(† Eugeneodontiformes Zangerl, 1981) Отряд Евгенеодонтообразные, Евгенеодонтиды, Order † Eugeneodontiformes Zangerl, 1981 4 семейства
Синонимы:
Eugeneodontida Zangerl, 1981
Материал подготовил Фактистов И. Е., 2021 г. Сайт www.fishbiosystem.ru
Евгенеодонтообразные, Евгенеодонтиды (Eugeneodontiformes) — отряд вымерших, преимущественно верхнепалеозойских, хрящевых рыб (Chondrichthyes Huxley, 1880), относящийся к классу Цельноголовые (Holocephali Bonaparte, 1832). Они обладали уникальным «зубчатым венчиком» на симфизе нижней челюсти и грудных плавниках, поддерживаемым длинными лучевыми костями. Palatoquadrate был либо слит с черепом или уменьшен. (У некоторых рыб нёбно-квадратный хрящ (palatoquadrate) является дорсальным хрящом нижней челюсти, а вентральный — меккелевским хрящом.) Представители отряда близки к общим предкам химер и акул, иногда считаются «настоящими» акулами и сближаются с кладоселяхиями († Cladoselachiformes).
Значение имени Eugeneodont коррелирует с «зубами истинного происхождения» и происходит от греческого ευ — eu (хороший / истинный), γενεος — geneos (раса, род, происхождение) и οδον — odon (зуб).
Евгенеодонтообразные — это морские специализированные хрящевые рыбы. Строение зубной системы и тип питания разнообразны, но в основном характерно наличие давящих зубов. Нейрокраний, в тех случаях, где он известен, относительно узкий, с вытянутой преорбитальной частью, короткой отико-окципитальной (заглазничной). Может присутствовать длинный ростр. Нёбно-квадратный хрящ (palatoquadrate) небольшой или редуцирован, при этом верхнечелюстные зубы смещены на нижнюю поверхность нейрокрания. Нёбно-квадратный хрящ, по-видимому, прирастал к нейрокранию (как у химер). Может присутствовать своеобразный нижнечелюстной ростр, расположенный впереди меккелева хряща.
Зубная система гетеродонтная. Гетеродонтизм (или гетеродонтность, также гетеродонтия) — наличие у животного зубов, различающихся по форме и функциональному назначению. Боковые зубы обычно давящие, многовершинные, в виде поперечных рядов, разного размера на протяжении челюсти (например, у † Caseodus и † Fadenia наиболее крупные по размерам зубы расположены примерно посередине ветви нижней челюсти). Иногда, по-видимому, боковые зубы могут быть утрачены.
Самая выдающаяся особенность группы — наличие у большинства представителей так называемых «зубных спиралей». Собственно, иногда это просто дугообразный ряд симфизных зубов, но у ряда родов известны настоящие спирали, содержащие до 130 зубов. Положение такой спирали на симфизе оспаривается некоторыми учёными (есть, например, мнение, что спираль каким-то образом крепилась в районе глотки, являясь производным глоточных зубов — вернее, зубчиков слизистой оболочки глотки, подобные структуры есть у костных рыб и примитивных акул). В то же время, известны черепа с явно сохранившимися в прижизненном положении симфизными зубами. У некоторых форм, вероятно, присутствуют крупные симфизные дуги в обеих челюстях. Дуги и спирали были непарными, то есть образовывали один ряд зубов. Симфизные зубы могут быть либо давящими (и в этом смысле мало отличаться от боковых зубов), либо крупными, остроконечными. Характерно развитие на симфизных зубах так называемых «шпор» — крупных парных выростов у основания зуба, в итоге зубы как бы «вкладывались» один в другой, иногда образуя довольно жёсткую конструкцию. Направление «шпор» считалось диагностическим признаком (у геликоприонтид — вперёд, у эдестид — назад), но не исключено, что шпоры были всегда направлены назад. В таком случае самые мелкие зубы дуги — молодые, самые крупные — оттеснённые ими вперёд более поздние. Такой тип роста зубов известен и для современных акул. Иногда симфизные зубы гетеродонтны (более мелкие — давящие, более крупные — высокие). Зубы сложены тубулярным дентином или остеодентином, покрытым паллиальным ортодентином и энамелоидом.
Жаберные дуги расположены позади мозговой коробки, как у акул, но их ряд относительно короткий, число — пять. Вероятно, при жизни были отдельные жаберные щели, присутствие жаберной крышки не доказано. Посткраниальный скелет наиболее полно известен для рода † Fadenia (пермь Гренландии), внешний вид других представителей группы мог быть иным.
Тело веретенообразное, с довольно длинной хвостовой частью. Скапулокоракоид узкий, с парой дополнительных переднебрюшных хрящевых элементов. Грудные плавники с многочисленными радиалиями, несегментированными, крепящимися к скапулокоракоиду или элементам метаптеригиальной оси. Свободная часть метаптеригия выходит назад за пределы плавника (строение как у многих палеозойских акул). Спинной плавник один, прямо над плечевым поясом. Плавникового шипа нет, но присутствует крупный базальный элемент в основании плавника. Брюшных и анальных плавников нет. Хвостовой плавник полулунный, равнолопастный, невральные и гемальные дуги, а также базалии в его лопастях, слиты в крупные пластины. Есть мелкие чешуи в виде зубчиков или их комплексов. Внешний облик фадении соответствует пелагической рыбе, но озубление (включая симфизные зубы) — давящее. Скелет близкого рода Caseodus известен хуже, но очень похож по строению. Не исключено, что подобные рыбы питались аммонитами.
Образ жизни евгенеодонтов остается загадочным. Первые исследователи предполагали, что зубные дуги были плавниковыми шипами. Когда истинная природа этих образований была обнаружена, возникло предположение, что дуги и спирали использовались для добывания пищи на дне. В частности, для эдестуса предлагали использование дуг в качестве «ножниц» для отдирания моллюсков от субстрата. Для геликоприона спираль сравнивали с радулой брюхоногих моллюсков. Интересно, что первооткрыватель геликоприона А. П. Карпинский считал «спираль» частью ростра верхней челюсти. Этот взгляд сохранялся до 1970-х годов, особенно в советской литературе. Зубы на дугах и спиралях, по-видимому, не несут явных следов истирания — поэтому «вспахивание» ими дна или отдирание моллюсков от камней маловероятно. Существует мнение, что эти рыбы питались пелагическими аммонитами. Известные по скелетным остаткам евгенеодонты — тахипелагические рыбы, то есть быстроходные обитатели толщи воды. Евгенеодонты интересны, как самые крупные рыбы палеозоя, весьма разнообразная и долго существовавшая группа, пережившая даже пермо-триасовую «катастрофу».
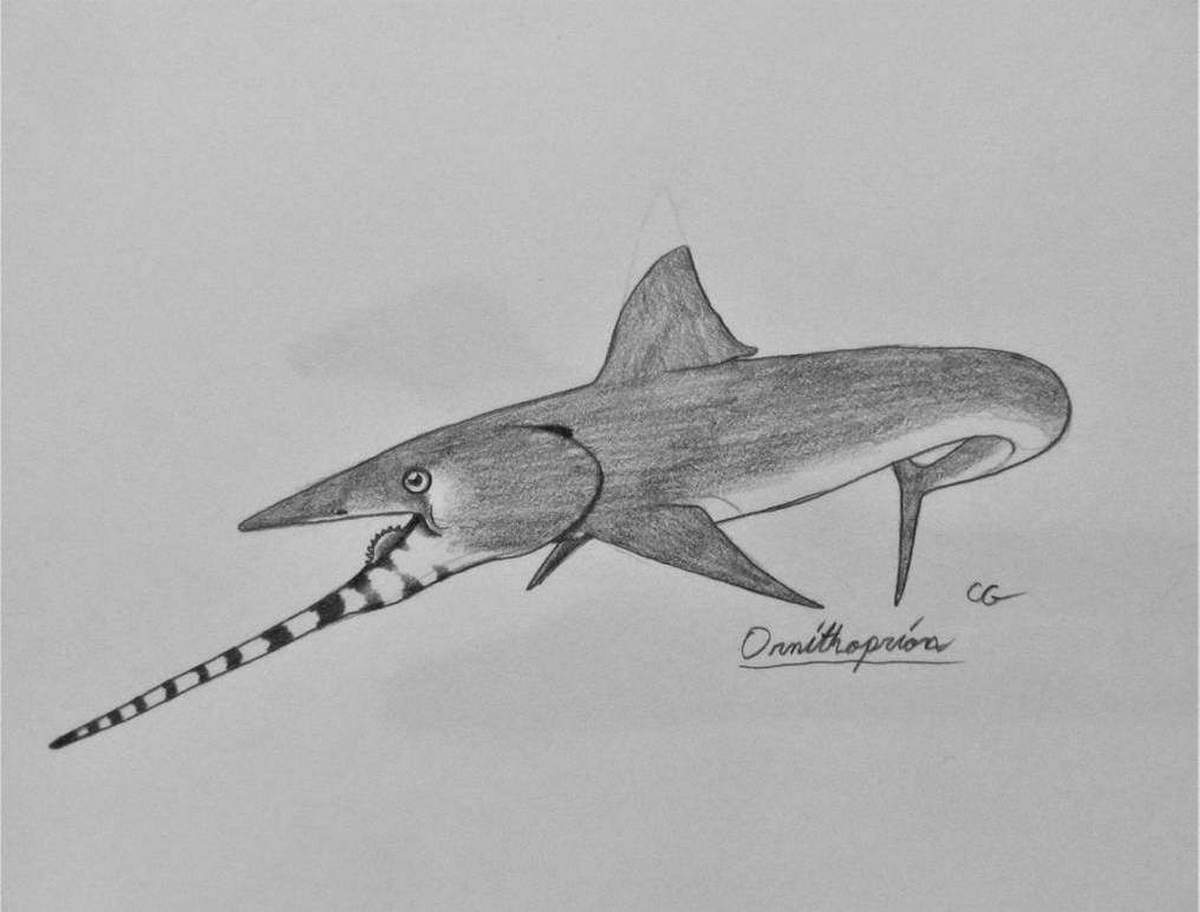
Надсемейство Казеодонты († Caseodontoidea) — включает в себя семейства † Eugeneodontidae и † Caseodontidae. Отличаются поперечно-гребенчатыми или вздутыми симфизными зубами. Сюда относится самый полно известный представитель группы — † Fadenia из карбона и перми Гренландии и Северной Америки. Это относительно некрупная (до 1,5 метров в длину) рыба. Род † Ornithoprion из среднего карбона Индианы — мелкая рыба (череп не более 10—15 см длиной) с необычным нижнечелюстным ростром. По-видимому, в этой группе не всегда формировалась настоящая симфизная спираль или даже развитые симфизные зубные дуги. Среди казеодонтов были очень крупные рыбы — известны челюсти † Caseodus до 60 см длиной, что может соответствовать общей длине до 4—5 метров. Всего в надсемейство входят примерно 4 рода, существовавших с карбона по пермь, в основном в Северном полушарии.
Надсемейство Эдестоиды († Edestoidea) — симфизные зубы с высокими режущими коронками. Также включает два семейства, известных почти исключительно по ископаемым частям зубного аппарата. † Helicoprionidae (или † Agassizodontidae) — геликоприоны («спиральные пилы»). Самый известный для широкой публики представитель — геликоприон. Симфизные зубы расположены на круто изогнутой дуге, у некоторых родов образуется полная спираль, с несколькими оборотами. Вероятно, «дуга» была лишь в нижней челюсти. Очень плохо известен, единственный представитель с более полными остатками — саркоприон († Sarcoprion edax) из перми Гренландии. Известен фрагмент черепа длиной около 50 см. В симфизе нижней челюсти, по-видимому, находилась дуга из высоких, сжатых с боков зубов, по сторонам от дуги — мозаика мелких давящих зубов; такие же зубы находятся на жевательных поверхностях обеих челюстей. Верхнечелюстной симфиз прямой, ростр длинный. На верхней челюсти в районе симфиза есть зубы, сходные с зубами нижнечелюстной дуги, расположенные, по-видимому, в 2 ряда. Возможно, при закрытой пасти нижнечелюстная дуга входила в углубление на верхней челюсти. Интересно, что у этого рода, судя по реконструкции Нильсена, шпоры симфизных зубов были направлены вперёд.
Род † Agassizodus из раннего карбона Северной Америки известен по симфизной дуге, состоящей из давящих зубов с высоким срединным килем. Род † Campyloprion известен из карбона Северной Америки и Подмосковья, отличается очень высокими зубами (до 10 см и более), образующими, по-видимому, полную спираль. Род † Parahelicoprion, известный из ранней перми Приуралья и Боливии имел короткую симфизную дугу с зубами более 10 см высотой.
В семейство входят очень крупные рыбы — известны спирали геликоприона и кампилоприона диаметром более 50 см. Возможно, владелец спирали превышал 6 метров в длину (есть указания на спирали большего размера). Остатки геликоприонов часто приурочены к геосинклинальным областям (рыба обитала в открытом море), но найдены и в прибрежных морских отложениях. Представители семейства (примерно 6—7 родов) известны из раннего карбона — поздней перми всех материков.
Семейство † Edestidae — эдесты («пожирающие»). Две симфизных дуги (по 10 зубов в каждой) — как считается, в верхней и нижней челюстях. В действительности, прижизненное положение дуг неизвестно. Коронки симфизных зубов режущие. Вероятно, зубы вытеснялись спереди назад и затем выпадали (опять же, есть гипотеза о происхождении зубных дуг из жаберных и положении первых в районе глотки, подобные глоточные зубы известны у костных рыб). Боковые зубы давящие или вообще отсутствуют. Шпоры симфизных зубов всегда направлены назад. Самый знаменитый представитель — эдестус. Рыбы очень крупных размеров, известны зубные дуги до 50 см длиной. Предполагается, что собственно эдестус мог быть пелагическим хищником, питавшимся другими хрящевыми и костными рыбами. Остатки приурочены к прибрежным отложениям, иногда даже к зонам каменноугольных бассейнов. Примерно 9 родов, известны от среднего карбона до раннего триаса всех материков. Род † Helicampodus, например, описан из поздней перми Индии и раннего триаса Армении.
В эту группу иногда включают загадочную рыбу † Listracanthus, известную по игловидным кожным зубчикам из карбона-триаса Северного полушария. Есть сведения, что полный экземпляр (обнаруженный доктором Р. Зангерлом в карбоне Северной Америки, но потом утерянный) имел змеевидную форму и огромные размеры, иглы покрывали спинную поверхность рыбы. Это показывает, что, возможно, евгенеодонты были весьма разнообразны по форме тела.
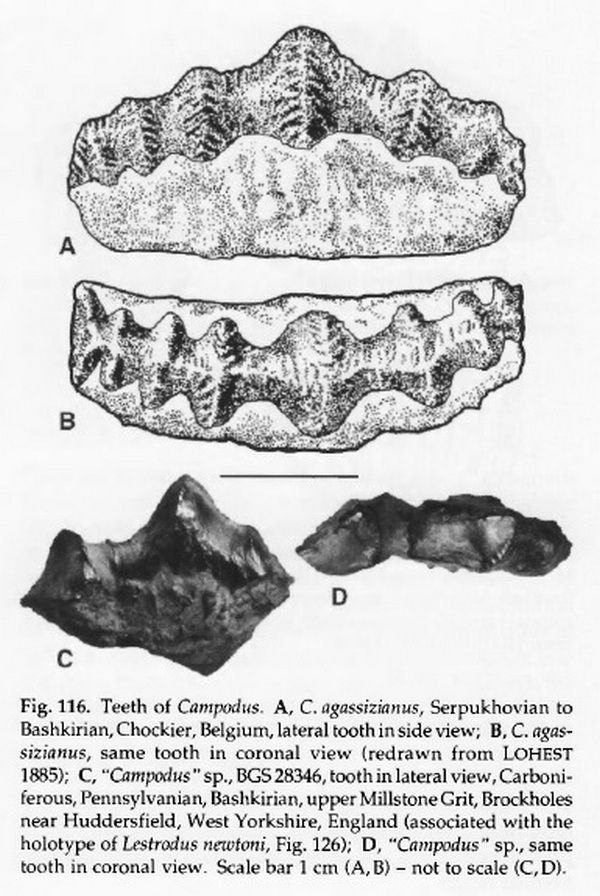
Род † Campodus, сближаемый с эвгенеодонтами, известен преимущественно по зубам из карбона Северного полушария. Зубы такого типа характерны и для других представителей группы, но к роду † Campodus относят также и скелетные остатки. Интересно, что относительно хрупкие гребни на зубах этого рода могут свидетельствовать о питании мелкими животными. У особи около 1 метра длиной, известной по отпечатку передней половины тела, зубы были 15 мм в поперечнике. В то же время, в коллекциях есть зубы до 10 см в поперечнике. Рыба могла быть 13 метров в длину.
В 2008 году практически полный череп евгенеодонта средних размеров описан из раннего триаса Канады. Нейтронное сканирование показало наличие симфизной дуги в районе ростра нижней челюсти, симфизные зубы высокие. Интересно, что сканирование подтвердило отсутствие настоящей верхней челюсти — верхние зубы крепились к нейрокранию.
 |
Надсемейство † Caseodontoidea Zangerl, 1981
• Род † Campodus Koninck, 1844
• Род † Chiastodus Trautschold, 1879
• Семейство † Caseodontidae Zangerl, 1981
• Род † Caseodus Zangerl, 1981
• Род † Erikodus Nielsen, 1952
• Род † Fadenia Nielsen, 1932
• Род † Ornithoprion Zangerl, 1966
• Род † Romerodus Zangerl, 1981
• Семейство † Eugeneodontidae Zangerl, 1981
• Род † Bobbodus Zangerl, 1981
• Род † Eugeneodus Zangerl, 1981
• Род † Gilliodus Zangerl, 1981
Надсемейство † Edestoidea Hay, 1930
• Семейство † Helicoprionidae Karpinsky, 1911 (синоним Agassizodontidae Zangerl, 1981)
• Род † Agassizodus St. John & Worthen, 1875
• Род † Arpagodus St. John & Worthen, 1875
• Род † Campyloprion Eastman, 1902
• Род † Helicoprion Karpinsky, 1899
• Род † Parahelicoprion Karpinsky, 1924
• Род † Sarcoprion Nielsen, 1952
• Род † Toxoprion Hay, 1909
• Семейство † Edestidae Jaekel, 1899
• Род † Edestus Leidy, 1855
• Род † Helicampodus Branson, 1935
• Род † Listracanthus Newberry & Worthen, 1870
• Род † Lestrodus Obruchev, 1953
• Род † Metaxyacanthus Obruchev, 1964
• Род † Parahelicampodus Nielsen, 1952
• Род † Physonemus Agassiz, 1843
• Род † Prospiraxis Williams, 1887
• Род † Syntomodus Obruchev, 1964
Надсемейство † Caseodontoidea Zangerl, 1981, Род Камподусы — † Campodus Koninck, 1844 (известен из каменноугольного периода): Campodus agassizianus Koninck, 1844; Campodus arizonensis Gass, 1963; Campodus basalis Cope, 1894; Campodus corrugatus Newberry & Worthen, 1870; Campodus scitulus St. John & Worthen, 1875; Campodus variabilis Newberry & Worthen, 1870; Campodus virginianus St. John & Worthen, 1875.
 |
 |
 |
 |
 |
%201f%20fossil.jpg) |
%202f%20fossil.jpg) |
%201f%20fossil.jpg) |
Надсемейство † Caseodontoidea Zangerl, 1981, Род † Chiastodus Trautschold, 1879 состоит из одного вида: Chiastodus obvallatus Trautschold, 1879.
%201f%20teeth.jpg) |
Надсемейство † Caseodontoidea Zangerl, 1981, Семейство † Caseodontidae Zangerl, 1981, Род Казеодусы — † Caseodus Zangerl, 1981 (известен из каменноугольного периода в США (Иллинойс, Индиана, Канзас, Южная Дакота) и раннего триаса из Канады, Британская Колумбия): Caseodus basalis Zangerl, 1981; Caseodus eatoni Zangerl, 1981; Caseodus varidentis Mutter & Neumann, 2008. Представители рода были среднего размера, 1—1,5 м в длину. Caseodus — один из немногих родов Евгенеодонтид, переживших массовое вымирание в конце перми. Это один из последних выживших родов этой клады.
 |
Надсемейство † Caseodontoidea Zangerl, 1981, Семейство † Caseodontidae Zangerl, 1981, Род Эрикодусы — † Erikodus Nielsen, 1952 состоит из одного вида: Erikodus groenlandicus Nielsen, 1932.
%201f%20fossil.jpg) |
%202f%20fossil.jpg) |
%203f%20fossil.jpg) |
Надсемейство † Caseodontoidea Zangerl, 1981, Семейство † Caseodontidae Zangerl, 1981, Род Фадении — † Fadenia Nielsen, 1932 (известен из каменноугольного периода в Миссури (США), из пермского периода в Гренландии, и раннетриасовая эпоха из Гренландии и из формации Серная гора (Sulphur Mountain Formation) в Британской Колумбии, Канада): Fadenia crenulata Nielsen, 1932; Fadenia gigas Eaton, 1962; Fadenia uroclasmato Mutter & Neuman, 2008. Представители рода были среднего размера, до 1,5 м в длину. Нет анального и брюшных плавников, хвостовой плавник полулунный симметричный. Fadenia — один из немногих родов Евгенеодонтид, переживших массовое вымирание в конце перми. Это один из последних выживших родов этой клады.
 |
%201f.jpg) |
Надсемейство † Caseodontoidea Zangerl, 1981, Семейство † Caseodontidae Zangerl, 1981, Род Орнитоприоны — † Ornithoprion Zangerl, 1966 (известен из московского этапа каменноугольного периода — от 315,2 до 307 миллионов лет назад): типовой вид — Ornithoprion hertwigi Zangerl, 1966. У разных видов была удлинённая нижняя челюсть. Открытие орнитоприона помогло установить тот факт, что у геликоприона (Helicoprion) «зубная спираль» была на нижней челюсти.
 |
 |
 |
Надсемейство † Caseodontoidea Zangerl, 1981, Семейство † Caseodontidae Zangerl, 1981, Род Ромеродусы — † Romerodus Zangerl, 1981 состоит из одного вида: Romerodus orodontus Zangerl, 1981.
 |
 |
 |
 |
 |
Надсемейство † Caseodontoidea Zangerl, 1981, Семейство † Eugeneodontidae Zangerl, 1981, Род Боббодусы — † Bobbodus Zangerl, 1981 (известен из каменноугольного и пермского периодов): Bobbodus schaefferi Zangerl, 1981; Bobbodus xerxesi Hampe, Hairapetian, Dorka, Witzmann, Akbari & Korn, 2013.
Bobbodus schaefferi — типовой вид, известный по трём экземплярам, все с восточного побережья Панталласского океана. Они были найдены в касимовских отложениях (от 303,7 до 307,0 миллионов лет назад) из штата Айова, в гжельских отложениях (от 298,9 до 303,7 миллионов лет назад) из штата Небраска, в асселианских отложениях (от 295,0 до 298,9 миллионов лет назад) из Канзаса. У особей этого вида были частичные верхние и нижние зубные ряды, нёбно-квадратный хрящ (palatoquadrate) и жаберные дуги. Bobbodus xerxesi Hampe et al., 2013 известен по единственному зубу, найденному в отложениях Учьяпинги (Wuchiapingian — от 254,14 до 259,1 миллионов лет назад) в горах Багук в центральном Иране. Вид назван в честь Ксеркса, известного правителя археменидов, живших в этой местности, памятники которого произвели впечатление на первого автора, описавшего этот вид.
 |
%201f.jpg) |
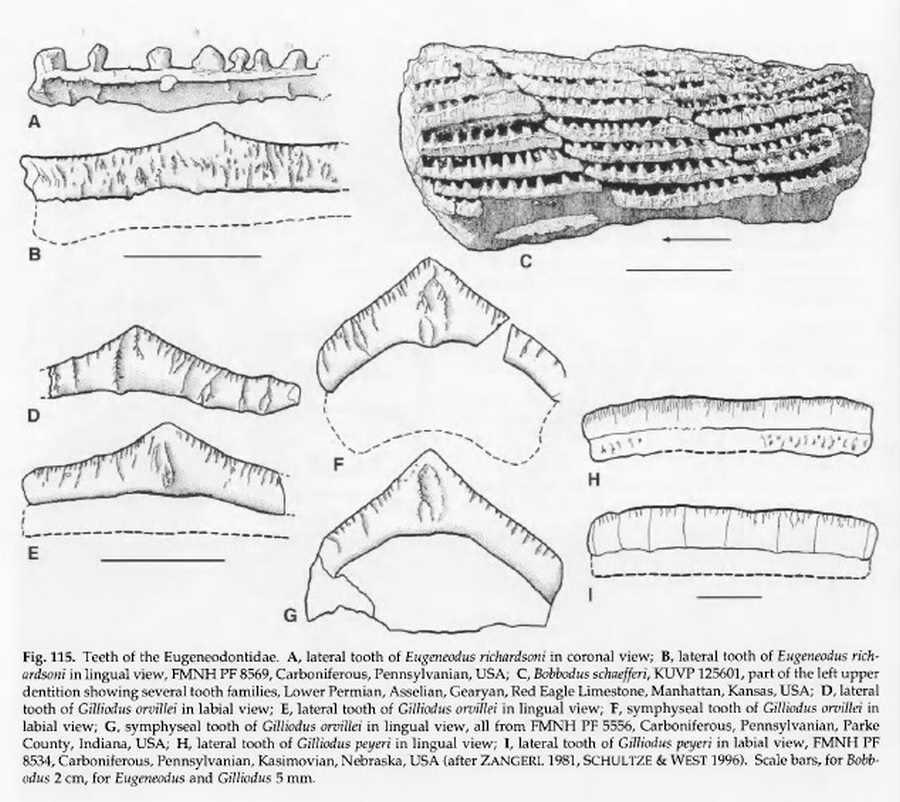
Надсемейство † Caseodontoidea Zangerl, 1981, Семейство † Eugeneodontidae Zangerl, 1981, Род Евгенеодусы — † Eugeneodus Zangerl, 1981 состоит из одного вида: Eugeneodus richardsoni Zangerl, 1981.
%201p.jpg) |
%202p%20teeth.jpg) |
Надсемейство † Caseodontoidea Zangerl, 1981, Семейство † Eugeneodontidae Zangerl, 1981, Род Гиллиодусы — † Gilliodus Zangerl, 1981 состоит из следующих видов: Gilliodus orvellei Zangerl, 1981; Gilliodus peyeri, Zangerl 1981.
%201p%20teeth.jpg) |
 |
Надсемейство † Edestoidea Hay, 1930, Семейство † Helicoprionidae Karpinsky, 1911 (синоним † Agassizodontidae Zangerl, 1981), Род † Agassizodus St. John & Worthen, 1875 (известен из каменноугольного периода): Agassizodus variabilis Newberry & Worthen, 1870.
%204f.jpg) |
 |
%201f%20fossil.jpg) |
%202f%20fossil.jpg) |
%203f%20fossil.jpg) |
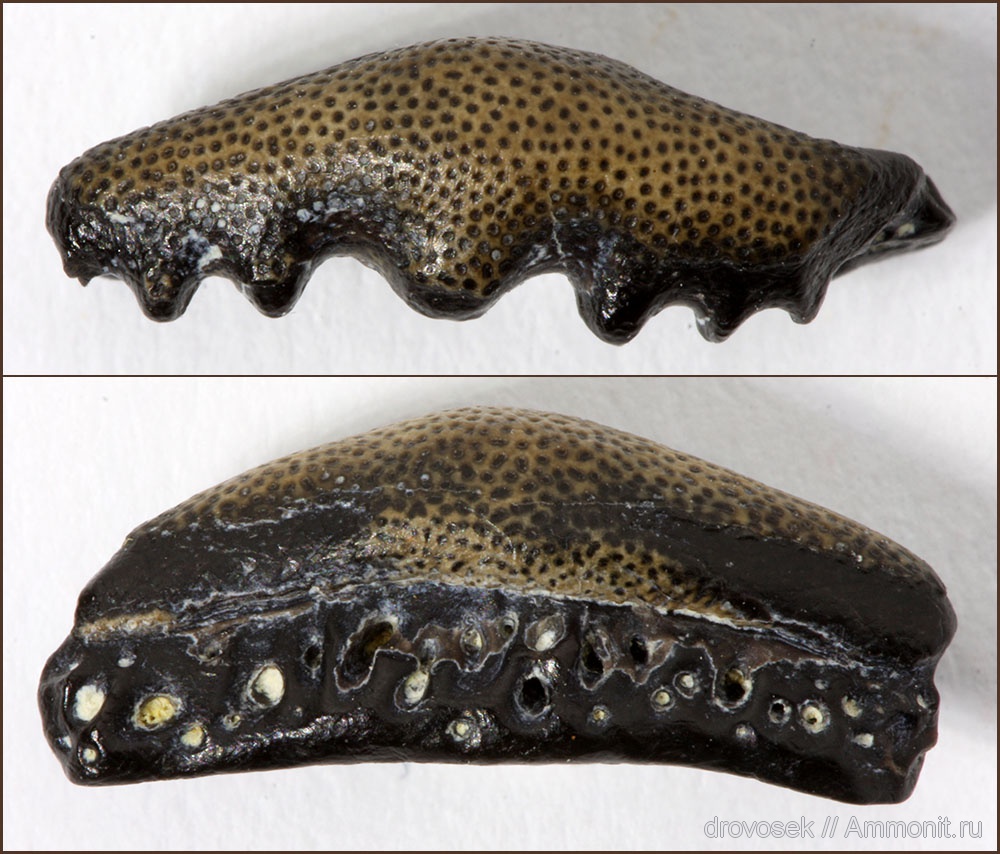
Надсемейство † Edestoidea Hay, 1930, Семейство † Helicoprionidae Karpinsky, 1911 (синоним † Agassizodontidae Zangerl, 1981), Род † Arpagodus St. John & Worthen, 1875 (известен из каменноугольного периода): Arpagodus rectangulus Trautschold, 1879. Только один вид, известен в основном из пенсильванского подпериода (от 298,9 до 323,2 миллионов лет назад), такого как ранний пенсильванский период в России и поздний пенсильванский период в формации известняков Рэйтауна (Raytown Limestone Formation) в Канзасе, США.
 |
 |
Надсемейство † Edestoidea Hay, 1930, Семейство † Helicoprionidae Karpinsky, 1911 (синоним † Agassizodontidae Zangerl, 1981), Род † Campyloprion Eastman, 1902 (известен из каменноугольного периода Северной Америки и Подмосковья): Campyloprion annectans Eastman, 1902; Campyloprion ivanovi Karpinsky, 1922 — Кампилоприон Иванова. Отличается очень высокими зубами (до 10 см и более), образующими, по-видимому, полную спираль.
 |
%201f%20fossil.jpg) |
%202f%20fossil.jpg) |
Надсемейство † Edestoidea Hay, 1930, Семейство † Helicoprionidae Karpinsky, 1911 (синоним † Agassizodontidae Zangerl, 1981), Род † Helicoprion Karpinsky, 1899 (известен из каменноугольного и пермского периодов): Helicoprion bessonowi Karpinsky, 1899; Helicoprion davisii Woodward, 1886; Helicoprion ergassaminon Bendix-Almgreen, 1966; Helicoprion ferrieri, Hay 1907; Helicoprion karpinskii Obruchev, 1953; Helicoprion mexicanus Mullerried, 1945; Helicoprion nevadensis Wheeler, 1939; Helicoprion sierrensis Wheeler, 1939; Helicoprion svalis Siedlecki, 1970. Ближайшими из ныне живущих родственников Helicoprion (и других eugeneodontids) являются химеры.
Впервые описан А. П. Карпинским в 1899 году. Основой для описания послужила так называемая «зубная спираль», найденная в Пермской губернии в 1897 году краеведом А. Г. Бессоновым. На основании строения зубов Карпинский отнёс найденное образование к эдестидам, акулоподобным (или химероподобным) палеозойским рыбам.
Найденная «зубная спираль» оказалась загадочным образованием. А. П. Карпинский предположил, что спираль располагалась на носу рыбы. Это мнение сохранялось в литературе до 1960-х годов. Предлагались и альтернативные гипотезы — например, спираль могла находиться на переднем спинном или на хвостовом плавнике. Существовало предположение, что спираль могла быть подвижной (схожей с циркулярной пилой). Геликоприон был представлен со спиралью на нижней челюсти в руководстве по палеонтологии К. А. Циттеля (1902), где спираль геликоприона названа симфизной. В 1930-х годах были описаны остатки челюстей близкородственной эдестиды саркоприона. У этой рыбы из пермских отложений Гренландии зубная спираль находилась на симфизе нижней челюсти, в специальной полости.
Полные остатки геликоприона пока не найдены. Не вполне ясно, как именно росла спираль. Первоначально считалось, что самые крупные зубы — самые новые и, соответственно, располагались ближе к месту крепления «спирали» на челюсти (нижняя челюсть могла быть загнута вниз). Академик Л. П. Татаринов считал это маловероятным — скорее всего, зубы росли от центра спирали, и самые мелкие зубы — самые молодые. Это подтверждается направлением «шпор» на зубах — у всех родственных видов они направлены назад. По мере роста более крупные старые зубы выталкивались вперёд и сбрасывались. Спираль была «упакована» в симфизную полость и снаружи были видны лишь крупные «старые» зубы. Верхняя челюсть имела обычное строение, ростр мог быть довольно длинным.
В 2008 году была опубликована гипотеза американского палеонтолога Р. Пруди о размещении спирали в районе глотки. Он предположил, что спираль представляет собой глоточные зубы, при этом снаружи она была не видна. Он обосновывает своё предположение отсутствием следов износа на зубах, а также соображениями гидродинамики.
Зубная спираль геликоприона (Helicoprion bessonowi) — одна из самых загадочных окаменелостей пермского периода. Она представляет собой серию острых зубов-лезвий, имеющих общий спирально-свернутый корень с 80—190 зубами. Строение тела остаётся неизвестным.
У близкого рода фадения († Fadenia Nielsen, 1932) нет анального и брюшных плавников, хвостовой плавник полулунный симметричный. Интересно, что у многих евгенеодонтов (а к этой группе относят и эдестид, и геликоприонид) помимо зубной спирали на симфизе есть и ряды давящих зубов на челюстях. Такое сочетание формы тела пелагической рыбы и давящих зубов кажется весьма странным. Поэтому предположение, что геликоприон «выпахивал» моллюсков из морского дна может быть ложным. Не исключено, что геликоприон и родственные ему формы питались аммонитами. В таком случае спираль позволяла, например, обрезать щупальца, а челюстные зубы — раздавить раковины. Но, возможно, геликоприоны питались рыбой (спираль наносила жертве серьёзные раны, а остальные зубы позволяли съесть толсточешуйную добычу). Спирали на зубном симфизе известны у акантод и саркоптеригий, но ни у одной другой рыбы спираль не является столь развитой (2—3 оборота). Современные рыбы не дают нам никакого примера подобного зубного оснащения. Диаметр спирали типового вида геликоприона достигал 25 см. Это предполагает длину всей рыбы около 2—3 метров. В литературе описаны спирали до 90 см в диаметре. Длина их владельца могла доходить до 9—12 метров.
Типовой вид — Helicoprion bessonowi из ранней перми (артинского яруса) Приуралья. Описаны 6—7 видов, из ранней — поздней перми Европы, Азии, Австралии и Северной Америки. В семейство Agassizodontidae (Helicoprionidae) относят примерно 6 родов, распространённых от раннего карбона до поздней перми всех материков. Эти рыбы — не акулы в современном смысле слова, а представители примитивных хрящевых, близких к предкам химер и акул.
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
%201f.jpg) |
%202f.jpg) |
%203f%20fossil.jpg) |
%201f.jpg) |
%201f.jpg) |
Надсемейство † Edestoidea Hay, 1930, Семейство † Helicoprionidae Karpinsky, 1911 (синоним † Agassizodontidae Zangerl, 1981), Род † Parahelicoprion Karpinsky, 1924 (известен из пермского периода из Уральских гор и формации Копакабана (Copacabana Formation), Боливия): Parahelicoprion clerci (Karpinsky, 1916); Parahelicoprion mariosuarezi Merino-Rodo & Janvier, 1986. Название рода, от греческого «почти свернутая пила», напрямую относится к Helicoprion, родственному голоцефалиду, имеющему схожие черты, включая спиралевидный зубной оборот.
Одним из основных качеств, которые отделяют Parahelicoprion от геликоприон является форма, толщина и угол «зубной спирали». Его зубы выступают наружу не как у плотно свернутой пилы. Вместо этого изогнутое расположение режущих лезвий указывает на то, что парагеликоприон меньше полагался на дробление медленно движущихся беспозвоночных и ловлю головоногих моллюсков или других мелких моллюсков, а наносил травматические повреждения более прочной и быстрой добыче. Их зубы росли гораздо медленнее, чем у других подобных акул, в результате чего спираль обесценилась, и вырастало только половина зубов, по сравнению с количеством зубов, которые у геликоприонов вырастает за всю их жизнь. Зубная спираль также могла указывать на возраст рассматриваемого животного.
По оценкам, парагеликоприон вырастал до 12 м в длину. Когда-то считалось, что это самая большая рыба семейства. Однако в 2013 г. была описана «зубная спираль» большого геликоприона, что указывает на то, что представители этого рода могут вырасти до более крупных размеров. Окаменелости парагеликоприона указывают на то, что животное было в целом более стройным и менее тяжелым, чем геликоприоны, несмотря на сопоставимые размеры. Считается, что парагеликоприон был нектоническим хищником, который, вероятно, охотился на множество различных видов, используя свои похожие на лезвия зубы, чтобы разрезать обнажённую плоть, как топор или клин.
 |
 |
%201f.jpg) |
%202f.jpg) |
%203f%20fossil.jpg) |
%201f.jpg) |
Надсемейство † Edestoidea Hay, 1930, Семейство † Helicoprionidae Karpinsky, 1911 (синоним † Agassizodontidae Zangerl, 1981), Род † Sarcoprion Nielsen, 1952 (известен из пермского периода морского горизонта Учьяпинги (Wuchiapingian — от 254,14 до 259,1 миллионов лет назад) в Гренландии): Sarcoprion edax Nielsen, 1952. Название рода, от древнегреческого — «пила для плоти».
Подобно другим евгенеодонтидам, таким как Edestus и Helicoprion, саркоприон был наиболее известен своей чрезвычайно причудливой морфологией зубов по сравнению с другими видами акул и их ближайшими родственниками, химерами. По сравнению с другими представителями Helicoprionidae, его «зубные спирали» оказались более острыми, более компактными и в лучшем состоянии, чем у других акул того времени, и воздерживались от роста до чрезвычайно громоздких форм, которые могли бы вызвать вопросы о его способности правильно питаться. В род входит один вид, Sarcoprion edax («прожорливая мясорубка»), обнаруженный в морских слоях пермского периода в Медделельсер-ом-Гренланд (Meddelelser om Grønland).
Саркоприон имел такую структуру челюсти и рта, что позволяло ему быть более гидродинамичным, уменьшая размер и форму «зубной спирали» и увеличивая размер рострума. Считается, что саркоприон похож на сегодняшнюю акулу мако и преследовал более мелкую, быстро движущуюся добычу. По оценкам, его размер составлял в среднем 6 м по двум экземплярам, обнаруженными в Гренландии. Используя компактный зубчатый венец во время охоты, саркоприон охотился на большое количество видов, ныряя на них с большой скоростью и распиливая уязвимые места жертв. Любые существа, застрявшие между его рострумом и зубами, подвергались ударам по вертикали, получая максимальный урон.
%201f.jpg) |
%202f.jpg) |
%203f.jpg) |
Надсемейство † Edestoidea Hay, 1930, Семейство † Helicoprionidae Karpinsky, 1911 (синоним † Agassizodontidae Zangerl, 1981), Род † Toxoprion Hay, 1909 (окаменелости обнаружены в морских слоях с раннего карбона до поздней перми): Toxoprion lecontei Hay, 1909 — один вид. Название рода, с древнегреческого означает «лучковая пила».
Токсоприон был одним из многих евгенеодонтов каменноугольного периода, у которых был нёбно-квадратный хрящ (palatoquadrate), слитый с его черепом или уменьшенный в других формах, а его сильно зазубренные зубы росли наружу на симфизе нижней челюсти, как у круглой пилы. Несмотря на челюсти, показывающие сходство с другими евгенеодонтами (геликоприон), зубы токсоприонов не растут в «завиток», в котором более мелкие и более ранние зубы перекрыты большими зубами, выращенными позже в жизни, когда челюсть напоминает раковину аммонита. Как и в случае с другими геликоприонидами, токсоприоны вместо этого добавили зубы к конечному концу челюсти, и они выросли в виде нисходящей дуги, а не вокруг предыдущих зубов.
Род получил свое название от греческого слова τοξον (toxon) — «лук», «стрела», «арка», «дуга», из-за дугообразной челюсти, на которой были расположены зубы этих существ, и πριονι (prioni) — «пила», «ножовка» из-за количества треугольных зубов, которые у них были. Первоначально предполагалось, что это представитель рода Helicoprion, но позже было показано, что это отдельное животное.
 |
 |
 |
Надсемейство † Edestoidea Hay, 1930, Семейство † Edestidae Jaekel, 1899, Род † Edestus Leidy, 1855 (окаменелости обнаружены в морских слоях с позднего девона до позднего карбона и ранней перми): Edestus crenulatus Hay, 1909; Edestus giganteus Newberry, 1888; Edestus heinrichi Newberry & Worthen, 1870; Edestus karpinskii Missuna, 1908; Edestus minisculus Hay, 1909; Edestus minor Newberry, 1866; Edestus mirus Hay, 1912; Edestus newtoni Woodward, 1916; Edestus serratus Hay, 1909; Edestus triserratus Newton, 1904; Edestus vorax Leidy, 1855.
Название рода Edestus образованно от греческого слова edeste (пожирать), в связи с ненормальным строением и размером зубов этого вида. Род известен в основном по находкам зубов и челюстей. Представители имели две зубные «дуги», возможно, росшие на симфизах верхней и нижней челюстях. Вероятно, дуги торчали далеко вперёд из пасти. Самые старые зубы находились у конца дуги, молодые — у её основания. В отличие от геликоприона, зубная «спираль» не образовывалась. Коронки зубов на дугах расширены у основания, так называемые «шпоры» направлены назад. Истинное положение зубных дуг достоверно не известно, есть мнение, что это глоточные зубы.
Судя по всему, рыба выглядела так, словно на морде у неё растут зазубренные ножницы. Назначение столь неуклюжего зубного аппарата остается совершенно непонятным. Интересно, что боковые («обычные») зубы часто описывают как давящие, хотя их могло вообще не быть. Неясно, могли ли дуги плотно смыкаться. Предполагается, что эдестус был хищником. Он даже сравнивается с современными ламновыми акулами (тахипелагический охотник за крупной добычей). Если так, то это самая крупная хищная рыба своего времени. Судя по остаткам вида Edestus giganteus (поздний карбона Северной Америки — Оклахома), длина эдестуса могла доходить до 6 метров. Высота зубных коронок этого вида доходит до 8 см. Всего описано примерно 11 видов, из среднего карбона — ранней перми Европы (в том числе России) и Северной Америки. Синонимы рода — Edestodus, Protospirata.
Как и другие родственники эдестуса, такие как Helicoprion , и в отличие от современных акул, у видов Edestus зубы росли в изогнутых «дугах», и они не теряли зубы по мере их износа. В случае эдестуса в каждой челюсти имелся только один ряд зубов, поэтому рот напоминал чудовищную пару зазубренных ножниц. Степень кривизны «дуги» зубов, а также её размеры различны у каждого вида.
Поскольку зубы острые и зазубренные, предполагается, что все виды были плотоядными. То, как они ловили или поедали свою жертву, а также их внешний вид, остаётся чистым предположением до тех пор, пока не будет найдено более полное ископаемое, или череп. Одна из таких теорий относительно того, как эдестус мог охотиться и убивать свою добычу, была обнародована Уэйном М. Итано из Музея естественной истории Университета Колорадо (Natural History Museum of the University of Colorado). Итано исследовал образцы Edestus minor из позднекаменноугольных отложений Техаса и обнаружил закономерности износа, которые предполагают, что Edestus мог охотиться, используя свой причудливый набор зубов, чтобы вертикально травмировать свою добычу, создавая режущие раны и выводя её из строя, а затем проглатывать. Такой метод хищничества оказался бы уникальным и пока ещё неизвестным где-либо в животном мире, если бы он был правильным. Изучение следов износа на образцах в исследовании Итано, также позволяет предположить, что эдестус мог охотиться на животных с жесткой кожей, хотя на каких из них остаётся неизвестными.
Edestus giganteus (также известный как «акула с ножницами») обитал в океанах в течение позднего карбона (306–299 миллионов лет назад). Мало что известно об Edestus giganteus, за исключением одного набора зубов, который в настоящее время хранится в Американском музее естественной истории (American Museum of Natural History) в Нью-Йорке. Палеонтологические исследования показывают, что Edestus giganteus, в отличие от современных акул, не терял изношенных или сломанных зубов. Скорее, он продолжал выращивать новые зубы и десны около задней части рта, в конечном итоге выталкивая старые зубы и десны вперед, пока они не выступили изо рта. Какую функцию выполняли странные зубы, не ясно. Edestus giganteus по некоторым оценкам, вырастал примерно до размеров современной большой белой акулы, что, вероятно, сделало этот вид одним из главных морских хищников своего времени. Как и в случае со всеми другими представителями этого рода, неясно, как Edestus giganteus мог поймать или съесть свою добычу, хотя исследование Edestus minor, опубликованное в 2015 году Уэйном М. Итано, предполагает, что Edestus giganteus мог охотиться и убивать свою добычу вертикальными, сокрушительными ударами своих «ножниц», разрубая и калеча свою добычу.
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
%201f.jpg) |
%202f.jpg) |
%201f%20fossil.jpg) |
%201f%20holotype.jpg) |
%201p.jpg) |
Надсемейство † Edestoidea Hay, 1930, Семейство † Edestidae Jaekel, 1899, Род Геликамподусы — † Helicampodus Branson, 1935 состоит из следующих видов: Helicampodus egloni Obruchev, 1965; Helicampodus kokeni Branson, 1935; Helicampodus qomolangma Zhang, 1976.
%201f%20fossil.jpg) |
%202f%20fossil.jpg) |
%201f%20fossil.jpg) |
%201f%20(Helicampodus%20kokeni).jpg) |
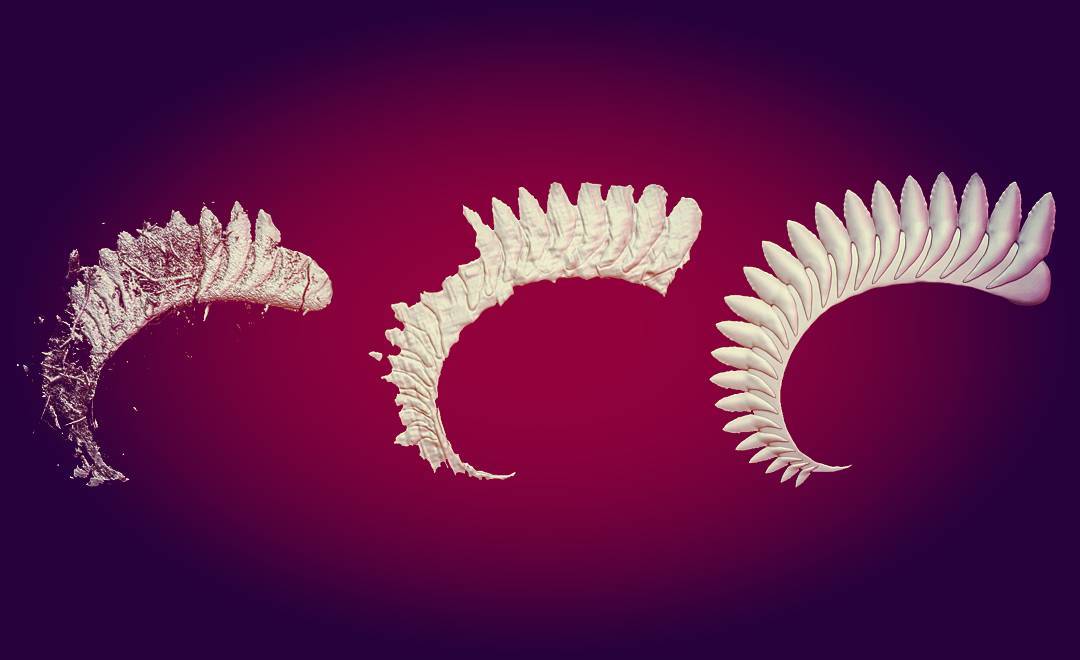
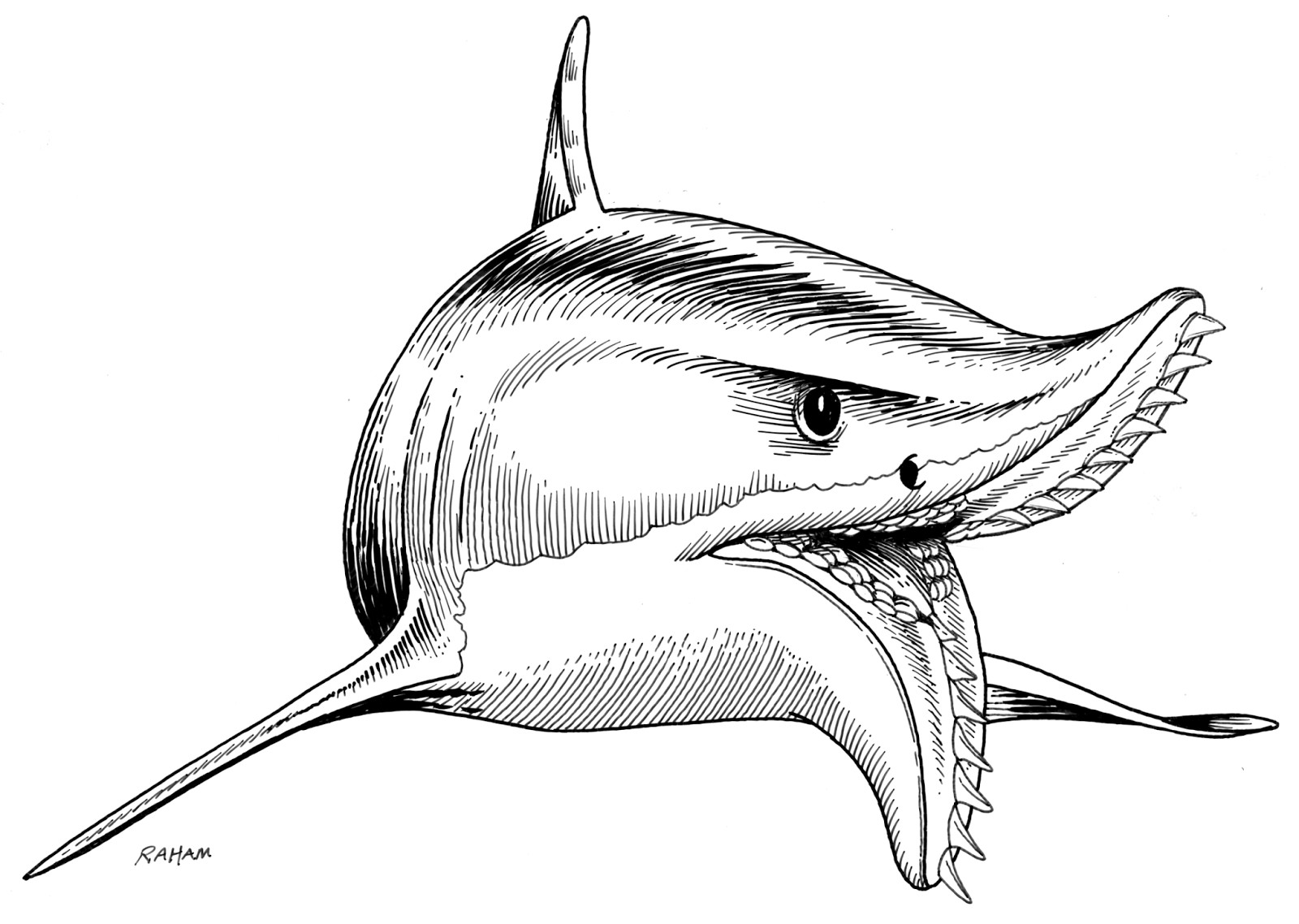
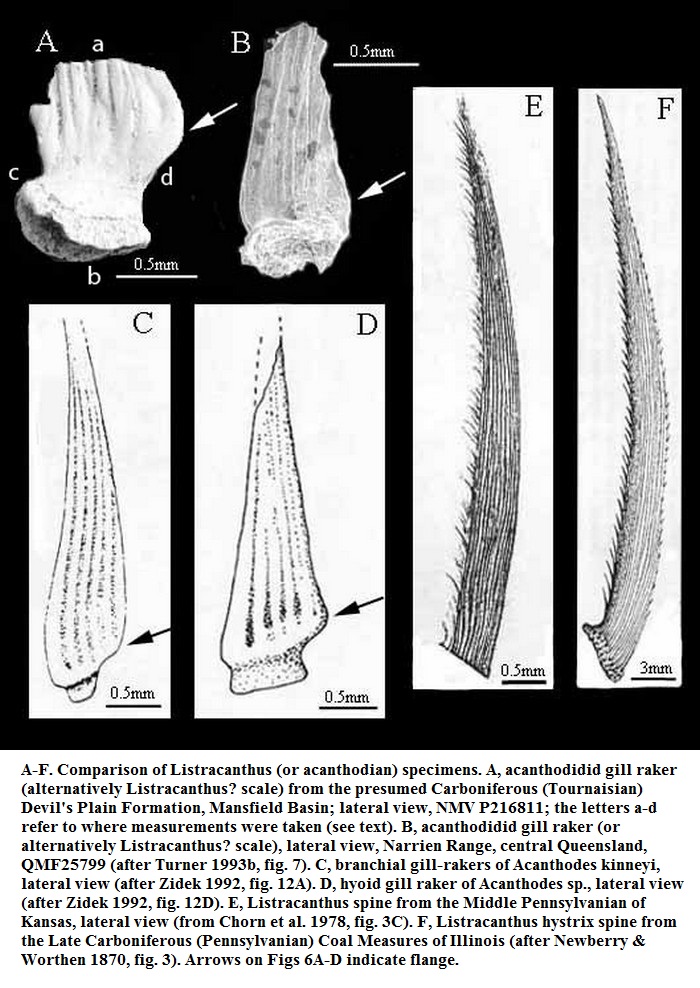
Надсемейство † Edestoidea Hay, 1930, Семейство † Edestidae Jaekel, 1899, Род Листракантусы — † Listracanthus Newberry & Worthen, 1870 (живший с карбона по нижний триас — 326,4—247,2 млн лет назад): типовой вид — Listracanthus hystrix Newberry & Worthen, 1870 (синонимы: Listracanthus beyrichi von Koenen, 1879; Listracanthus eliasi Hibbard, 1938; Listracanthus hildrethi Newberry, 1875; Listracanthus woltersi Schmidt, 1949); Listracanthus pectenatus Mutter & Neuman, 2006. Ещё 2 ранее описанных биномена, относившиеся к роду: Listracanthus spinatus Bolton, 1896 и Listracanthus wardi Woodward, 1891, синонимизированы и перенесены в род Acanthorhachis.
Окаменелости листракантов найдены в Англии, на Южном Урале (Россия), штатах Канзас и Южная Дакота (США), а также в провинции Британская Колумбия (Канада). Окаменелости хрящевых рыб редко когда сохраняются хорошо, чаще всего попадаются их зубы. Листраканты же известны в основном по отпечаткам длинного шипа на голове и спинного плавника, чья высота составляла 10 см.
Положение листракантов в классе хрящевых рыб спорно. Тонкий шип на голове сближает их с представителями отряда ктенакантообразных (Ctenacanthiformes Cappetta, 1988) — самого древнего отряда всех хрящевых рыб, однако отмечено сильное сходство и с евгенеодонтообразными, особенно эдестами (Edestus).
В 2014 году Martill, Del Strother и Gallien при выделении нового рода Acanthorhachis Martill et al. 2014 описали новое семейство Listracanthidae Martill et al. 2014, включающее 2 рода (Acanthorhachis Martill et al. 2014 и Listracanthus Newberry & Worthen, 1870) и занимающее в подклассе пластиножаберных неопределённое положение (incertae sedis).
Внешний вид этих акул в значительной степени неизвестен. Однако автор и иллюстратор Рэй Тролль (Ray Troll) упоминает в своей книге «Шаркабет» («Sharkabet») о том, как палеонтолог Райнер Зангерл (Rainer Zangerl) однажды обнаружил большую глиняную плиту, содержащую длинную, похожую на угря рыба, покрытую длинными шипами и шиповидными зубчиками, которая после высыхания рассыпалась в пыль. Таким образом, согласно рассказу Зангерла, Тролль реконструирует Листраканта, как похожего на огромную акулу с огромными щетинистыми оборками.
 |
 |
 |
 |
 |
 |
%201f.jpg) |
%201f.jpg) |
Надсемейство † Edestoidea Hay, 1930, Семейство † Edestidae Jaekel, 1899, Род Лестродусы — † Lestrodus Obruchev, 1953 состоит из одного вида: Lestrodus newtoni (Woodward, 1916).
 |
Надсемейство † Edestoidea Hay, 1930, Семейство † Edestidae Jaekel, 1899, Род Метаксиакантусы — † Metaxyacanthus Obruchev, 1964 состоит из одного вида: Metaxyacanthus rossicus Inostranzeff, 1888.
%201f%20fossil.jpg) |
Надсемейство † Edestoidea Hay, 1930, Семейство † Edestidae Jaekel, 1899, Род Парагеликамподусы — † Parahelicampodus Nielsen, 1952 состоит из одного вида: Parahelicampodus sparcki Nielsen, 1952.
Надсемейство † Edestoidea Hay, 1930, Семейство † Edestidae Jaekel, 1899, Род Физонемусы — † Physonemus Agassiz, 1843 состоит из следующих видов: типовой вид — Physonemus arcuatus McCoy, 1848; Physonemus asper Leidy, 1859; Physonemus carinatus Saint John & Worthen, 1875; Physonemus gemmatus Newberry & Worthen, 1866; Physonemus gigas Newberry & Worthen 1870; Physonemus hamuspiscatorius Eastman, 1903; Physonemus mirabilis St John & Worthen, 1875; Physonemus pandatus Eastman, 1903; Physonemus stellatus Newberry, 1889; Physonemus striatus Moore, 1929; Physonemus subteres Agassiz, 1843.
 |
 |
%201f.jpg) |
%201f%20fossil.jpg) |
Надсемейство † Edestoidea Hay, 1930, Семейство † Edestidae Jaekel, 1899, Род Проспираксисы — † Prospiraxis Williams, 1887 состоит из следующих видов: типовой вид — Prospiraxis major Newberry, 1885; Prospiraxis randallii Newberry, 1885.
Проспираксис (по-гречески «передняя винтовая ось») достигал длины 5 метров. Эти похожие на акул химеры использовали свою недавно обнаруженную нишу для экспериментов со смертоносными и причудливыми типами зубов, используя свою нижнюю челюсть, как нож для резки мяса, чтобы срывать мясо с утонувших трупов и для медленной охоты.
 |
 |
Надсемейство † Edestoidea Hay, 1930, Семейство † Edestidae Jaekel, 1899, Род Синтомодусы — † Syntomodus Obruchev, 1964 состоит из одного вида: Syntomodus abbreviatus Obruchev, 1964.
|