИНРО, Anarhichas latifrons встречается реже, чем другие виды зубаток; вдоль побережья Лабрадора по направлению к западу численность этого вида возрастает и он начинает преобладать; встречен здесь до 60°14' с. ш. и 61°50' з. д. Экземпляр длиной 82 см выловлен возле о. Ян-Майен.
В марте 1959 г. самки III стадии зрелости (8 экз.) в Баренцевом море встречены только вблизи свала (банка Копытова), при температуре примерно 3—5°; восточнее 18° в. д. ловились лишь самки I, I—II, II, II—III и VI—II стадий, по большей части при более низких температурах. Оба пелагических малька Anarhichas latifrons, по данным А. С. Бараненковой и Н. С. Хохлиной, выловлены в мае, также поблизости от свала (74°20' с. ш. и 14°00' в. д.; 73°10' с. ш. и 16°38' в. д.).
Рыбы северных морей СССР. А.П.Андрияшев 1954 г.
1. Lycichthys denticulatus (Kröyer) — Синяя зубатка, вдовица, синюха.
Anarrhichas latifrons Steenstrup, Forh. Scand. Naturf., 3 Mote (1842), 1843 : 647 (Исландия; nom. nudum). — Steenstrup et Hallgrimsson in.: Steenstrup, Vidensk. Meddel. Naturh. Foren., (1876), 1877 : 177, 201, tab. III, fig. 3 (Гренландия, Исландия). — Collett, Christ. Vidensk.-Selsk. Forhandl., (1879), 1, 1880 : 46, pl. (fig.) II (Финмаркен; сравнение с другими видами). — Smitt, Scand. Fish., I, 1893 : 237, pl. XIII, fig. 2 (цветн. рис.). — Gооdea. Bean, Ocean. Ichth., 1895 : 301, fig. 271 (42°55'30" с. ш., 50°51' з. д., глуб. 471 саж.). — Книпович, Определитель, 1926 : 94, фиг. 60 (Баренц. м. до Нов. Земли; дл. до 128 см). — Есипов в изд.: Промысл. рыбы СССР, 1949 : 616, атлас табл. 189 (сводка, цветн. рис.).
Anarrhichas denticulatus Kröyer, Overs. Dansk. Vidensk. Selsk. Forhandl., 1844 : 139 (Гренландия); Gaimard's Voy. Scand. Lapp. Spitzb. Feroe, Paris, 1845 (?) (Год издания на атласе рисунков (С. Thornam и др.) не проставлен. Коллетт (1905 : 116) принимает 1845 г.; Дин (Dean, Bibliogr. Fish., I, 1916 : 435) указывает 1842 г. (все издание 1842—1856).) : pl. 12 (цветн. рис., зубы). — Günther, Cat. Fish., III, 1861 : 211 (Гренландия; D 80). — Collett, Rep. Norw. Mar. Fish.-Invest., II, 3, 1905 : 114 (синонимия).
Anarrhichas fortidens Gill, 1911 : 166, fig. 8 (sec. Goode a. Bean, 1895; зал. Мэн и Ной, Шотландия).
Распространение, биология. Книпович, Эксп. научно-пром. иссл. Мурмана, I, 1902 : 55 и сл., фиг. 7 (многочисленные местонахождения у берегов Мурмана; фотогр.). — Dannevig, Canad. Fish. Exp. 1914—1915, Ottawa, 1918 : 16 (зал. Св. Лаврентия; пелагические личинки 21—25 мм). — Thielemann, Wiss. Meeresunters. (Helgol.), XIII, 2, 1921 : 206 (Мурман; питание). — Jensen, Rap. Proc.-Verb., XXXIX, 1926 : 99 (зап. Гренландия). — Saemundsson, Synops. Fish. Iceland, 1927 : 18 (вокруг Исландии на глубине 100—800 м). — Рiеtsсhmann, Meddel. Gronl., XCII, 3, 1932 : 53 (Дэвисов прол.). — Bigelow a. Schroeder, Proc. Bost. Nat. Hist., XLI, 2, 1935 : 15, pl. 3 (Ново-Шотландск. банки под 44° с. ш., хороший рис. зубов). — Merriman, Copeia, 3, 1935 : 143 (Ново-Шотландск. банки; дл. 118.5 см, вес около 15 кг, фотогр.). — Есипов, Промысл. рыбы Баренц. м., 1937 : 51, 107, фиг. 17 (промысел). — Маслов, Тр. Полярн. научно-иссл. инст. рыбн. хоз. океаногр., VIII, 1944 : 154 (темп роста, плодовитость, промысел).
D 77—80, А 45—47, Р 20—22, позвонков 79—81.
Голова заметно заостряется к рылу. Зубы на челюстях более заостренные, многочисленные и реже расставленные, чем у других зубаток. Сошниковый ряд зубов очень короткий (простирается назад значительно меньше нёбного ряда), его длина составляет лишь 2/3—3/4 длины нёбного ряда зубов. Грудной плавник короткий, менее 12% L. Длина хвостового плавника содержится более 10 раз в длине туловища (без головы и С). Тело темное, серовато-бурого или шоколадного цвета, однотонное или с неясными темными пятнами. Длина в Баренцовом море до 134 см и вес до 20 кг, чаще длина 70—110 см и вес 4—15 кг.
Этот вид чаще известен в литературе под названием Anarhichas (или Lycichthys) latifrons Steenstr. et Hallgr., однако давно уже был доказан приоритет видового названия Anarhichas denticulatus Kröyer, 1844 (см. у Коллетта, а также у Гюнтера и Смитта), так как Anarhichas latifrons, упомянутая без описания и рисунка в 1843 г. для Исландии, фактически была впервые описана лишь в 1876 г. по экземпляру из Гренландии (см. синонимию), откуда происходит и тип описанной ранее Anarhichas denticulatus Kr. Таким образом, нет оснований сохранять название Стенструпа. Впоследствии описанные американские виды Lycichthys (Гилл) до сего времени ближе не изучены. Из них Lycichthys fortidens Gill вряд ли отличается от обычной Lycichthys denticulatus, другой же вид [Lycichthys paucidens (Kr.)] имеет лишь один ряд нижнечелюстных зубов, что у других взрослых зубаток не было обнаружено. Следует отметить, что вообще точное сравнение различных видов зубаток по ряду пластических признаков, строению задней части спинного плавника, а также по особенностям зубного вооружения в настоящее время еще затруднительно из-за слабой изученности их возрастной изменчивости, которая у зубаток при их крупных размерах должна быть весьма значительной.
Распространение. Почти все Баренцево море, в промысловых количествах на север до Медвежинско-Шпицбергенского района и на восток до Канинских банок, единично до Новой Земли. В Белом море отсутствует. У берегов Норвегии на юг доходит до Лофотенских островов, но известна и южнее — на глубинах у Фарерских и Шетландских островов (Однажды найдена у берегов Великобритании (см.: Bigelow a. Schroeder, 1935 : 17).); обычна вокруг Исландии, а также в Дэвисовом проливе у Гренландии. У американских берегов попадается реже, хотя неоднократно ловилась в районе Ново-Шотландских банок и южнее почти до м. Код.
Биология. Встречается на глубине от 75 до 800 м, в Баренцовом море до 425 м, у берегов Норвегии и Фарерских островов 300—600 м. Предпочитает илистые грунты, высокие солености (не менее 34.5‰) и близкие к нулю положительные температуры (в Баренцовом море до 2—3°, в более южных районах до 5—6°), но иногда встречается и при температурах ниже нуля. Миграции и места нереста не изучены. Плодовитость 23 485—29 290 икринок (у 3 баренцовоморских экземпляров длиной 112—134 см, по подсчетам П. 3. Сахно; см. у Маслова, 1944). Диаметр икринок 7—8 мм. Темп роста в Баренцовом море (по данным П. 3. Сахно) следующий:

Питание синей зубатки составляют преимущественно иглокожие (офиуры, Ctenodiscus, Strongylocentrotus), двустворчатые моллюски (Pecten islandicus, Cardium ciliatum), отчасти рыбы.
Хозяйственное значение, несмотря на значительные уловы, меньше, чем пятнистой зубатки, в связи с низким качеством мяса (водянистое). Отдельной статистики уловов нет (см. Anarhichas minor).