(Serranidae Swainson, 1839) Семейство Серраниды, Серрановые, Каменные окуни, Family Serranidae Swainson, 1839 (Sea basses, Groupers, Fairy basslets) 72 рода 511 видов
Синонимы:
Grammistidae Bleeker, 1857
Материал подготовил Фактистов И. Е., 2020 г. Сайт www.fishbiosystem.ru
Семейство Serranidae впервые описал в 1839 году английский учёный Уильям Свенсон (Уильям Джон Свенсон (Вильям Джон Свайнсон) — William John Swainson, 1789 — 1855, английский орнитолог, малаколог, конхолог, энтомолог и художник, в 1815 году стал членом Линнеевского общества (Linnean Society), с 1816 по 1818 годы путешествовал по Бразилии, привёз коллекцию из более чем 20 000 насекомых, 1 200 растений, рисунков 120 рыб и примерно 760 шкур птиц, этот материал он использовал при написании книг «Zoological Illustrations» (1820—1823) и «Zoological Illustrations» (1832—1833), был членом Королевского общества (Royal Society) и почётным членом Королевского общества Тасмании (Royal Society of Tasmania), является соавтором книг «Fauna Boreali-Americana» (1831), 3 тома «Naturalist's Library» и 11 томов «Cabinet Cyclopedia», описал несколько высших таксонов и 6 новых видов рыб, в его честь названо 9 видов птиц).
Серраниды, Серрановые, Каменные окуни (Serranidae) — семейство лучепёрых рыб из отряда окунеобразных (Perciformes Bleeker, 1859), обитающих в водах тропического, субтропического и умеренного районов Атлантического, Индийского и Тихого океанов и в прилегающих морях, некоторые виды встречаются также в солоноватой и даже в пресной воде. В данном семействе выделяют три подсемейства: Anthiinae Poey, 1861; Epinephelinae Bleeker, 1874 (трибы Epinephelini, Niphonini, Liopropomatini, Diploprioni, Grammistini) и Serraninae Swainson, 1839. Некоторые авторы выделяют из подсемейства Epinephelinae ещё одно подсемейство — Grammistinae Bleeker, 1857. В прошлом, при использовании не кладистических методов, были предложены классификации с выделением до 15 подсемейств. Ряд авторов выделяли в отдельные подсемейства роды Giganthias (относимый к Anthiinae) и Liopropoma (Epinephelinae). Caesioscorpis считается incertae sedis («неопределённого положения» — латинское выражение, указывающее на то, что положение таксона в системе неясно). Это большое семейство насчитывало по состоянию на 2010 год 511 видов, а на 10.01.2019 — 570 видов.
Семейство в его настоящем составе считается монофилетическим на основе следующих апоморфных признаков: имеются три шипа на крышечной кости, наблюдается редукция задних уроневралий (до их полного отсутствия), редукция шпоры на нижнем дополнительном луче хвостового плавника и третьей преуральной радиалии. Состав семейства и обоснование монофилии заимствованы из работ Джонсона (G.D.Johnson) 1983 и 1984 годов.
Подсемейство Anthiinae Poey, 1861 (антиины). По меньшей мере 21 род, всего приблизительно 170 видов, в основном в Индийском и западной части Тихого океана.
Подсемейство Epinephelinae Bleeker, 1874 (эпинефелины). Личинка имеет одну (обычно вторую) или несколько удлиненных колючек переднего спинного плавника; в трибе Epinephelini, кроме того, удлинённая колючка брюшного плавника. Выделено пять триб на основании признаков личинок и взрослых, согласно данным из нескольких публикаций К. Болдуин (С. С. Baldwin), Д. Джонсона (G. D.Johnson) и А. Кендалла (A. W. Kendall, Jr.). Однако другие авторы (Heemstra & Randall, 1993) ограничивают состав подсемейства собственно груперами (здесь как триба Epinephelini), что соответствует классической точке зрения. Виды родов мыльных рыб — «soapfishes» и четыре рода рыб, содержащих в коже токсин — граммистин, отнесены в трибы Diploprionini и Grammistini подсемейства Epinephelinae.
Подсемейство Serraninae Swainson, 1839 (серранины). Примерно 13 родов.
Название семейства Serranidae образовано от имени типового рода Serranus Cuvier, 1816. На латыни serranus является производным от слова serra — «пила», или saw — «пила», «пилить», что относится к зубчатому преоперкулюму (preoperculum — предкрышечная кость) Serranus cabrilla и Serranus scriba (согласно Кювье — Cuvier, 1828).
Не всех серранид называют «groupers» — «груперами», в семейство входят и «sea basses» — «морские бассы», «морские окуни». Название «grouper» обычно дают рыбам одного из двух крупных родов: Epinephelus и Mycteroperca. Кроме того, виды классифицированные в малых родах Anyperodon, Cromileptes, Dermatolepis, Gracila, Saloptia и Triso также называются «groupers» — «груперами». Рыбы рода Plectropomus называют «coralgroupers» — «коралловыми груперами». Все эти роды относятся к подсемейству Epinephelinae Bleeker, 1874 (эпинефелины). Однако некоторые «hamlets» — «гамлеты», «гуазеты», «альфесты» (род Alphestes), «hinds» — «хайнды», «гаррупы» (род Cephalopholis), в «lyretails» — «вариолы», «лирохвостые груперы» (род Variola) и некоторые другие мелкие роды (гониоплектрусы — Gonioplectrus, нифоны — Niphon, кабинцы, креолы — Paranthias) также находятся в этом подсемействе. Иногда виды в других родах серрановых имеют общие имена, связанные со словом «grouper». Тем не менее, само по себе слово «grouper» обычно означает представителя подсемейства Epinephelinae.
Слово «grouper» — «групер» происходит от португальского названия «garoupa» — «гарупа», которое, как предполагается, произошло из коренного южноамериканского языка. В Австралии слово «groper» — «гропер» используется вместо «grouper» — «групер» для некоторых видов, например, для квинслендского групера — Queensland grouper (Epinephelus lanceolatus). В Новой Зеландии название «groper» — «гропер» относится к виду Polyprion oxygeneios, «wreckfish» — «рыба затонувших кораблей», которую маори называют «hapuku» — «хапуку» . На Филиппинах груперов обычно называют «lapu-lapu» — «лапу-лапу» на острове Лусон (Luzon), а на Висайях (Висайские острова — Visayas Islands) и на острове Минданао (Mindanao) они известны как «pugapo» — «пугапо». На Ближнем Востоке груперов называют «hammour» — «хаммур», их широко употребляют в пищу, особенно в регионе Персидского залива.
Виды из триб Grammistini и Diploprionini выделяют из своей кожи слизь, наподобие токсина, называемую граммистином. Когда они находятся в ограниченном пространстве и подвергаются стрессу, слизь образует пену, токсичную для близко находящихся живых организмов, этих рыб часто называют «soapfishes» — «мыльными рыбами». Они были классифицированы либо как собственные семейства, либо внутри подсемейств, хотя в 5-м изданием «Рыб мира» («Fishes of the World») они классифицируются как трибы внутри подсемейства Epinephelinae.
Найденные окаменелости (fossils) ранних серранид были датированы нижним палеоценом. Время появления некоторых современных родов семейства Серрановые [Sepkoski, Jack. A compendium of fossil marine animal genera // Bulletins of American Paleontology — Paleontological Research Institution, 2002. — Vol. 364. — P. 560.]:
• † Centropristis Cuvier, 1829 — в начале эоцена (окаменелые останки найдены в Германии и США);
• † Paralabrax Girard, 1856 — в начале эоцена (эоцен Великобритании, олигоцен Германии);
• † Serranus Cuvier, 1816 — в начале эоцена (эоцен Туркменистана и Великобритании; олигоцен Азербайджана, Чешской Республики, Германии, Румынии, Грузии, Российской Федерации; миоцен Алжира, Словакии; плиоцен Греции; четвертичный период Италии);
• † Nemanthias — в начале олигоцена;
• † Epinephelus Bloch, 1793 — в середине миоцена, 11,608 млн лет назад (миоцен Алжира, Италии, Словакии, США (Калифорния), Венесуэлы; плиоцен США (Северная Каролина); четвертичный период Италии, острова Теркс и Кайкос);
• † Mycteroperca Gill, 1862 — в середине плиоцена, 5,332 млн лет назад (плиоцен США (Северная Каролина), четвертичный период островов Теркс и Кайкос).
Из-за большого разнообразия сложно определить представителей семейства Serranidae по внешним признакам. Хотя большинство серранид внешне похожи на окуня, существует значительное разнообразие форм и повадок. Серрановые рыбы имеют стройное (иногда массивное), вытянутое тело, умеренно сжатое с боков, и покрытое мелкой, обычно грубой, ктеноидной чешуёй (у некоторых видов циклоидная). Спинной плавник обычно сплошной с длинным основанием, иногда с вырезкой, с хорошо развитой колючей частью, в ней 7—13 колючих лучей (большинство видов имеет 10 или меньше, 13 только у Acanthistius и Niphon). Иногда колючая часть представляет собой отдельный плавник. В анальном плавнике три колючих луча. В брюшных плавниках, расположенных на груди, один колючий и 5 мягких лучей. Нет очешуенного аксиллярного отростка у основания брюшного плавника. Хвостовой плавник обычно закруглённый, усечённый или с мелкой округлой вырезкой (редко глубоко раздвоенный). Боковая линия полная и непрерывная (отсутствует у одного вида), не простирается на хвостовой плавник (подходит близко к основанию спинного плавника у некоторых видов из подсемейства Anthiinae). Большой конечный рот, нижняя челюсть выступает за верхнюю. Когда рот закрыт, виден конец верхнечелюстной кости, т. е. он не скрыт под слезной костью. На челюстях более одного ряда острых зубов, большинство из них простые, передние зубы у некоторых видов — клыки (обычно с парой особенно больших, выступающих из нижней челюсти); верхняя и боковые стороны нёба обычно с простыми зубами. Крышечная кость с тремя шипами (верхний и нижний шипы часто незаметны) — главный шип с дополнительными шипами, из которых один длиннее главного шипа, а второй короче. Задний край предкрышечной кости (preopercle) почти всегда зазубрен или с мелкими шипами. Лучей жаберной перепонки — бранхиостегальных лучей (branchiostegal rays) — обычно 7. В позвоночнике обычно 24—26 позвонков (25—28 у Anthiinae и некоторых Grammistinae и 30 у Niphon).
Каменные окуни — гермафродиты, хотя гонады обоих полов обычно не развиваются в одно и то же время (большинство видов рода Serranus и близкородственные им таксоны являются функциональными гермафродитами — способностью к оплодотворению обладают оба типа гамет, как мужские, так и женские). Груперы — протерогинические гермафродиты (репродуктивный цикл начинается с женской особи, а позже пол меняли на мужской). Представители подсемейства Anthiinae меняют пол с самок на несколько доминирующих самцов. Синхронный гермафродитизм (особь способна одновременно продуцировать и мужские, и женские гаметы) примитивен по отношению к протерогиническому гермафродитизму в филогенной эволюции семейства Serranidae.
Многие виды серранид ярко окрашены, что характерно для рифовых рыб. Окраска обычно маскировочная, состоящая из полос и пятен, часто располагающихся в шахматном порядке. Однако, не смотря на приверженность серрановых к коричневым и песчаным тонам, встречаются в их окраске и красный, и даже синий цвета. Их яркие цвета, скорее всего, представляют собой разрушительный камуфляж, похожий на полосы тигра. Груперы достигают максимальной длины до 3 м и веса более 400 кг, хотя многие виды серранид, не вырастают более 10 см, а некоторые из подсемейства Anthiinae, такие как Plectranthias longimanus, достигают только 3 см.
Распространение (карта): океаны и моря тропического, субтропического и умеренного климата. Некоторые виды обитают или заходят в пресную воду. Обычно они встречаются над рифами, в тропических и субтропических водах вдоль побережья, предпочитая каменистые участки. Каменные окуни ведут преимущественно одиночный, территориальный образ жизни.
Все каменные окуни являются хищниками (плотоядны). Распространён также каннибализм. Хотя некоторые виды, особенно в подсемействе Anthiadinae, питаются только зоопланктоном, большинство серранид питаются рыбой и ракообразными. Как правило, они устраивают засаду, прячутся в укрытиях на рифе и быстро нападают, чтобы схватить проходящую добычу.
В своём большинстве — это донные хищники, а некоторые виды высокопроизводительные промысловые рыбы. Серрановых промышляют крючными орудиями лова, бьют острогами или гарпунами при подводной охоте.
Каменные окуни производят большое количество икринок, а их личинки являются планктонными, как правило, находятся во власти океанических течений, пока они не будут готовы поселиться во взрослой популяции.
Некоторые крупные виды серрановых обычно выставляются в общественных аквариумах и зоопарках, более мелкие — являются популярными аквариумными рыбками. Груперы — очень выносливые аквариумные рыбки, но быстро растущие. В отличие от ближайших родственников — груперов, каменные окуни более мирные рыбы. Благодаря не очень крупным размерам для их содержания не требуются чрезмерно большого аквариума. Представители подсемейства Anthiinae — это в основном мелкие красочные планктоноядные животные, питающиеся в основном крошечными ракообразными и икрой рыб. Несмотря на свою привлекательную окраску, им нужен зоопланктон в качестве пищи, поэтому они плохо подходят для домашних аквариумов. Подсемейство Grammistinae получили свое название от горького и ядовитого токсина кожи, граммистина, который может убить других животных в аквариуме.
Подобно другим рыбам, серраниды содержат паразитов, в том числе нематод (тип Нематоды, Круглые черви — Nemata), цестод (класс Цестоды, Ленточные черви — Cestoda), дигенеев (подкласс Дигенетические сосальщики (Digenea) — паразитические плоские черви из класса Трематоды — Trematoda), моногенеев (подкласс Моногенетические сосальщики (Monogenea) — класса Трематоды — Trematoda), равноногих (изоподы) и веслоногих (копеподы, надотряд Copepoda — класса Челюстеногие ракообразные (Maxillopoda) ракообразных. Исследование, проведенное в Новой Каледонии, показало, что у серрановых, связанных с коралловыми рифами, обитает около 10 видов паразитов на каждый вид рыб.
Недавние генетические исследования показывают, что Serranidae следует разделить как минимум на 2 семейства. Семейство Epinephelidae для групперов (groupers), включая Alphestes, Cephalopholis, Epinephelus, Dermatolepis, Hyporthodus, Liopropoma, Mycteroperca, Paranthias, Pseudogramma и Rypticus. Семейство Serranidae для морских окуней (sea basses), включая Centropristis, Diplectrum, Hypoplectrus, Paralabrax, Plectranthias, Schultzea, Serraniculus и Serranus). Возможно, даже отделение Anthiins (антиин) от Serranids (серранид), чтобы сформировать семейство Anthiidae, включая роды Anthias, Bathyanthias, Bullisichthys, Hemanthias, Parasphyraenops, Plectranthias, and Pronotogrammus). Однако эта ситуация не решена, и нет четких устойчивых морфологических различий между взрослыми особями этих групп. Поэтому на данный момент мы продолжаем использовать Serranidae как универсальную группу.
Вымершие представители семейства Серрановые:
• † Allomorone Frizzell & Dante, 1965 — 40,4—23,03 млн лет назад, олигоцен Германии, эоцен США (Алабама, Калифорния, Луизиана, Техас). [D. L. Frizzell and J. H. Dante. 1965. Otoliths of some early Cenozoic fishes of the Gulf Coast. Journal of Paleontology 39(4):687-718]
• † Diplectrum Holbrook, 1855 — 48,6—2.588 млн лет назад, эоцен Франции, миоцен Панамы, плиоцен Ямайки и США (Флорида, Северная Каролина).
• † Palaeopercichthys Greenwood, 1983 — (типовой вид — Palaeopercichthys arabis Greenwood, 1983) — 66,043—55,8 млн лет назад, неполный синкраниум, частичный череп обнаружены только в Джебель-Умм-Химар (Jabal Umm Himar), в палеоценовых лагунных аргиллитах в формации Умм-Химар (Umm Himar Formation), палеоцен Саудовской Аравии.
• † Paleoserranus Cantalice et al., 2018 — (типовой вид — Paleoserranus lakamhae Cantalice et al., 2018) — описан на основе хорошо сохранившихся окаменелостей из палеоценовых морских отложений геологической единицы Тенехапа-Лакандон (Tenejapa-Lacandón), принадлежащих как карьерам Дивизиона дель Норте (Division del Norte), так и Белисарио Домингес (Belisario Domínguez), недалеко от Паленке (Palenque), штат Чьяпас (Chiapas), на юго-востоке Мексики. Эта ископаемая рыба имеет некоторые общие черты с разными видами, входящими в подсемейства Serraninae, Anthiinae и Ephinephelinae. Самая ранняя окаменелость семейства Serranidae. Место появления этой новой летописи окаменелостей предполагает, что морские окуни произошли в Карибском регионе Северной Америки. [K. M. Cantalice, A. M. Martinez-Melo, and V. A. Romero-Mayén. 2019. The paleoichthyofauna housed in the Colección Nacional de Paleontología of Universidad Nacional Autónoma de México. Zoosystematics and Evolution 95(2):429-452]
• † Percostoma Casier, 1966 — (вид — Percostoma angustum Casier, 1966) — 55,8—48,6 млн лет назад, встречается только на острове Шеппи (Isle of Sheppey), в ипрском морском аргиллите в формации Лондонской глины (London Clay Formation), эоцен Соединенного Королевства. [E. Casier. 1966. Faune ichthyologique du London Clay 1-510]
• † Plesioserranus Casier, 1966 — (вид — Plesioserranus wemmeliensis (Storms, 1898)), встречается только на острове Шеппи (Isle of Sheppey), в ипрском морском аргиллите в формации Лондонской глины (London Clay Formation), эоцен Соединенного Королевства. [E. Casier. 1966. Faune ichthyologique du London Clay 1-510]
• † Prolates Priem, 1899 — (вид — Prolates dormaalensis Casier, 1967 — 55,8—50,3 млн лет назад, встречается только на участке Фишер (Fisher) / Салливан (Sullivan), Вирджиния, США), эоцен Соединенного Королевства, США (Вирджиния).
• † Properca Sauvage, 1880 — (вид — Properca sabbae Pauca, 1929) — 33,9—28,4 млн лет назад, встречается только в Сусланештах (Suslanesti), олигоцен Румынии. [M. Pauca. 1929. Vorläufige mitteilung über eine fossile fischfauna aus den Oligozänschiefern von Sulanesti, Muscel. Academia Romana 12(4-5):26-34]
• † Proserranus Patterson, 1964 — вымерший род доисторических костистых рыб, которые жили в данианский период — 66,0—61,6 млн лет назад, эпохи палеоцена.
• † Protanthias David, 1943 — (вид — Protanthias fossilis David, 1943) — 11,608—7,246 млн лет назад, частичный скелет обнаружен в карьере Ломпок (Lompoc Quarry), который находится в тортонских базальных (силикокластических) диатомитах в формации Монтерей (Monterey Formation) в Калифорнии, США. [L. R. David. 1943. Miocene Fishes of Southern California. Geological Society of America Special Paper 43:1-187]
• † Serraninus Pauca, 1931 — (вид — Serraninus moravicus Pauca, 1931 = Oligophus moravicus Pauca, 1931) — 37,2—28,4 млн лет назад. [M. Pauca. 1931. Zwei Fischfaunen aus den oligozaenen Menilitschifern von Mähren. Annalen des Naturhistorischen Museums in Wien 46:147-152]
• † Serranopsis Casier, 1966 — (вид — Serranopsis londinensis Casier 1966) — 55,8—48,6 млн лет назад, встречается только на острове Шеппи (Isle of Sheppey), в ипрском морском аргиллите в формации Лондонской глины (London Clay Formation), ранний эоцен Соединенного Королевства. [E. Casier. 1966. Faune ichthyologique du London Clay 1-510]
• † Tavania Landini & Menesini, 1978 — 5,332—2,588 млн лет назад, обнаружен на территории Италии.
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
Семейство Серрановые (Serranidae)
Отличается коротким анальным плавником, представленным обычно лишь тремя колючками и несколькими ветвистыми лучами. Рот конечный, большой. Чешуя мелкая, циклоидная. Жаберные крышки и «щеки» с двумя типами чешуи. Морские теплолюбивые рыбы.
Декоративное рыбоводство. А.М.Кочетов 1991 г.
Семейство Серрановые (Serranidae)
Каменные окуни для морского аквариума родов Anthias, Cromileptis, Cephalopholis, Epinephelus, Grammistes, Hypoplectrus, Paranthias, Promicrops, Myrolabrichthys и другие; размер 10—60 см. Окраска многоцветная (редко однотонная) с глазчатыми пятнами, светящимися точками и контрастными полосами. Тело умеренно удлиненное, сжато с боков. В Черном море — окунь-зебра (Serranus scriba). Рыбы — хищники. Известны случаи нереста в неволе. Аквариум от 300 л.
Иллюстрированная энциклопедия рыб. Ст.Франк 1983 г.
Рыбы обычно раздельнополы. Однако и тут исключение подтверждает правило. У некоторых костистых рыб двуполость, гермафродитизм - явление обычное; например, у морских окуней семейства Serranidae.
Представители следующего семейства серановые, или каменные окуни (Serranidae) широко распространенные морские рыбы, известные с эпохи эоцена до наших дней. Они встречаются главным образом в тропических и субтропических морях, однако некоторые виды живут и в пресной воде. Сюда относится много родов, причем род Serranus включает в себя более 200 видов, некоторые из них заходят в реки.
Ихтиопланктон Черного моря. Т.В.Дехник 1973 г.
СЕМ. SERRANIDAE — СЕРРАНОВЫЕ
В Черном море встречаются три вида, относящиеся к двум родам — Serranus scriba (Linne), Serranus cabrilla (Linne) и Morone labrax (Linne) (Pacc, 1949, 1965; Световидов, 1964).
Достоверно размножается здесь один вид — Serranus scriba.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Семейство Серрановые, или Каменные окуни (Serranidae)
Представители этого семейства
обычно имеют один спинной плавник
с хорошо развитой колючей частью,
которая иногда бывает обособлена в
отдельный плавник. Мягкая часть
спинного плавника намного длиннее анального.
В анальном плавнике три сильные
колючки. Брюшные плавники расположены на
груди и имеют одну колючку и 5 мягких
лучей. Подкрышечная кость
оканчивается острым зазубренным краем. Мелкие
острые щетинковидные зубы на челюстях
расположены в несколько рядов. У
некоторых видов в передней части верхней
и нижней челюстей имеются клыковидные
зубы.
Семейство объединяет около 75 родов
и более 400 видов преимущественно
морских рыб, обитающих в субтропической
и тропической областях Атлантического,
Тихого и Индийского океанов. Некоторые
серрановые (китайский окунь) обитают
в пресных водах.
Каменные окуни — чрезвычайно
разнообразная группа рыб. Есть виды
длиной не более 20 см и весом 100 г, но в то
же время к этому семейству относятся
настоящие гиганты, превышающие рост
взрослого человека и весящие более
300 кг. Окраска каменных окуней очень
яркая и разнообразная. На общем, чаще
всего темном, фоне имеется большое
количество ярких полос, пятен, точек. Такой
тип окраски, как правило, наблюдается
у рыб, обитающих среди камней, скал или
коралловых рифов. Само название
«каменные окуни» свидетельствует о том, что
серрановые живут в основном в
прибрежных районах на скалистых и коралловых
грунтах, прекрасно маскируясь в ярких
зарослях кораллов, актиний, среди
причудливого нагромождения камней и
подводной тропической растительности.
Все без исключения каменные окуни —
прожорливые хищники, обычно
подстерегающие свою жертву в засаде: расщелине
скалы, среди камней, в зарослях
водорослей или кораллов. Часами может
поджидать каменный окунь свою добычу, не
выходя из укрытия; защитная окраска
делает его почти неотличимым от
окружающих предметов. Только ритмичное
движение жаберных крышек да слабые,
едва заметные повороты золотистых глаз
могут выдать присутствие затаившейся
рыбы. Но кажущееся спокойствие
обманчиво: стоит появиться вблизи небольшой
рыбке, как стремительным броском окунь
вырывается из засады. Движение
челюстей, и... только серебристые чешуйки,
медленно опускающиеся на дно, остались
от неосторожной жертвы. Хищник же
снова занимает свою позицию среди
камней. Каменные окуни рано переходят на
хищный образ жизни; молодые рыбки
длиной 2—3 см питаются уже
преимущественно молодью рыб, при этом нередко
наблюдаются случаи каннибализма,
который отмечается и у взрослых особей.
Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Часть 3. (Perciformes). Г.У.Линдберг, З.В.Красюкова 1969 г.
ХС. Сем. SERRANIDAE — КАМЕННЫЕ ОКУНИ, СЕРРАНОВЫЕ
Тело продолговатое, сжатое с боков. Рот выдвижной, большой или умеренной величины, обычно не очень косой. Верхнечелюстные кости сзади расширены, почти всегда свободны, редко скрыты под предглазничной костью, которая, как правило, узкая. Верхнечелюстные кости иногда имеют дополнительную косточку. С каждой стороны рыла по 2 ноздри. Зубы на челюстях, сошнике и нёбных конические или заостренные; на языке и крыловидных костях зубы встречаются реже. Крышечная кость имеет от 1 до 3 колючек. Предкрышечная кость обычно более или менее зазубрена по краю, реже гладкая. Жаберные тычинки длинные или короткие, почти всегда жесткие и вооружены зубчиками. 4 жабры, за четвертой — длинная щель. Ложножабры имеются, большие. Жаберные перепонки разделены, свободны от межжаберного промежутка. Колючая часть спинного плавника хорошо развита, с 6—15 (По сравнению с Катаямой (Katayama, 1960 : 3) исправлено 6 вместо 2, так как 2—3 колючих луча в спинном плавнике встречаются только в подсем. Grammistiinae, которое мы рассматриваем как самостоятельное семейство, так же как и подсемейства Acropominae и Ostracoberycinae, а род Synagrops включаем в сем. Apogonidae.) колючими лучами; мягкая его часть с 10—30 лучами. Анальный плавник короткий, с 3 колючими лучами, реже с 2 (Diploprion); мягких лучей от 7 до 12. Брюшные плавники расположены на груди, обычно с 1 колючим и 5 мягкими лучами; отчетливо выраженной аксиллярной пластинки нет. Грудные плавники хорошо развиты, с узким основанием. В хвостовом плавнике 15—17 основных лучей, из которых 13—15 ветвистые. Чешуя умеренной величины или мелкая, циклоидная или чаще ктеноидная, иногда погружена в кожу. Боковая линия, как правило, полная и одна, не заходит на хвостовой плавник. Вторая подглазничная косточка имеет подглазничный выступ, поддерживающий глазное яблоко. Череп без шипов и обычно без хорошо развитой пещеристой структуры. Позвонков 24, из них 14 хвостовых, иногда число позвонков бывает большим, но никогда не превышает 37. Все или большинство ребер прикреплены к парапофизам, когда они развиты; передние позвонки без парапофизов. Плавательный пузырь имеется, почти всегда мал и прикреплен к стенке полости тела. Пилорических придатков много или мало; кишечник короткий.
Хищные, главным образом морские рыбы тропических и умеренных морей, иногда живут в пресных и солоноватых водах. Большей частью являются ценным пищевым объектом промысла (Katayama, 1960 : 3).
Много родов и видов. В Японском море 17 родов и 32 вида, 2 рода и 4 вида в сопредельных водах.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ СЕМ. SERRANIDAE
(Таблица составлена по работе Катаямы (Katayama, 1960 : 7), с изменениями применительно лишь к Японскому морю.)
1 (2). Боковая линия проходит под самым основанием спинного плавника и по верху хвостового стебля, где она может и отсутствовать. 6 лучей жаберной перепонки. Края предкрышки гладкие. 1. Callanthias Lowe.
2 (1). Боковая линия проходит в значительном отдалении от основания спинного плавника по боку тела, где она образует дугу, и по средней линии хвостового стебля. 7 лучей жаберной перепонки. Края предкрышки обычно зазубрены.
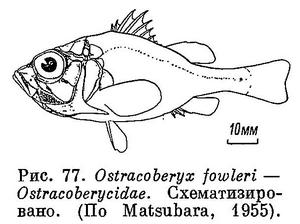
3 (10). Крышечная кость с 2 шипами или выступами, иногда с 1. Крышечная кость без шипов, направленных назад, а предкрышечная — с длинным сильным выростом, направленным назад и заходящим за основание грудного плавника (рис. 77), характерны для рода Ostracoberyx, распространенного вблизи Занзибара, у Мальдивских и Филиппинских островов и у тихоокеанских берегов Японии.

4 (9). Спинных плавников 2, если плавники соприкасаются своими основаниями, то в таком случае последний колючий луч много больше предпоследнего, а основание мягкой части довольно длинное, около 1.5 раз в длине основания колючей части (рис. 115).
5 (8). Чешуя довольно крупная (l.l. 28—52), легко опадающая. Позвонков 25. Перитонеум черный.
6 (7). Впереди на челюстях 1—2 пары клыков и несколько клыков по бокам нижней челюсти. На симфизисе нижней челюсти нет пары колючих зубовидных шипов, направленных книзу и вперед. 2. Doederleinia Steindachner.
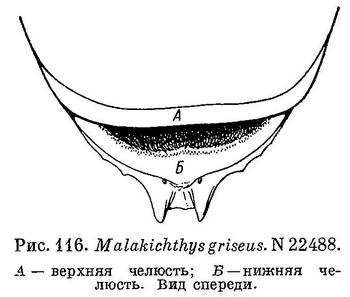
7 (6). Впереди на челюстях и по бокам челюстей нет клыков. На симфизисе нижней челюсти имеется пара колючих зубовидных шипов, направленных вниз и вперед (рис. 116). 3. Malakichthys Doderlein.

8 (5). Чешуя довольно мелкая (в боковой линии прободенных чешуй 71—86), ктеноидная, плотно сидящая. Позвонков 34—37. Перитонеум белый. 4. Lateolabrax Bleeker.
9 (4). Спинной плавник 1, он глубоко выемчатый, последний колючий луч примерно равен предпоследнему, а если превышает его, то основание мягкой части снинного плавника короткое — около 2 раз в основании колючей части плавника (рис. 117). 5. Stereolepis Ayres.
10 (3). Крышечная кость с 3 шипами или выступами.
11 (12). Спинных плавников 2, если они соприкасаются, то последний колючий луч в несколько раз больше предыдущего колючего луча. Чешуя мелкая; 140—150 поперечных рядов чешуй; прободенных
чешуй 84—93. 6. Niphon Cuvier.
12 (11). Спинной плавник 1, иногда с более или менее глубокой выемкой, но чаще без нее.
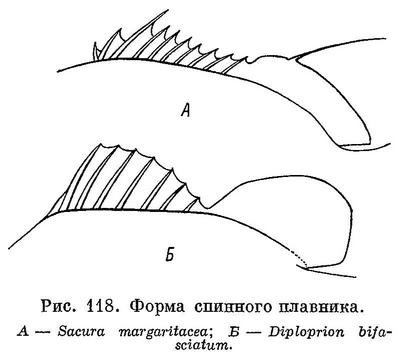
13 (26). Первые ветвистые лучи спинного плавника в 1.5—3, а то и более (до 8) раз длиннее последнего колючего луча (рис. 118). Выемка в плавнике выражена достаточно отчетливо.
14 (17). Верхушка черепа и поверхность костей на боках головы шершавые. Чешуя мелкая, в боковой линии 80—87 прободенных чешуй.
15 (16). Тело укороченное и высокое (его высота содержится 2.1—2.3 раза в длине тела без С). Спинной плавник глубоко выемчатый, его колючая часть высокая. D VIII 13—15; А II 12. На рисунке у Катаямы (Katayama, 1960 : 42, pl. 6) в анальном плавнике ошибочно изображено 3 колючих луча, а в тексте указано 2, что правильно.7. Diploprion Kuhl et Van Hasselt.
16 (15). Тело удлиненное (высота его 2.6—3.5 раза в длине тела без С). Спинной плавник с умеренной выемкой; его колючая часть не очень высокая. D IX 12; А III 9. 8. [Aulacocephalus Temminck et Schlegel].
17 (14). Верхушка черепа и поверхность костей на боках головы не шершавые. Чешуя относительно крупная, в боковой линии 26—51 прободенных чешуй.
18 (21). Чешуя сравнительно мелкая, l.l. 42—51. D VIII—X 9—12.
19 (20). Верхнечелюстная кость покрыта чешуей. Боковая линия с крутым изгибом в передней части, l.l. 50—52. D VIII 13—14; А III 9—10. 9. Chorististium Gill.
20 (19). Верхнечелюстная кость голая, не покрыта чешуей. Боковая линия в виде очень пологой кривой, l.l. 42—45. D IX—X 9—10; А III 6. 10. Chelidoperca Boulenger.
21 (18). Чешуя сравнительно крупная, l.l. 26—36. D X 14—18.
22 (25). Хвостовой плавник умеренного размера, заметно короче длины головы. Третий колючий луч спинного плавника не удлинен по сравнению с соседними.
23 (24). Впереди на нижней челюсти нет клыков. Спинной плавник глубоко выемчатый. Хвостовой плавник слегка закруглен или обрезан и не имеет удлиненных лучей. Третий луч мягкого спинного плавника не удлинен. 11. Sayonara Jordan et Seale.
24 (23). Впереди на нижней челюсти имеются клыки. Спинной плавник значительно вырезан, но меньше, чем у Sayonara. Хвостовой плавник обрезан; его верхний крайний луч удлинен. Третий луч мягкого спинного плавника также удлинен. 12. Zalanthias Jordan et Thompson.
25 (22). Хвостовой плавник увеличен: у самок он равен длине головы, у самцов — в 2 раза длиннее; верхний и нижний лучи плавника сильно удлинены и придают плавнику вильчатую форму. Третий колючий луч спинного плавника заметно удлинен по сравнению с соседними; третий луч мягкой части спинного плавника также удлинен. 13. Sacura Jordan et Thompson.
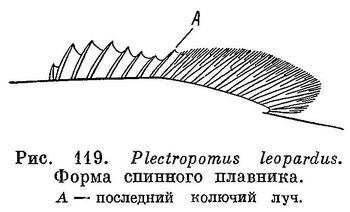
26 (13). Первый ветвистый луч спинного плавника примерно равен последнему колючему лучу; выемка в плавнике не выражена или выражена очень слабо (рис. 119).
27 (32). Хвостовой плавник закруглен или обрезан или даже слегка вогнут (Plectropomus). Подглазничный выступ развит только на второй подглазничной косточке.
28 (29). В боковой линии более 80 прободенных чешуй (l.l. 81—83). D VI—VIII 11—12. Тело и плавники густо покрыты очень мелкими черными пятнышками (около 25 косых рядов от начала спинного плавника до начала анального). Хвостовой плавник обрезан или слегка вогнут. 14. Plectropomus Oken.
29 (28). В боковой линии менее 80 прободенных чешуй (l.l. 47—71). D IX 12—21. Тело и плавники не имеют такой окраски.
30 (31). Мягких лучей в спинном плавнике 12—18, в анальном — 7—8. У Epinephelus septemfasciatus в анальном плавнике 9—10 мягких лучей. 15. Epinephelus Bloch.
31 (30). Мягких лучей в спинном плавнике 19—21, в анальном — 10. 16. Trisotropis Gill.
32 (27). Хвостовой плавник с выемкой, иногда очень большой. Подглазничный выступ развит на второй и третьей подглазничных костях, а иногда на четвертой и первой.
33 (34). Грудной плавник заострен; его верхние лучи значительно длиннее средних и нижних. Ноздри заметно разобщены. D VIII—IX. Выемка хвостового плавника слабая. 9. Chorististium Gill.
34 (33). Грудной плавник закруглен или симметричен. Ноздри сближены. D X.
35 (36). Зубы на сошнике в виде большого ромбовидного пучка. По бокам нижней челюсти, в задней части, зубы образуют бархатистые скопления. Выемка хвостового плавника слабая. В боковой линии 57—61 прободенная чешуя. Мягких лучей спинного плавника 19—21. 17. Caprodon Temminck et Schlegel.
36 (37). Зубы на сошнике в виде маленького треугольного пучка. По бокам нижней челюсти, в задней ее части, по 1 ряду клыков. Выемка хвостового плавника очень глубокая. В боковой линии 39—46 прободенных чешуй. Мягких лучей в спинном плавнике 15—17.
37 (38). Колючая часть спинного плавника не покрыта чешуей. Подкрышечная и межкрышечная кости гладкие или слабо зазубрены по нижнему краю. 18. [Pseudanthias Bleeker].
38 (37). Колючая часть спинного плавника покрыта мелкой чешуей. Подкрышечная и межкрышечная кости зазубрены. 19. Franzia Jordan et Thompson.
Рыбы северных морей СССР. А.П.Андрияшев 1954 г.
Подотряд Percoidei
XXXI. Сем. SERRANIDAE — СЕРРАНОВЫЕ
Берг, Рыбы пресн. вод СССР. III, 1949 : 1011.
Колючая часть спинного плавника обычно хорошо развита, мягкая его часть обычно не на много длиннее анального. Брюшные плавники на груди, без аксиллярной лопасти, с одной колючкой и не более чем с 5 ветвистыми лучами. В начале анального плавника обычно 3 колючки. Рот выдвижной. Верхнечелюстная кость сзади расширена, обычно свободна, реже скрыта под предглазничной. Вторая подглазничная косточка снабжена пластинкой, поддерживающей глазное яблоко. По 2 postcleithra с каждой стороны (Берг).
Много родов, главным образом морских, в умеренных и тропических частях всех океанов. На севере один род: Morone
Рыбы пресных вод СССР и сопредельных стран. Л.С.Берг 1948 г.
XXV. Семейство SERRANIDAE
Regan, Ann. Mag. Nat. Hist. (8), XII, 1913, p. 113.
Колючая часть спинного плавника обыкновенно хорошо развита; мягкая часть спинного плавника обыкновенно не на много длиннее анального. Брюшные плавники на груди, без аксиллярной лопасти, с колючкой и не более чем 5 ветвистыми лучами. Колючек в анальном плавнике обычно 3. В хвостовом плавнике, как правило, 17 развитых лучей (из них 15 ветвистых). 4 жабры, за 4-й — щель. 5—8 лучей жаберной перепонки. Жаберные перепонки свободны от межжаберного промежутка. Псевдобранхии обычно есть. С каждой стороны по две ноздри. Рот выдвижной; щетинковидные или гребневидные зубы на челюстях и обычно на нёбе. Вторая подглазная косточка снабжена пластинкой, поддерживающей глазное яблоко. Верхнечелюстная кость сзади расширена, обычно свободна, редко скрыта под предглазничной. Две заднеключичных кости с каждой стороны. 24 позвонка или более; первые 2 или более без парапофизов; некоторые или все ребра прикреплены к парапофизам.
Много родов и видов, главным образом морских, в умеренных и тропических частях всех океанов. В пресных водах СССР:
1 (4). Спинные плавники отделены друг от друга.
2 (3). На языке нет зубов. На верхнечелюстной есть придаточная косточка. 1. Lateolabrax Bleeker
3 (2). На языке есть зубы. На верхнечелюстной нет придаточной косточки. 2. Morone Mitchill
4 (1). Спинные плавники слиты вместе. На языке нет зубов. На верхнечелюстной есть придаточная косточка. 3. Siniperca Gill