Gadus morhua Linnaeus, 1758
Атлантическая треска, Обыкновенная треска, Океаническая треска, Американская треска, Ньюфаундлендская треска, Европейская треска, Балтийская треска, Кильдинская треска, Беломорская зимняя треска, Беломорская мигрирующая треска, Баренцовоморская треска, Новоземельская треска, Лофотенская треска, Мурманская треска, Мурманская мигрирующая треска, Мурманская фиордовая треска, Мурманская прибрежная треска, Турянка, Шпицбергенская треска, Медвежинско-шпицбергенская треска, Гадус морхуа (Rus),
Atlantic cod, Northern cod, Baltic cod, Kil'din cod, Newfoundland cod, Bank cod, Shoal cod, Shore cod, Trap cod, Red-cod, Codling, Foxy tom-cod, Harbour tom-cod, Seal-head cod, Inshore cod, Newfoundland fish, Mother fish, Winter fish, Grog fish, Logy fish, Berry fish, BlackBerry fish, Old soaker, Codde, Loader, Haberdine, Breeder, Schrod, Bacalao, Scraud, Swallow tail, Bastard, Atlanticheskaya treska (Eng).
Синонимы:
Gadus arenosus Mitchill, 1815
Gadus callarias hiemalis Taliev, 1931
Gadus callarias kildinensis Derjugin, 1920
Gadus callarias Linnaeus, 1758
Gadus heteroglossus Walbaum, 1792
Gadus morhua Linnaeus, 1758
Gadus morhua callarias Linnaeus, 1758 Балтийская треска (Rus), Baltic cod (Eng),
Gadus morhua kildinensis (Derjugin, 1920) Кильдинская треска (Rus),
Gadus morhua morhua Linnaeus, 1758
Gadus morhua morhua n. hiemalis Taliev, 1931 Беломорская зимняя треска (Rus),
Gadus nanus Faber, 1829
Gadus ruber Lacepède, 1803
Gadus rupestris Mitchill, 1815
Gadus vertagus Walbaum, 1792
Morhua punctatus Fleming, 1828
Morhua vulgaris Fleming, 1828
Morrhua americana Storer, 1839
Morrhua callarias (Linnaeus, 1758)
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
Треска атлантическая — Gadus morhua morhua (Linnaeus, 1758)
Наиболее характерный представитель отряда и семейства. Тело длинное, веретенообразное. У этого и всех последующих видов три спинных и два анальных плавника. Хвостовой плавник прямой, без выемки. Голова большая с хорошо развитым подбородочным усиком и большим конечным ртом. Позади и над основаниями грудных плавников боковая линия плавно изогнута. D1 14—15. D2 18—22. D3 17—20. А1 19—23. А2 15—19. Окраска тела и головы зеленовато-оливковая, по бокам, спине и плавникам разбросаны темные пятна разной формы и размера. Брюхо светлое, белое. Собственно атлантическая треска достигает длины более 1,5 м и массы порядка 30 кг. В Баренцевом море в уловах преобладают особи длиной тела 60—80 см и массой около 6 кг. Обычный вид Северной Атлантики, у наших берегов обычен в Баренцевом море (особенно молодь).
Треска балтийская — Gadus morhua callarias (Linnaeus, 1758)
Близкий подвид, отличающийся биологией, меньшими, чем у атлантической трески, размерами тела и ареалом. Длина тела не превышает 80 см. Типично солоноватоводный обитатель Балтийского моря и его заливов.
! Треска кильдинская — Gadus morhua kildinensis (Derjugin, 1920)
Редчайший подвид атлантической трески. Имеет серьезные экологические отличия, меньшие размеры и более темную (серо-зеленую) окраску. D1 12—14. D2 16—20. D3 17—20. А1 18—22. А2 17—20. Длина тела, очевидно, не более 60 см. Обитает в уникальном озере Могильное (остров Кильдин, юг Баренцева моря), характеризующемся «слоеной» водой (слои морской воды перемежаются со слоями пресной). Численность единственной уникальной популяции крайне мала и не изучена. Требует строжайшей охраны и включения в Красные книги.
Редкие и исчезающие животные. Рыбы Под редакцией академика В.Е.Соколова 1994 г.
Кильдинская треска Gadus morhua kildinensis (Derjugin, 1920)
Редкий подвид трески, находящийся под угрозой исчезновения (I категория). Намечен к внесению в Красную книгу России.
Достигает длины 69 см и массы 2,4 кг. Продолжительность жизни не превышает 7 лет. Окрашена своеобразно и чрезвычайно ярко. У взрослых экземпляров на спине и боках крупные коричневые с буроватым оттенком пятна, обрамленные светлой каймой, которые заходят и на голову. Нижняя часть головы и брюхо ослепительно белые. Мелкая треска (до 30 см длиной) окрашена иначе: темная, почти черная спинка, темно-серые бока и светлое брюшко.
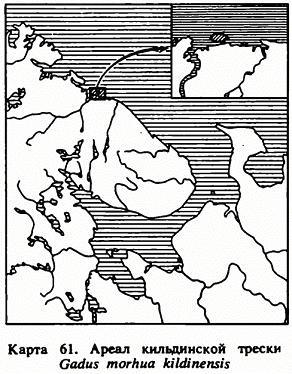
Эта уникальная треска обитает в реликтовом оз. Могильное, располагающемся в юго-восточной части о. Кильдин у побережья Мурмана в Баренцевом море (карта 61). Верхние слои этого озера, имеющего в длину чуть более 0,5 км и максимальную глубину 16,3 м, содержат пресную воду, а нижние заражены сероводородом. Треска живет здесь в слое воды с соленостью от 8—12 до 27—28‰ , составляющем по вертикали всего 3—4 м. Впервые о ее существовании в оз. Могильном упоминается в 1809 г. акад. Н. Я. Озерецковским.
В этом озере треска придерживается хорошо освещенных берегов около свалов, покрытых галькой, валунами и заросших
водорослями; молодь концентрируется главным образом в центральных районах озера над глубинами, зараженными сероводородом. Самки достигают половой зрелости в возрасте 5—6 лет при длине около 50 см, самцы — в 3—4 года, имея размеры чуть более 40 см. Плодовитость известна всего для одной самки, у которой в яичниках было 1,4 млн. икринок. Нерест порционный, растянут с конца марта до конца июня; разгар его в мае и первой половине июня. Икра пелагическая. Размножается в центральной части озера в очень узком слое воды с соленостью 22—28‰ на глубине 6,5—8 м от поверхности. Температура воды в этот
период составляет 4—5,5°С. Основным пищевым компонентом кильдинской трески являются гаммарусы; значительно реже в желудках встречаются другие беспозвоночные — изоподы, полихеты, хирономиды, а также рыба (маслюк, колюшка, молодь трески).
Численность трески в озере всегда была очень невысокой, а в настоящее время измеряется всего несколькими десятками особей. Основными факторами, негативно воздействующими на состояние ее популяции, являются браконьерский вылов и загрязнение озера нефтепродуктами; в последние десятилетия наблюдается также подъем сероводородной зоны к поверхности. Меры охраны этой интереснейшей формы трески не разработаны.
Иллюстрированная энциклопедия рыб. Ст.Франк 1983 г.
Из семейства тресковые (Gadidae) наиболее известна треска обыкновенная, или атлантическая (Cadus morhua), достигающая 150 см и веса 40 кг. Живет в северной части Атлантического океана у американских и европейских берегов, а также в Балтийском и Белом морях. Образует ряд подвидов. Нерест происходит с января по март. Весьма многочисленные икринки (1,5 - 10 миллионов штук) всегда пелагические.
Треску обрабатывают по-разному: ее сушат, что проще всего, но чаще снимают кожу, отделяют мясо от костей, тщательно упаковывают, замораживают и продают под названием рыбное «филе».
Словарь названий пресноводных рыб СССР. Г.У.Линдберг и А.С.Герд 1972 г.
19.4.1 (1). Gadus morhua Linne, 1758 — Атлантическая треска (Б. : 950).
[1] русск. Атлантическая треска — Б., 1933 : 744. Обыкновенная треска — А. Ник. : 579. [14] дат. Torsk — Hoek : 31. [15] шведск. Gadda — Б., 19406 : 18. Kabelja — Hoek : 31. Torsk — Б., 19406:23. [16] норв. Torsk - Hoek : 31. [18] англ. Atlantic cod Канада — Ricker : 127. Cod — Hoek : 31; Poll : 195; Европа и Сев. Африка — Мартинсен, 1961 : 93. [19] нем. Dorsch, Dösch, Gad morh — Benecke : 87. Kabeljau — Hoek : 31. Pamuchel, Permochel, Pomochel, Pomuchel — Benecke : 87. [20] голланд. Dogger, Dorsch, Gul (молодь), Kabeljaum, Kolvisch, Muiltje — Poll : 195. [21] флам. Kabeljaw — Hoek : 31. [22] франц. Cabiau валл., Cabiawe валл. — Poll : 195. Cabillaud — Hoek : 31; Poll : 195; Names of fishes : 38. Cabonau валл. — Poll : 195. Morue — Hoek : 31; Names of fishes : 38.
19.4.1 (la). Gadus morhua morhua n. hiemalis Taliev, 1931 — Беломорская зимняя треска (Б. : 951).
[1] русск. Беломорская зимняя треска — Св., 1948:171. Мигрирующая треска — Андрияшев, 1954 : 171. Пришлая морская треска — РБМ : 92.
19.4.1 (2). Gadus morhua callarias Linnaeus, 1758 — Балтийская треска (Б. : 951).
[1] русск. Балтийская треска — Б., 1933:744. Менза — Зол. Ментуз Балт. м.: Литва — Гейнем. : 520. ~ Дорш — Торговый устав 1724 г.; Теряев : 46; Ловецкий : 468. Треска — Кн. тамож. Вел. Устюга, 1633 г.; Поликарпов; Краш., 1 : 307; Леке. нач. XVIII в.; Росс, целл.; Лепехин, I : 517, IV : 328; Паллас, III, 1 : 40; Начерт. : 388; Озерецк., 1791 : 42; Блуменбах : 381; САР; Двиг. : 132; Теряев : 46; Ловецкий : 467; Сл. 1847 г. Treska — Сл. Р. Джемса. [4] польск. Pomuchel кашуб., Pomuchla кашуб. — Benecke : 87. [12] латыш. Menca — ZK. [13] лит. Menke, Menzas — Benecke : 87. [18] англ. Baltic cod Канада — Ricker : 127.
19.4.1 (3). Gadus morhua kildinensis Derjugin, 1920 Кильдинская треска (Б. : 952).
[1] русск. Кильдинская треска — Св., 1948 : 175.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Самых крупных
размеров достигают хищные: атлантическая
треска (Gadus morhua) и мольва (Molva
molva), для которых известны экземпляры
длиной до 180 см.
Треска (Gadus morhua) — наиболее
многочисленный вид семейства, с самым
широким ареалом, охватывающим бореальную (умеренную) область Атлантического
и Тихого океанов, образует несколько
подвидов и значительное число рас.
Имеет три спинных и два анальных
плавника, крупную голову с большим
конечным ртом. Усик хорошо развит. Окраска
сильно варьирует; цвет спины зеленый
или желтовато-коричневый, иногда
бурый, с многочисленными мелкими
желтовато-коричневыми пятнами. Бока того же
тона, более светлые, брюхо желтоватое
или белое. Боковая линия светлая,
хорошо заметна на протяжении всего тела,
образует небольшой изгиб над грудным
плавником.
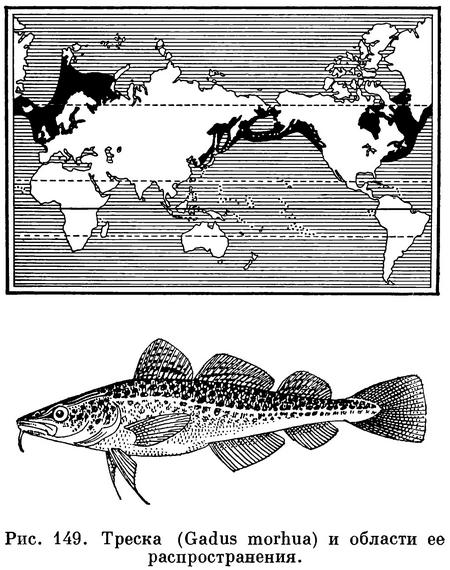
В Атлантическом океане треска
распространена от мыса Код и Бискайского
залива до Гренландии, Шпицбергена и
Новой Земли. В Тихом океане треска
известна от Берингова пролива до Калифорнии
и Кореи. Треска — эврифаг,
использующий все виды пищи от
планктона до сравнительно крупных рыб.
Наиболее многочисленна атлантическая
треска (Gadus morhua morhua),
содержащая несколько рас (стад), жизненный
цикл которых привязан к системам
течений северной части Атлантического
океана и прилегающих областей Ледовитого
океана. Главные из рас: норвежско-баренцевоморская, или арктическая,
размножающаяся у берегов Норвегии от Мере
до Северных Лофотенских островов и
откармливающаяся в Баренцевом море и
на медвежинско-шпицбергенском
мелководье; исландско-гренландская,
связанная с течением Ирмингера,
размножающаяся у южных и юго-западных берегов Исландии и мигрирующая к
северным берегам острова и к Гренландии; лабрадорско-ньюфаундлендская, связанная
с круговоротом теплых атлантических вод,
проникающих в Девисов пролив,
размножающаяся у Северного Лабрадора,
откуда молодь дрейфует к северо-западным
склонам ньюфаундлендского мелководья;
ньюфаундлендская, обитающая вдоль
фронтальной зоны вод шельфа и склона
и Гольфстрима от мыса Код до
западного свала Ньюфаундлендской банки.
Наиболее широко распространенной и
мощной является норвежско-баренцевоморская раса. Особи ее достигают
наибольших размеров: обычная величина 40—80 см, но может достигать и 150—180 см
длины и свыше 40 кг веса. Основные
нерестилища этой расы находятся в Вест-фьорде, у Лофотенских островов,
Северо-Западной Норвегии, почему ее часто
называют лофотенской. Нерест происходит в
марте — апреле на глубине до 100 м,
на границе теплых подстилающих вод
атлантического происхождения и местных,
более холодных и несколько опресненных
вод фиордов.
Выметанные и оплодотворенные
пелагические икринки подхватываются
течением. Несомые течением икринки и
выклюнувшиеся из них личинки
проплывают до 200 км. Первые дни личинки
питаются за счет желточного пузыря, а
затем переходят на активное внешнее
питание мелкими формами планктона. В
соответствии с направлением течений часть
молоди попадает к Медвежьему острову,
но большое количество ее с Нордкапским
течением приносится в Баренцево море.
Все это время молодь живет в толще воды
и питается планктонным рачком — калянусом. К июлю мальки длиной 3—4 см,
дрейфующие на север, достигают 72—73° с. ш., а дрейфующие на восток —
Кольского меридиана (33° в. д.). В сентябре
молодь трески достигает восточных
районов Баренцева моря, где она переходит
к донному образу жизни.
Неоднократно сеголеток трески в
Шпицбергенском течении находили за 80° с. ш.,
т. е. уже в бассейне Северного Ледовитого
океана. Судьба их неясна. Возможно,
они погибают, но не исключено, что они
попадают в Баренцево море с водами,
проникающими между Шпицбергеном и
Землей Франца-Иосифа.
В первые два года жизни молодь трески
малоактивна и не способна вернуться в
более теплые воды. Да она к этому и не
стремится. Молодая треска переносит
температуру 1°С и ниже, активно питаясь
в это время мелкими ракообразными.
С трехлетнего возраста треска начинает
совершать заметные миграции летом по
течению на север и восток, зимой против
течения на юг и запад. С возрастом область
миграций трески расширяется за счет
охвата восточных районов Баренцева моря. С этого времени треска становится
хищником, и рост ее резко повышается.
Трехлетние особи весят 300—350 г,
четырехлетние 600—700 г, а пятилетние
рыбы 1000—1200 г. Объектом питания
трески Баренцева моря служат в основном
три многочисленных вида планктоноядных
рыб — сельдь (преимущественно молодь),
мойва и в некоторых районах сайка.
В сравнительно теплые годы важное
значение в пище трески, особенно зимой,
приобретает сельдь, зимующая в придонном
горизонте. Весной всегда треска питается
мойвой, которую она встречает в
открытых районах моря на пути
последней из северных районов и стаи
которой сопровождает к нерестилищам у
берегов Мурмана или Северной Норвегии.
В холодные годы, когда расширяется
ареал и увеличивается численность сайки,
треска находит большие скопления ее на
востоке.
Помимо этого, летом треска часто
откармливается черноглазкой — рачками
из семейства евфаузиевых, которые
образуют плотные скопления в центральных
районах моря. Иногда она использует и
донную фауну, главным образом
двустворчатых моллюсков, у которых
откусывает вытягиваемые ими ноги. Треска
питается и собственной молодью, а
крупные особи, весом в несколько
килограммов, чаще всего питаются своими
собратьями меньших размеров. В треске весом в
несколько килограммов можно встретить
иногда треску весом в килограмм, а в
последней — десятки сеголеток или
годовиков трески.
Восьми — десяти лет, весом 3—4 кг, баренцевоморская треска начинает
готовиться к продолжению своего рода. В
последнее лето она посещает дальние
восточные или северные районы моря,
особенно богатые пищей. Свой откорм она
заканчивает в сентябре — октябре,
собирается в большие стаи в районах,
находящихся под воздействием теплого Нордкапского течения, и начинает свою
миграцию к Лофотенским островам. Этот путь,
протяженностью свыше 1500 км, она
совершает за 5—6 месяцев со средней
скоростью 7—8 км в сутки. В пути треска
местами отдыхает, иногда питается, но
путь этот она может совершить, используя
запасы жира, накопленные в печени —
главном жировом депо тресковых,
обеспечивающем миграции и развитие гонад.
В движении трески на нерест главным
ориентиром является Нордкапское течение,
с водами которого молодая треска
проникает в Баренцево море.
Приход стай трески на нерест к
Лофотенским островам — большое событие
в жизни населения Северной Норвегии,
собирающего у порога своего дома
богатый урожай, выросший на просторах
Северного Ледовитого океана. В этом
промысле принимает участие все население, тысячи мелких моторных ботов с
несложным вооружением в виде ручных
удочек, ярусов и сетей устремляются по
общему для всех сигналу к стаям трески.
Желающих ловить треску так много, что
все боты разделяются на две группы, одна
из которых по очереди облавливает
особенно мощные скопления, а другая в этот
день использует второстепенные
скопления. С субботы до понедельника лов
трески полностью прекращается. Для
рыбаков наступает долгожданный отдых и
время на ремонт орудий лова, треске
предоставляется возможность выполнить долг
по воспроизведению своего потомства.
Каждая самка выметывает 2—3 и даже
4 порции икры, это заставляет ее пробыть
на нерестилищах несколько недель; столь
же долго, а иногда и больше времени,
держатся на нерестилищах самцы,
участвующие в оплодотворении многих самок. Не
удивительно поэтому, что интенсивность
промысла на нерестилищах очень велика
и из каждых двух рыб, пришедших на
нерестилища, одна изымается промыслом,
а в некоторые годы из трех пришедших
рыб вылавливается две.
Отнерестовавшие особи устремляются
к местам нагула, чтобы восполнить
потерянные силы, откормиться и через год
вновь появиться на нерестилищах для
продолжения рода. Треска может жить до
20—25 лет. Далеко в открытом море над
большими глубинами она встречается
редко, хотя там достаточно пищи в виде
жирующей атлантическо-скандинавской
сельди.
Исландско-гренландская треска своей
биологией напоминает норвежско-баренцевоморскую, она также совершает
дальние миграции и обладает большим
нагульным ареалом и небольшим
нерестовым. Треска северо-западной части
Атлантического океана (лабрадорско-ныофаундлендская) образует местные
стада; снос молоди течениями и активные
миграции имеют в ее жизни меньшее
значение.
Биология отдельных рас этого
района изучена недостаточно. Яснее других
вырисовывается миграционный цикл
трески, размножающейся у Северного
Лабрадора, совершающей нагульные миграции
к северным склонам ньюфаундлендского
мелководья.
Особые подвиды трески живут в
Балтийском и Белом морях. Они приспособились
к жизни в этих опресненных морях, не
совершают дальних миграций, созревают
раньше атлантической трески, на 3—4-м
году жизни. Размеры их особей меньше,
особенно в Белом море.
Балтийская треска (Gadus morhua callarias) — одна из важнейших промысловых
рыб Балтийского моря. Она достигает
длины свыше 100 см и веса 11 кг.
Созревает на третьем году жизни, по
достижении длины 20—27 см, живет до 10 лет.
Основную пищу молоди трески на
первом-втором году жизни составляют
ракообразные и полихеты. С ростом рыбы
на смену ракообразным приходит рыба и
главной пищей трески становятся салака
и шпрот. Нерест трески происходит в
западной части моря в зимне-осеннее время,
в районах впадин Балтийского моря на
глубине 80—100 м при солености 12—18‰ и температуре 4—5° С.
Атлантическая треска — одна из
важнейших промысловых рыб мира. Давая
улов 2,6—4 млн. т в год, она занимает
по объему улова третье место в мире, за
перуанским анчоусом и атлантической
сельдью, определяя 5—6% всего мирового
улова.
Треска пользуется широким спросом
на рынках всех европейских стран. Она
доставляется в парном или замороженном
виде, особенно ценится приготовленное из
нее филе. Жир из тресковой печени богат
витаминами А и D и представляет собой
ценный медицинский препарат, особенно
для детей, против заболевания рахитом.
В Норвегии заготавливают также икру
трески, которая продается в небольших
тюбиках и заслуженно пользуется
спросом во многих странах Европы.
Рыбы северных морей СССР. А.П.Андрияшев 1954 г.
la. Gadus morhua morhua Linne — Атлантическая треска.
Gadus morhua Linne, Syst. Nat., ed. X, 1758 : 252 (Атлантич. океан у берегов Европы). — Берг, Рыбы пресн. вод СССР, II, 1933 : 744, фиг. 715.
Gadus callarias Linne, 1758 : 252 (частью: Атлантич. океан у берегов Европы). — Williamson, Ann. Rep. Fish. Board Scottl., XX, 3, 1902 : 228, табл. (череп). — Mиxин, Изв. Отд. прикл. ихт., III, 2, 1925 : 189 (Мурман; биометрика). — Липин, Тр. Научн. инст. рыбн. хоз., II, 4, 1928 : 31 (измерения черепов). — J. Schmidt, Compt. Rend. Trav. Labor. Carlsberg, XVIII, 6, 1930 : 71 стр., 10 карт (вариация числа лучей во II D и позвонков в связи с условиями обитания). — Есипов, Тр. Инст. изуч. Севера, 48, 1931 : 75, цветн. рис. (биометрическое сравнение турянки и океанической трески). — Дементьева, Плечкова, Розанова и Танасийчук, Докл. 1-й сессии Гос. Океаногр. инст., 2, 1932 : 49 (число лучей в плавниках, позвонков, жаберных тычинок, измерения черепов, расы). — Сынкова, Изв. Ихт. инст., XIII, 2, 1932 : 89 (сравнение новоземельской трески с мурманской). — Dannevig, Journ. Conseil., VII, 1, 1932 : 60 (зависимость числа позвонков от условий развития). — Vladykov, Contr. Canad. Biol. Fish., VIII, 29, 1933 : 26 (Гудсонов прол.; отличия от Gadus ogac). — Дементьева и Танасийчук, За рыбн. индустр. Севера, 5, 1935 : 40 (расы, обзор литературы).
Gadus arenosus Мitсhill, Trans. Litt. Philos. Soc. New York, I, 1815 : 318 (Нью-Йорк).
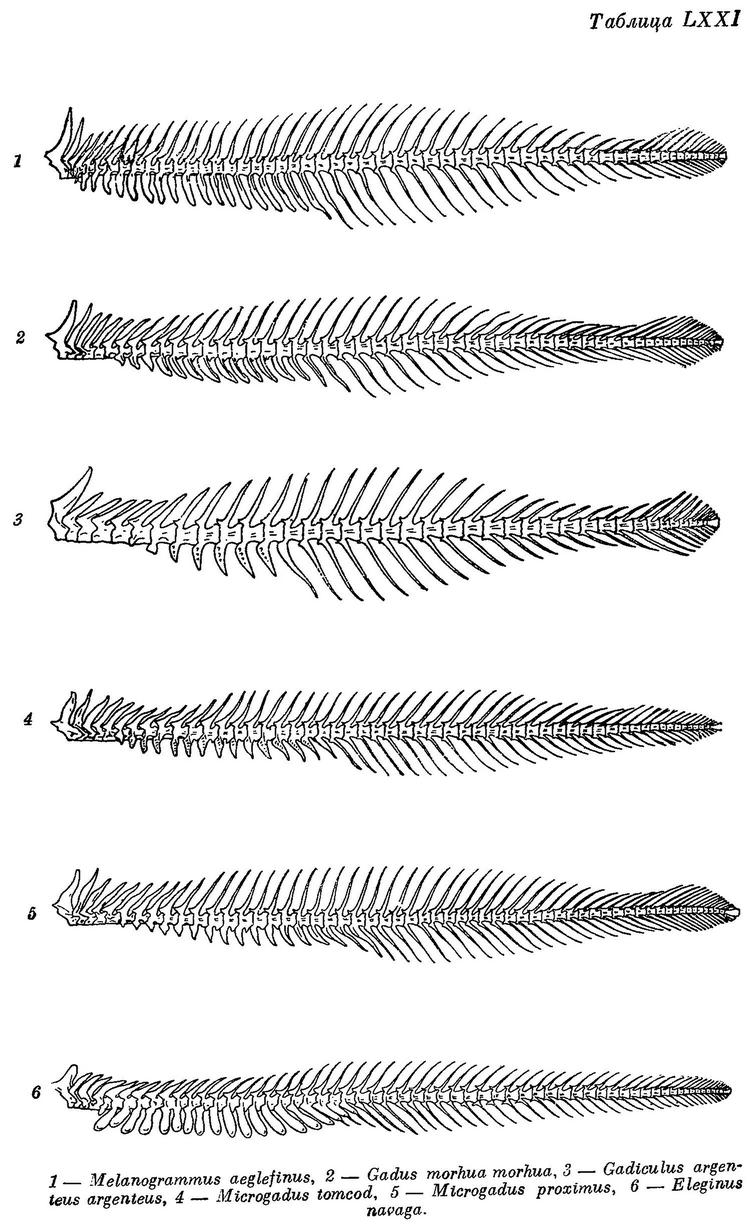
Gadus morhua morhua Световидов, 1948 : 165, фиг. 26—27, табл. X, LI, LXXI (библиография, остеология, распространение, сводка биологических сведений). — Маслов в изд.: Промысл. рыбы СССР, 1949 : 498, атлас табл. 152 (сводка, цветн. рис.).
Распространение, биология. Smitt, Scand. Fish., I, 1893 : 472, pl. XXII, fig. 3 (частью: Атлантич. океан у берегов Европы и Америки; биология, цветн. рис.). — Брейтфус и Гебель, Мат. естеств. истории трески и пикши, 1908 : 113 (Баренц. м., биология, расы). — Ehrenbaum, Eier und Larven, 1905—1909 : 224, fig. 84 (развитие). — J. Schmidt, Rapp. Proc.-Verb., X, 1909 : 17 (нерест у берегов Европы). — Bigelow a. Welsh, Fish. Gulf Maine, 1925 : 409, fgs. 204—210 (на север до Лабрадора, единично до Гудсонова зал.; биология). — Книпович, Определитель, 1926 : 150, фиг. 116 (на Мурмане до 144 см). — Jensen, Rapp. Proc.-Verb., XXXIX, 1926 : 85 (нерест у юго-зап. берегов Гренландии; питание). — Сомов, Сб. в честь Н. М. Книповича, 1927 : 365 (нерест на Мурмане). — Суворов, Тр. Инст. изуч. Севера, 38, 1927 : 38 (возраст). — Тарасов, там же, 48, 1931 : 50 (нерест в Мотовск. зал.). — Талиев, Изв. Отд. прикл. ихт., XI, 2, 1931 : 137 (Поньгома у входа в Онежск. зал.). — Есипов, Кучина, Суворов, Вадова, Сб. научно-пром. работ на Мурмане, 1932 (нерест, размеры, темп роста, вес, питание и др.). — Гурвич, Изв. Ихт. инст., XIII, 2, 1932 : 101 (юго-зап. побережье Нов. Земли; биометрика, возраст, питание). — Броцкая, Докл. 1-й сессии Гос. Океаногр. инст., 4, 1933 : 20 (питание). — Расс, там же, 5, 1933 : 4 (нерест). — Пробатов, Мат. научно-пром. обслед. Карск. губы, 1934 : 94 (Карск. губа). — Еhrеnbaum, Handb. Seefischer. N. Europas, II, 1936 : 94, fig. 74—75 (биология, уловы). — Агапов и Топорков, Пробл. Арктики, 2, 1937 : 107 (юго-зап. берег Нов. Земли). — Владимиров, Учен. зап. Пермск. Гос. унив., III, 1937 : 95 (биология новоземельской трески). — Шапошникова, Тр. Аркт. инст., 100, 1937 : 47—58 (губы Грибовая, Самойловича и Моллера на Нов. Земле, Костин Шар; питание). — Есипов, Промысл. рыбы Баренц. м., 1937 : 76 (биология, химический состав, уловы). — Шмит, За рыбн. индустр. Севера, 2—3, 1937 : 47 (нерест). — Дубровский, Природа, 11—12, 1938 : 105 (зал. Шуберта на юго-вост. побережье Нов. Земли; вмерзшая в лед). — Задульская и Смирнов, Тр. Всес. Научно-иссл. инст. морск. рыбн. хоз. океаногр., IV, 1939 : 321 (суточный ход питания). — Месяцев, там же : 369 (о структуре косяков). — Камерницкая, там же : 396 (различия в структуре косяков). — Перцева, там же : 420 (нерест в Мотовск. зал.). — Марти, Тр. Полярн. научно-иссл. инст. морск. рыбн. хоз. океаногр., 3, 1939 : 3 (биология, расовый состав прибрежной мурманской трески). — Зацепин и Петрова, там же, 5, 1939 : 170 стр. (всестороннее исследование питания трески в Баренц. м. по 162 тыс. экз.). — Маслов, там же, 6, 1939 : 70 (траловый промысел, запасы). — Глебов, Природа, 4, 1941 : 87 (биология). — Sund in: Andersson, Fiskar Fiske Norden, I, 1942 : 152, pl. 48 (биология, цветн. фотогр.). — Маслов, Тр. Полярн. научно-иссл. инст. морск. рыбн. хоз. океаногр., 8, 1944 : 50—140 и др. (промыслово-биологич. исслед.). — Saemundsson, Zool. Iceland, IV, 72, 1949 : 53 (биология, уловы). — Маслов в изд.: Промысл. рыбы Баренц. Белого м., 1952 : 125 (обстоятельный биологич. очерк).
I D 14—15, II D 18—22, III D 17—20, I А 19—23, II А (15) 17—19 (Световидов), Р 18—21, позвонков (49) 51—55 (56), наичаще 53—54. В Баренцовом море на основании изучения нескольких тысяч экземпляров трески (Дементьева и др.) получены следующие колебания средних чисел: I D 13.8—14.8, II D 19.2—20.3, III D 18.8—19.4, I А 21.1—22.4, II А 18.5—19.3, позвонков 52.1—52.6 (без уростиля), жаберных тычинок 24.1—25.0. Жаберных тычинок 21—28.
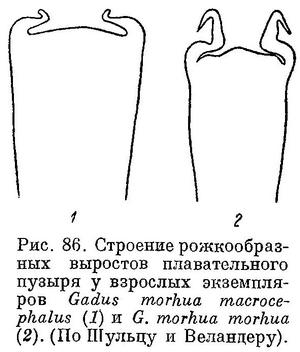
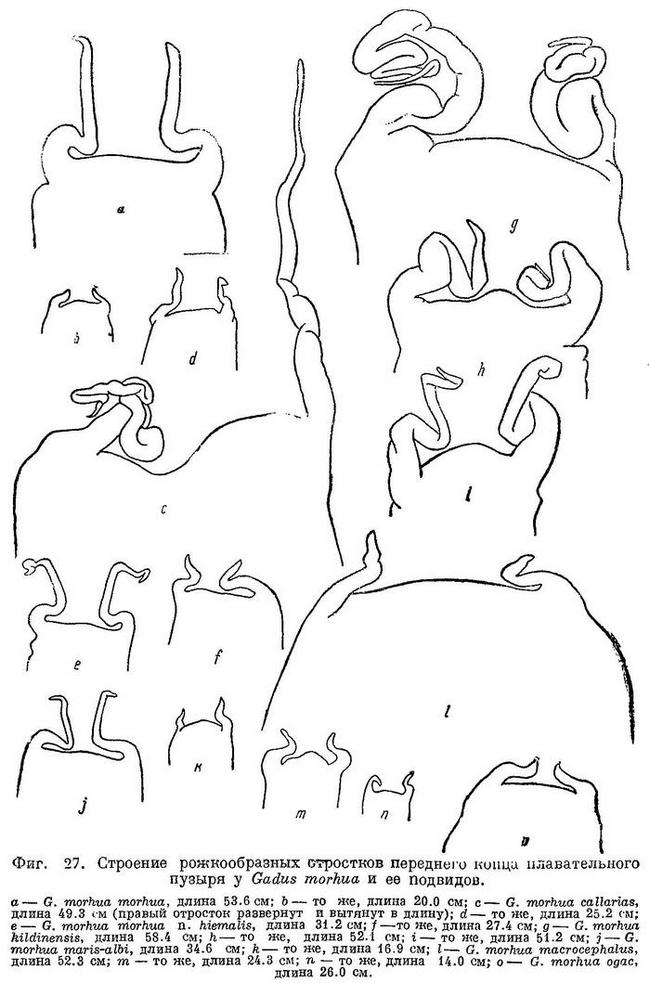
Рожкообразные выросты в передней части плавательного пузыря у взрослых длинные, обычно троекратно изогнутые — к средней линии, вперед и наружу (рис. 86, 2). Голова небольшая и узкая, длина ее, по Световидову, 24.6—26.6 % L, ширина межглазничного промежутка (костного) — 15.8—20.7% длины головы. Антедорсальное расстояние 28.1—31.5% L. 1-й спинной плавник высокий, его прижатые к спине лучи обычно значительно заходят за начало II D. Усик короткий, наичаще менее диаметра глаза, 11.7—15.6% длины головы. Окраска сильно варьирует; верхняя часть тела серовато- или зеленовато-оливковая с многочисленными мелкими желтоватыми или коричневатыми пятнами. Непарные плавники темные, пятнистые; края спинных и хвостового плавников без светлой оторочки. Мелкая прибрежная треска, в период жизни в зоне ламинарий (на Мурмане «турянка») имеет оранжево-красную или желтоватую окраску («morpha erythrina»). Длина до 100—110 см, в редких случаях до 150—180 см. Самая крупная треска в Баренцовом море имела длину 169 см и вес около 40 кг.
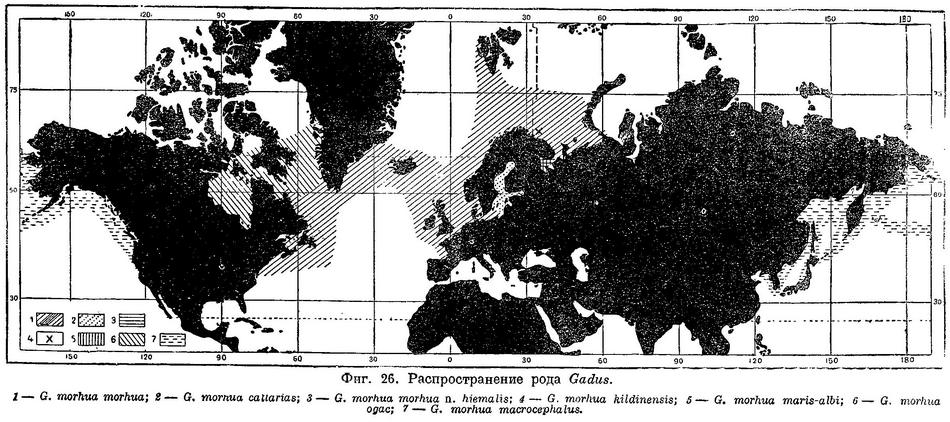
Атлантическая треска на протяжении своего огромного ареала образует ряд подвидов и рас, отличающихся средними числами лучей в плавниках и позвонков, жаберных тычинок, некоторыми пластическими признаками, окраской, местом и временем нереста, темпом роста и т. д. Наибольшие отличия (подвидовые) имеет треска беломорская (Gadus morhua maris-albi Derjugin), балтийская (Gadus morhua callarias L.) и кильдинская (Gadus morhua kildinensis Derjugin). Треска американского побережья отличается от европейской в среднем большим количеством лучей и позвонков. Такого же характера отличия, независимо от географического района, имеет мигрирующая океаническая треска по сравнению с жилой фиордовой. Так, у местной трески из зал. Св. Лаврентия число позвонков в среднем 53.6—53.9, а у мигрирующей ньюфаундлендской формы — 54.9—55.3; соответственно в Тронгеймском районе — в фиорде 52.4, в море 53.8. Аналогичные, хотя и менее выраженные различия имеет лофотенская и мурманская треска. В Баренцовом море в настоящее время большинство исследователей различает две (Некоторые авторы различают среди мурманской трески мигрирующую и фиордовую формы.) основные расы: мурманскую и океаническую, или лофотенскую, которая играет основную роль в нашем тресковом промысле. Возможно, что выделения в самостоятельную расу заслуживает и медвежинско-шпицбергенская треска, нерестующая в районе о. Медвежьего.
Распространение. Северная часть Атлантического океана. У берегов Европы повсеместно от Бискайского залива до Баренцова моря, где в западной части моря доходит на север до крайних северо-западных фиордов Шпицбергена, о. Надежды и северных склонов Центральной возвышенности (76° с. ш.), но отсутствует на глубинах Центральной впадины. По данным В. А. Бородатова, единичные экземпляры встречаются и на возвышенности Персея, и у островов Короля Карла (78°50' с. ш.), а несколько южнее 78° с. ш. треска уже попадает в трал в полупромысловых количествах. У берегов Нов. Земли на север по крайней мере до п-ова Адмиралтейства. В юго-восточной части Баренцова моря до Карских ворот и Югорского Шара; единичные экземпляры могут заходить и в юго-западную часть Карского моря (Карская губа, зал. Кротова на Новой Земле). Атлантическая треска нередко заходит также в ареал своих подвидов в Белом и Балтийском морях. У берегов Сев. Америки от м. Гаттераса до Гудсонова и Дэвисова проливов, но обычна от Лабрадора до Нью-Йорка. До недавнего времени лишь изредка попадалась у южных берегов Гренландии, однако с 1917 г. стала встречаться чаще и продвинулась на север по западному побережью до зал. Диско и даже до Упернавика (около 72.5° с. ш.). Многочисленна вокруг Исландии.
Биология. Обитает в придонных слоях воды (как у самого дна, так и на расстоянии нескольких десятков метров от него) в пределах континентальной ступени, обычно на глубине до 250—300 м, реже до 500 м. Треска менее теплолюбива, чем пикша, температурные ее пределы весьма широки, в Баренцовом море наичаще 0—5°; лофотенская треска более теплолюбива, чем мурманская. В сильно опресненных районах, как правило, не встречается, но известны единичные случаи захода в реки. Некоторые формы трески (балтийская, кильдинская, беломорская) постоянно живут при значительно пониженной солености. Нерест происходит почти по всему ареалу. Мощные нерестилища океанической трески расположены у Лофотенских островов и севернее (от Вест-фиорда до Нордкапа, частично до Мотовского залива), где нерест происходит главным образом с конца марта до начала апреля при температуре 4—6 (7)°. Главные места нереста мурманской трески находятся в Мотовском заливе (в меньших количествах и во всех губах мурманского побережья) на глубине до 100 м при температуре от —0.4 до +2.5° или несколько выше. Время нереста растянуто с февраля по июнь, в основном в апреле—мае. Плодовитость трески составляет несколько миллионов (от 0.6 до 9.3 млн) икринок диаметром 1.1—1.7 мм. Личинки, выклевывающиеся, примерно, через месяц, имеют длину около 4 мм; они долго ведут пелагический образ жизни и в это время заносятся течениями далеко от нерестилищ. Половозрелости баренцовоморская треска достигает в возрасте 8—10 лет при длине не менее 60—70 см, фиордовые формы, возможно, и при меньшей длине. Рост трески в Баренцовом море характеризуется следующими цифрами (цит. по сводке «Промысл. рыбы СССР»; длина по данным 1934—1937 гг., вес — 1936—1937 гг.):

Средние размеры трески в траловом улове Баренцова моря колеблются в зависимости от времени и места лова от 40 до 80 см. Наибольший возраст (24 года) имела треска длиной 169 см и весом около 40 кг. Наибольшие миграции совершает лофотенская треска. После нереста она продвигается для откорма вдоль берегов Мурмана и восточнее (до Новой Земли), а также на север — до Медвежинской банки и даже до Шпицбергена; поздней осенью она отходит обратно в юго-западные районы. Мурманская треска зимует, повидимому, в юго-восточной части Баренцова моря (Канинско-Колгуевский район), откуда уже в январе—феврале продвигается к берегам Мурмана для нереста. Отнерестовавшая мурманская треска откармливается в восточных районах (вплоть до Нов. Земли), а также заходит изредка в Белое море. В апреле к берегам Мурмана для откорма нерестующей мойвой подходит неполовозрелая океаническая треска, зимовавшая в центральных и западных районах Баренцова моря; это так называемая «мойвенная треска». Различают еще «ярусную» или «голодную» треску, лов которой на Мурмане происходит в мае—августе. Эта неоднородная в биологическом отношении группа состоит частично из «мойвенной» трески (после отхода мойвы), частично из отнерестовавшей лофотенской и мурманской трески и др. Питание трески, детально изученное в последнее время Зацепиным и Петровой, чрезвычайно разнообразно (более 200 пищевых объектов), но основу питания составляет не более 20—25 видов. Главной пищей взрослой трески является рыба (сельдь, мойва, молодь трески и пикши, а также сайка, камбала-ерш, Lumpenus, Lycodes, многие виды Cottidae и др.); большое значение имеют также планктонные ракообразные, так называемый «капшак» (Euphausiidae и Hyperiidae). Организмы бентоса (Hyas, Pagurus, Mesidothea, моллюски, полихеты и др.) имеют сравнительно небольшое кормовое значение и служат существенной пищей лишь в юго-восточных районах Баренцова моря.
Хозяйственное значение чрезвычайно велико — в уловах северной части Атлантического океана треска занимает второе место (после сельди). Уловы европейских стран (без СССР) достигают 10—13 млн ц, причем около трех четвертей улова приходится на долю Норвегии, Великобритании и Исландии. Наши уловы Баренцовом море достигают 2309 тыс. ц (1938 г.), что почти в 15 раз превышает уловы России в 1913 г.; эта цифра также значительно больше суммарных уловов всех иностранных государств в Баренцовом море.
1б. Gadus morhua morhua forma hiemalis Taliev — Беломорская зимняя, или мигрирующая, треска (рис. 82).
Gadus callarias hiemalis Талиев, Изв. Отд. прикл. ихт., XI, 2, 1931 : 113, фиг. 2 (Кандалакшск. зал. Белого м.).
Gadus morhua hiemalis Берг, Рыбы пресн. вод СССР, II, 1933 : 746. — Европейцева, Расов. анализ беломорск. трески, изд. Ленингр. Гос. унив., 1937 : 1—98 (биометрика, питание, вес, рост и др.).
Gadus callarias Ильин и Певзнер, Сб., посвящ. Н. М. Книповичу, 1939 : 320 (биометрика; неотличима от мурманской прибрежной трески),
Gadus morhua morhua natio hiemalis Световидов, 1948 : 171, фиг. 27, e, табл. XIII (сводка).

I D (12) 13—16, среднее 14.3, II D (17) 18—21 (22), ср. 19.5, III D (17) 18—20 (21), ср. 19.0, I A (19) 20—23 (24, 25), ср. 21.6, II A (16) 17—20 (21), ср. 18.6, позвонков 52—55 (56), наичаще 53—54. Жаберных тычинок обычно 21—26. По данным Европейцевой; число позвонков увеличено на один, так как Европейцева считала без последнего позвонка (уростиля). Сходные данные получены позднее Ильиным и Певзнер.
Морфологически не отличима от молодой мурманской трески. Детальная морфометрическая характеристика дана Европейцевой и в работе Ильина и Певзнер. Окраска пятнистая. Длина до 40—45 см, редко до 50 см, обычно 20—40 см.
Ильин и Певзнер считают эту форму (hiemalis) идентичной мурманской треске. К сходным выводам приходит и Световидов. Возможно, что «зимняя» треска Белого моря («осенняя» по Ильину и Певзнер), отличающаяся от мурманской главным образом более медленным темпом роста, не размножается, как правило, в Белом море, а заносится сюда мальками-сеголетками, которые, прожив несколько лет, выходят за пределы Белого моря (треска с замедленным темпом роста ловилась у п-ова Канин). Нами название hiemalis пока сохраняется, так как современных данных еще недостаточно для решения вопроса о всем жизненном цикле «зимней» беломорской трески и ее полном тождестве с мурманской расой.
Распространение. Белое море. В больших количествах подходит к берегам Кандалакшского залива осенью и зимой.
Биология. Нерест в Белом море точно не установлен, хотя известны единичные, близкие к зрелости экземпляры (Европейцева). Рост значительно замедлен по сравнению с ростом мурманской трески, но несколько более быстрый, чем у Gadus morhua maris-albi (по данным Европейцевой для Кандалакшского залива; обратное расчисление):

2Следует учесть, что вес второй графы относится к возрасту не 2 года, а 2+, следующая цифра веса 3+ и т. д.
Питание в основном сходно с таковым жилой беломорской трески, но крупные экземпляры питаются не колюшкой, а главным образом мойвой, также мелкой сельдью.
Хозяйственное значение небольшое, но ловится в больших количествах, чем жилая беломорская треска.
1в. Gadus morhua kildinensis Derjugin — Кильдинская треска (рис. 83).
Gadus callarias kildinensis Дерюгин, Тр. Петроградск. общ. естеств., LI, 1920 : 2 (оз. Могильное на о. Кильдине вблизи Кольск. зал.). — Дерюгин, Тр. Петергофск. инст., 2, 1925 : 78, табл. (рис.), см. также: Derjugin, Fauna Arctica, V, 2, 1928 : 545, pl. VIII. — Pace, Раб. Мурм. биолог. ст., III, 1929 : 25 (измерения 2 экз.). — Есипов, Гидробиолог. журн., IX, 4—6, 1930 : 131 (сравнение с атлантической треской; биология).
Gadus morhua kildinensis Берг, Рыбы пресн. вод СССР, II, 1933 : 746. — Световидов, 1948 : 175, табл. XI, LII (описание, измерение 8 экз., остеология).

I D 12—14, II D 16—20, III D 17—20, I A 18—22, II A 17—20, позвонков 52—54, жаберных тычинок 20—24.
Рожкообразные выросты плавательного пузыря очень длинные и обычно свернуты в клубочек (как и у балтийской трески). Ширина межглазничного промежутка 17.5—22.0% длины головы (Световидов). Прижатые к спине лучи первого спинного плавника едва заходят за начало II D. Брюшина свинцово-серебристая или свинцово-черная. Окраска темная и пятнистая — темнокоричневые круглые пятна густо разбросаны по телу, а также на хвостовом и спинных плавниках. Длина до 80 см.
Распространение. Известна лишь из оз. Могильного на о. Кильдине (восточнее входа в Кольский залив).
Биология. Условия обитания этой реликтовой формы весьма своеобразны. Живет она в небольшом озере (около полукилометра длиной) наибольшая глубина 17.5 м) с чрезвычайно своеобразным гидрологическим режимом — поверхностные воды (0—5 м) почти пресные, в нижележащей зоне (5—8 м) колебание солености весьма значительно, в среднем около 15‰; в 3-й зоне соленость 15—32‰, и, наконец, ниже 12—13 м идет сероводородная, лишенная жизни зона. Треска держится приглубых мест, но попадается и в сильно опресненных поверхностных слоях. Нерест, повидимому, в феврале—марте. На какой глубине происходит развитие икры, не известно. В июне и начале сентября ловились мальки длиной от 7 до 9.5 см. В 5 лет имеет длину в среднем 60.7 см, в 6 лет — 65.1 см (Есипов). Питается ракообразными.
Хозяйственного значения не имеет.
Фауна СССР. Рыбы. Том IX, вып.4. Трескообразные. А.Н.Световидов 1948 г.
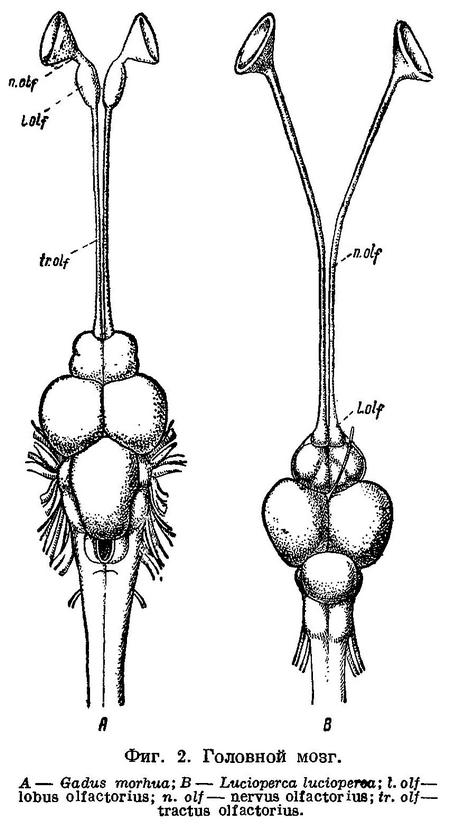
N. facialis выходит из черепа у тресковых вместе с n. trigeminus через отверстие или различной глубины выемку в передней части prooticum сбоку черепа (фиг. 5,b,с). У Brosme brosme нерв этот выходит через отверстие, а у следующего за ним по ряду Gaidropsarus mediterraneus — через глубокую выемку в prooticum, края которой настолько сближены, что почти замыкают ее спереди. У Ciliata mustela и далее следующих за ним в ряду рыб выемка становится спереди более широкой и менее глубокой. У Lota lota выемка так же глубока и края спереди сужены почти так же, как у Gaidropsarus mediterraneus, а у всех видов Molva и Merluccius n. facialis из черепа выходит опять через отверстие. Через отверстие n. facialis выходит и у следующих далее в ряду видов рода Trisopterus. При этом ширина костной перемычки у Trisopterus luscus несколько шире, чем у Trisopterus minutus, а у последней шире, чем у Trisopterus esmarkii. Odontogadus merlangus и далее следующие за ним представители семейства имеют в prooticum выемку, которая последовательно становится спереди более широкой и менее глубокой. У обеих форм Gadiculus argenteus выемки почти нет. Далее, у видов родов Eleginus, Microgadus, Boreogadus, Arctogadus и Theragra выемка постепенно углубляется, а края ее становятся более сближенными. Micromesistius имеет сбоку черепа отверстие. Таким образом, по этому признаку ряд тресковых, расположенных по степени дифференциации их непарных плавников, разделяется в свою очередь на два параллельных ряда, соответственно делению семейства на подсемейства. Такой же закономерности подчиняется изменение и некоторых других признаков.
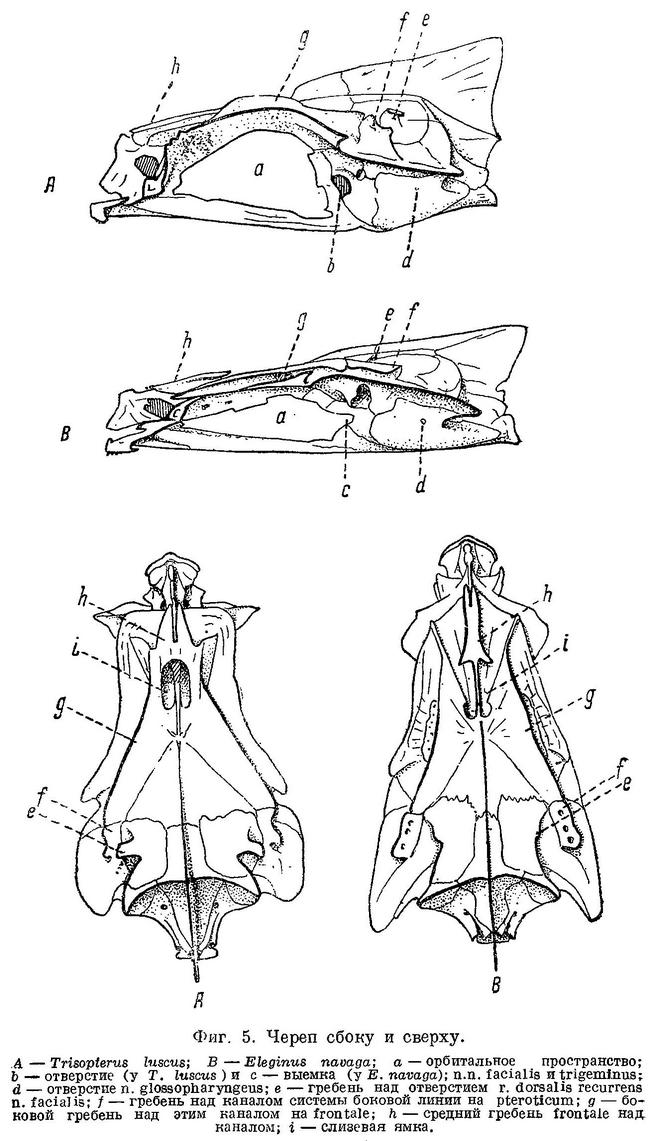
Сверху черепа тресковых на каждом parietale расположено по отверстию, через которое выходит r. dorsalis recurrens n. facialis (фиг. 5,е и рис. В на табл. XXI—LXVII). У одних тресковых каждое из этих отверстий прикрыто у каждого вида в различной степени развитыми гребнями; у других гребень не развит, и отверстие сверху открыто. У Brosme brosme и далее следующих за ним в ряду Gaidropsarus mediterraneus, Ciliata mustela, Enchelyopus cimbrius гребни, прикрывающие эти отверстия, имеются. У Raniceps ranius и всех видов родов Phycis и Urophycis отверстия r. dorsalis recurrens n. facialis не прикрыты, так как гребни у них имеют вид едва заметных выростов, расположенных несколько сзади отверстий. У Lota, всех видов Molva и Merluccius гребни развиты и прикрывают отверстия. В пределах подсемейства Gadinae гребни на parietalia развиты сильнее. У всех видов родов Trisopterus, Odontogadus, Pollachius, Gadus и Melanogrammus гребни развиты, размеры их уменьшаются от Trisopterus к Melanogrammus. Нет гребней у Gadiculus. Далее гребень есть, постепенно увеличиваясь к концу ряда, у Eleginus, Microgadus, Boreogadus, Arctogadus, Theragra и Micromesistius, причем у одних особей Eleginus navaga гребни есть, у других они не развиты.
N. glossopharyngeus (Stensiö, 1921 : 155) выходит из черепа через небольшое отверстие на opisthoticum (фиг. 5,d и рис. А на табл.XXI—LXVII). Величина этого отверстия, изменяясь в пределах ряда, подчиняется той же закономерности. У Brosme brosme отверстие почти не заметно. У Gaidropsarus mediterraneus, Ciliata mustela, Enchelyopus cimbrius, Raniceps raninus оно небольшое, у видов родов Phycis и Uropkycis несколько больше, у Lota и Molva большое и у Merluccius вновь небольших размеров. Далее, у всех видов рода Trisopterus отверстие едва заметно, размеры его постепенно увеличиваются до Gadus и Melanogrammus и, начиная с Gadiculus, уменьшаются вновь.
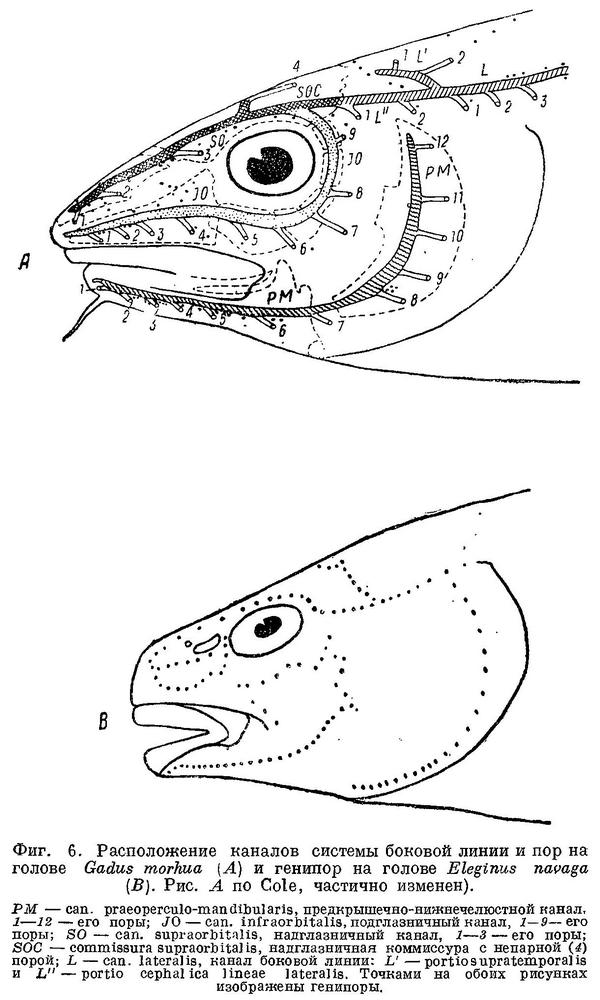
Боковая линия у тресковых образует на голове с каждой стороны каналы (фиг. 6): предкрышечно-нижнечелюстной, подглазничный и надглазничный с короткой комиссурой, соединяющей правый и левый каналы. Сама боковая линия имеет на голове надвисочную часть (L '), над которой образуется окостенение из 4—5 косточек ossa tabularia (= supratemporale). Каждый канал системы боковой линии на голове прикрыт сверху почти на всем своем протяжении гребнем тех костей, на которых эти каналы расположены (frontale, pteroticum, sub- и praeorbitale, nasale, praeoperculum, tabulare). Среди этих гребней наиболее существенное значение имеет средний гребень frontale (фиг. 5, h) и боковые гребни frontale и pteroticum (фиг. 5, f и g). Межглазничная комиссура надглазничного канала расположена в особом углублении frontale — слизевой ямке (фиг. 5, i). Слизевая ямка у одних представителей Gadidae спереди замкнута сросшимися краями боковых передних гребней над каналами системы боковой линии на frontale, у других широко открыта, так как края боковых и средних гребней frontalia укорочены и не срастаются впереди слизевой ямки. У Brosme brosme края боковых и средних гребней frontalia настолько сближены, что почти замыкают спереди небольших размеров слизевую ямку. У Gaidropsarus mediterraneus, Ciliata mustela и Enchelyopis cimbrius слизевая ямка спереди открыта, очень небольших размеров. Raniceps raninus имеет слизевую ямку спереди открытой. У одних видов родов Phycis и Urophycis слизевая ямка спереди открыта, у других закрыта; оба эти рода представляют в этом отношении небольшое отклонение от общего правила, о причинах чего будет сказано ниже. У Lota lota слизевая ямка спереди почти замкнута сближенными краями боковых и средних гребней frontalia, которые у видов рода Molva, срастаясь, образуют хорошо замкнутую ямку. Слизевая ямка у всех видов рода Merluccius представляет собой обширное углубление посередине frontalia и имеет несколько уклоняющийся от обычного для тресковых вид. Слизевая ямка замкнута далее, начиная с Trisopterus и кончая Gadiculus. У всех далее следующих в ряду видов слизевая ямка спереди открыта и замкнута вновь у стоящих в конце его Theragra chalcogramma и Micromesistius poutassou и Micromesistius australis. При этом следует отметить, что закрытая или открытая спереди слизевая ямка обычно свойственна всем видам любого рода тресковых, за исключением, как указывалось выше, родов Phycis и Urophycis. Кроме того, есть один вид — Eleginus navaga, — у которого сближенные края боковых и средних гребней frontalia иногда бывают сращены с одной стороны, а на одном из исследованных черепов и с обеих сторон. Таким образом, эта особенность находится в неустойчивом состоянии в первом случае в пределах рода, во вторых — в пределах вида, стоящих в середине каждого из параллельных рядов.
Подобная же закономерность наблюдается и в строении гребней над самыми каналами боковой линии на frontalia. У Brosme, Gaidropsarus, Ciliata, Enchelyopus, Molva, Trisopterus, Micromesistius, т. е. у рыб, стоящих в начале и конце каждого ряда, края гребней на frontalia тесно примыкают к этим костям, почти замыкая канал. Более того — у некоторых из этих рыб (Trisopterus) края гребней местами соединяются с frontalia при помощи костных перемычек. У всех остальных расположенных в ряду между этими рыбами тресковых края гребней над слизевыми каналами на frontalia обычно приподняты. Несколько уклоняется в этом признаке род Merluccius, у которого гребни развиты очень слабо.
Вдоль каждого из каналов системы боковой линии на голове расположены многочисленные группы нервных окончаний (фиг. 6, В) — генипор (pit-organs), или каналы эти открываются наружу рядом пор (фиг. 6, А), причем единичные генипоры имеются и в этом случае. Боковая линия на теле у тресковых тянется от головы до хвостового плавника в виде непрерывной или прерывистой, разделенной небольшими промежутками, линии. У Brosme brosme боковая линия тянется непрерывной полосой от головы до конца спинного плавника, становясь прерывистой лишь на хвостовом стебле. У Gaidropsarus mediterraneus, Ciliata mustela и Enchelyopus cimbrius боковая линия прерывиста на всем протяжении, у Raniceps raninus ее, повидимому, нет совершенно. У видов родов Phycis и Urophycis и у Lota lota боковая линия непрерывна почти до конца анального плавника, далее прерывиста; у видов рода Moiva она прерывиста сзади от конца анального плавника, у видов Merluccius — непрерывна на всем протяжении. У видов родов Trisopterus, Odontogadus, Pollachius и Melanogrammus боковая линия непрерывна на всем протяжении; у видов родов Gadus, Gadiculus (Экземпляры Gadiculus обычно бывают без чешуй, и поэтому о строении их боковой линии судить трудно, однако до середины третьего спинного плавника она тянется без перерывов.) и Microgadus она прерывиста сзади от середины или конца третьего спинного плавника, у видов рода Eleginus прерывиста уже сзади от начала второго спинного плавника и у видов родов Boreogadus и Arctogadus боковая линия прерывиста на всем протяжении. Далее, у Theragra chalcogramma боковая линия прерывиста лишь сзади от начала второго спинного плавника и у Micromesistius poutassou и Micromesistius australis без перерывов на всем протяжении.
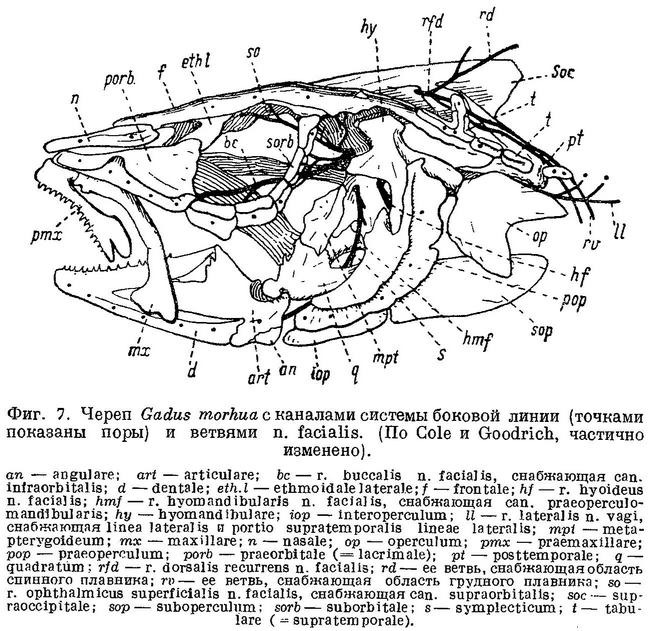
Непарные плавники, как мы видели, играют существенную роль в движении рыб. Под гребнями слизевых каналов и в слизевой ямке на frontalia расположены каналы системы боковой линии, при помощи которых, как известно, рыба ориентируется в струях воды при движении. Через отверстие или выемку в prooticum и отверстие в parietalia сверху черепа выходят ветви n. facialis, который, как известно, иннервирует разветвления органа боковой линии на голове и спинной и грудной плавники (фиг. 7, be, hmf, so, rfd). Через отверстие в opisthoticum у тресковых выходит n. glossopharyngeus (Stensio, 1921 : 125), через якобсонову анастомозу, являющуюся частью r. palatinus этого нерва, находящийся в связи с n. facialis (Cole, 1898 : 148).
* la. Gadus morhua morhua Linne — Атлантическая треска (табл. X, LI, LXXI).
— Gadus morhua Linne, Syst. nat., ed. X, 1758 : 252 (Атлантический океан у берегов Европы).
— Gadus callarias Linne, l. c. : 251 (ex parte : Атлантический океан у берегов Европы).
— Gadus vertagus Walbaum in: Artedi, Gen. pise., III, 1792 : 143.
— Gadus heteroglossus Walbaum, l. c. : 144.
— Gadus ruber Lacepède, Hist. nat. poiss., V, 1803 : 671, 673.
— Gadus arenosus Mitchill, Trans. Lit. Philos. Soc. N. Y., I, 1815 : 318 (Нью-Йорк).
— Gadus rupestris Mitchill, l. c. : 368.
— Morhua punctata Fleming, Hist. brit. animals, 1828 : 192.
— ? Gadus nanus Faber, Fish. Icelands, 1829 : 113 (Исландия).
— Morrhua americana Storer, Mem. Amer. Acad. Art. a. Sci., (n. s ), VI, II, 1858 : 343, pl. XXVII, f.4 (у берегов Сев. Америки).
— Gadus callarias Williamson, 20th. Ann. Rept. Fish. Board Scotl., pt. 3, (1901), 1902 : 228 (описание скелета).

Pаспpостpанeниe, биология. Smitt, Scand. fish., 1, 1893 : 472 (ex parte: Атлантический океан у берегов Европы и Сев. Америки, биология); Petersen, Rept. Danish Biol. St., XI, 1902 : 3; Fuiton, Publ. Circonst., 8—9, 1904 : 1 (осенний нерест в Северном море); Jensen, Medd. Komm. Havunders., Fisk., I, 7, 1905 : 8, f. 4 (69°13' N 8°23' W, отолиты); Ehrenbaura, Wiss. Meeresunters., Abt. Helgoland, VII, 1906 : 59 (у о. Медвежьего и Шпицбергена); Брейтфус и Гебель, Мат. по естеств. ист. трески и пикши, 1908 : 133 (Баренцово море, расы, биология); Hjort, Rapp. Procеs-Verb., X, 1909 : 48, f. 12 (места нереста); Damas, ibid. : 62 (биология); J. Schmidt, ibid. : 17, 139 (Атлантический океан у берегов Европы, нерест, икринки, личинки); Weigold, Wiss. Meeresunters., Abt. Helgoland, (N. F.), X, 1913 : 119 (южная часть Северного моря, мечение); Hjort, Rapp. Procеs-Verb., XX, 1914 : 81 (биология, миграция, периодичность уловов); Strubberg. Medd. Komm. Havunders., Fisk., V, 2, 1916 : 1 (опыты мечения у Фарерских о-вов); Strubberg, ibid., VIII, 1, 1922 (Каттегат, Бельты, мечение); Graham, Fish. Invest., (2), VI, 6, 1924 (биология); Bigelow a. Welsh, Fish, gulf Maine, 1925 : 409 (от мыса Гаттераса до Девисова и Гудсонова проливов); Михин, Изв. Отд. прикл. ихт., III, 2, 1925 : 189 (Мурман, биометрика); Jensen, Rapp. Proces-Verb., XXXIX, 1926 : 85 (у берегов западной Гренландии, нерест, питание); Скворцов, Бюлл. рыбн. хоз., 1927, 10 : 39 (нересту берегов Мурмана); Сомов, Сборн. в честь Книповича, 1927 : 365 (нерест у берегов Мурмана); Суворов, Тр. Инст. изуч. Севера, 38, 1927 : 38 (возраст); Суворов и Чудинов, Изв. Отд. прикл. ихт., VI, 2, 1927 : 233 (мальки); Липин, Тр. Научн. инст. рыбн. хоз., II, 4, 1928 : 31 (измерение черепов трески с Канинской банки); Idelson, Hydrobiol. Zeitschrift, VIII, 10—12, 1929 : 269 (питание); Зенкевич и др., Докл. I сессии Гос. Океаногр. инст., 4, 1931 : 20 (питание); J. Schmidt, C.-R. Trav. Labor. Carlsberg, XVIII, 6, 1930 (ex parte Атлантический океан у берегов Европы, количество позвонков и лучей во II D); Есипов, Тр. Инст. изуч. Севера, 48, 1931 : 75, рис. (Баренцово море на меридиане о. Кильдина, губа Порчниха, сравнение турянки и океанической трески); Тарасов, ibid. : 50 (Мотовский залив, нерест); Талиев, Изв. Отд. прикл. ихт., XI, 2, 1931 : 137 (Белое море у Поньгомы); Jensen a. Hansen, Rapp. Proces-Verb., LXXII, 1931 : 3—10 (у западных берегов Гренландии, нерест, питание); Idelson, Journ. Conseil, V, 3, 1931 : 432 (мечение в Баренцовом море); Schnakenbeck, Zool. Anz., 94, 1931 : 17 (р. Эльба в 15 км выше Гамбурга); Есипов, Сб. научно-промысл. работ на Мурмане, 1932 : 5 (западный Мурман, Мотовский зал., нерест); Есипов и Кучина, ibid. : 27 (западный Мурман, возраст и темп роста); Суворов и Вадова, ibid. : 67 (размеры, вес, возрастной состав); Кучина, ibid. : 103 (питание); Дементьева и др., Докл. I сессии Гос. Океаногр. инст., 2, 1932 : 49 (измерения черепов); Суворов, Изв. Ихт. инст., XIII, 2, 1932 : 61 (Мотовский зал., нерест, икринки); Сынкова, ibid. : 89 (Новая Земля, сравнение новоземельской трески с мурманской); Гурвич, ibid. : 101 (берега Новой Земли, на север до Крестовой губы, питание, размеры, возраст, биометрика); Танасийчук, Карело-мурм. край, 1932, 3—4 : 52 (расовый анализ); ibid.: 5—6 : 46 (нерест); Jeffers, Contrib. Canad. biol. fisher., (n. s.), VII, 15—23, 1932 : 7 (пролив Belle-Isle); Dannevig, Journ. Conseil, VII, 1, 1932 : 60 (изменение количеств позвонков при различных условиях развития личинок); Vladykov, Contrib. Canad. biol. fisher., (n. s.), VIII, 1—8, 1933 : 2 (Девисов пролив, питание); Есипов, Тр. Арктич. инст., VII, 1933 : 71, рис. (у берегов Новой Земли до губы Крестовой, сравнение новоземельской трески с мурманской турянкой); Расс, Докл. I сессии Гос. Океаногр. инст., 5, 1933 : 4 (нерест); Graham, Fishery Invest., (2), XIII, 4, (1933), 1934; Пробатов, Мат. научно-пром. обслед. Карск. губы и р. Кары, 1934 : 94 (1 экз. в Карской губе); Расс, Карело-мурм. край, 1934, 3—4 : 57 (нерест у берегов Мурмана); McKenzie, Contrib. Canad. biol. fisher, (n. s.), VIII, 27—31, 1934 : 435 (мечение у берегов Новой Шотландии, Нового Брауншвейга и о. Принца Эдварда); Rollefsen, Rapp. Proces-Verb., LXXXVIII, 1934 : 1; Hansen, Jensen a. Taning, Medd. Komm. Havunders., Fisk., X, 1935 : 3 (мечение трески у берегов Гренландии); Vladykov, Progress Rept., Biol. Board. Canada, 14, 1935 : 3, 2 figs.; Дементьева и Танасийчук, За рыбн. индустр. Севера, 1935, 5 : 40 (расы трески, литература по критике вопроса); Rass, Int. Rev. ges. Hydrob. Hydrogr., 33, 1936 : 251 (нерест); Агапов и Топорков, Пробл. Арктики, 2, 1937 : 107 (у западных берегов Новой Земли); Марти, Тр. Полярн. научно-исслед. инст. морск. рыбн. хоз. и океаногр., 3, 1939 : 3 (сводка сведений и литературы по биологии и расам); Зацепин и Петрова, ibid., 5, 1939 : 2 (питание); Маслов, ibid., 6, 1939 : 3 (промысел); ibid. : 8, 1944 : 3, 50 и след. (биология, промысел); Глебов, Природа, 1941, 4 : 87 (биология); Дубровский, ibid., 6 : 74 (зал. Кротова на Новой Земле в Карском море, 72° с. ш.) .
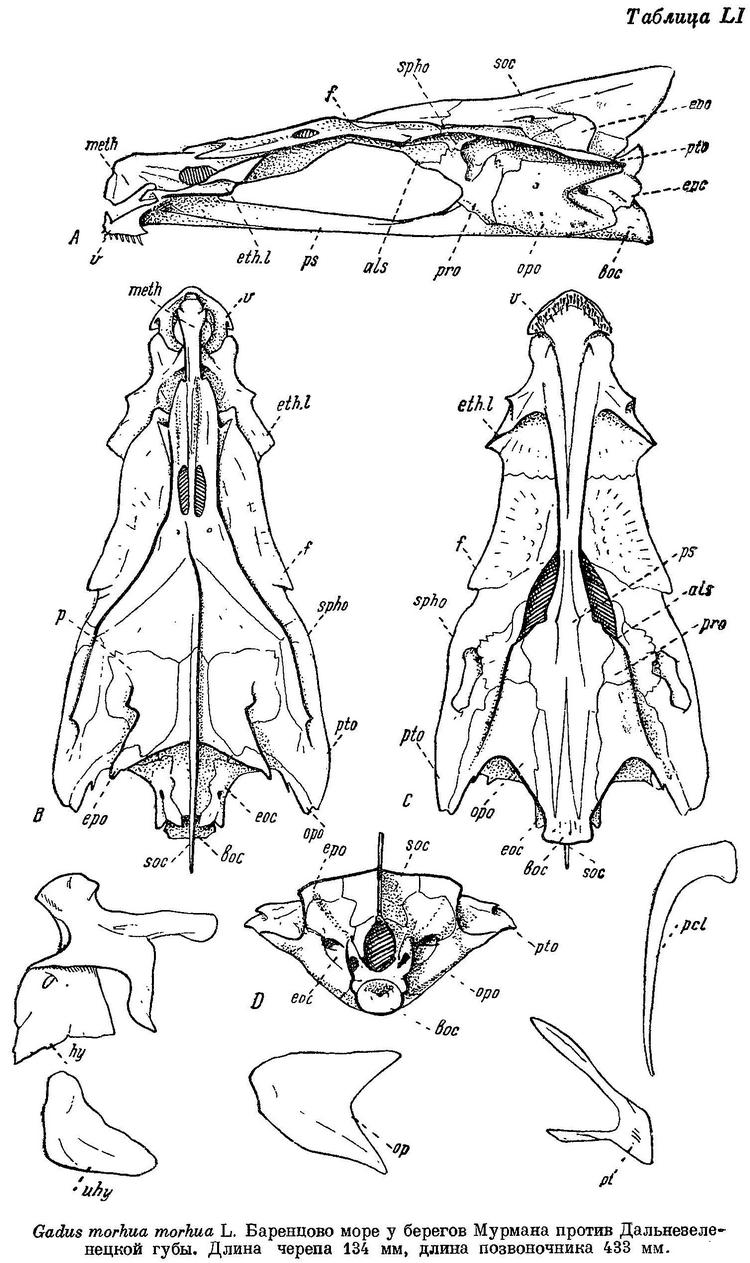
I D 14—15, II D 18—22, III D 17—20, I А 19—23, II А (15) 17—19, sp. br. 21—28, vert. 49—53. Голова небольшая и узкая, длина ее 24.6—26.6% длины тела, межглазничный промежуток 15.8—20.7% длины головы. По данным Дементьевой, средние, основанные на большом количестве экземпляров, колеблются для I D от 14.37 до 14.75, II D от 19.49 до 20.12, III D от 18.76 до 19.39, I А от 21.13 до 22.39, II А от 18.48 до 19.32, количество позвонков от 52.14 до 52.55 (пределы индивидуальных колебаний 50—55, без последнего, считаемого нами). Усик обычно короче продольного диаметра глаза, реже равен ему. Хвостовой стебель удлиненный и невысокий, длина его 11.6—13.1%, высота 4.2—5.1% длины тела. Прижатые к спине лучи 1-го спинного плавника обычно значительно заходят за начало 2-го спинного плавника, реже оканчиваются у начала его или не достигая его. Рожкообразные выросты у переднего конца плавательного пузыря обычно удлиненные и после изгиба к средней линии направлены вперед и на конце загнуты в стороны, у небольших короткие и изогнутые только к средней линии (фиг. 27, а, b).
Перитонеум свинцово-серебристый или свинцово-черный. В процентах к длине тела: антеанальное расстояние 41.2—48.8, антедорсальное расстояние 28.1—31.5, длина грудного плавника 12.1—13.8, длина брюшного плавника 9.1—11.1, высота 1-го спинного плавника 10.0—12.0, высота 1-го анального 8.9—12.3. В процентах к длине головы: длина рыла 33.9—38.4, продольный диаметр глаза 15.5—21.7, длина верхней челюсти 39.2—44.2, длина нижней челюсти 49.0—54.8, длина усика 11.7—15.6. Окраска в высшей степени изменчива. Верхняя часть тела темная, пепельно- или оливково-серая с частыми коричневатыми или сероватыми круглыми пятнами. Пятна имеются и на боках тела и головы, но отсутствуют в передней части головы. Нижняя часть тела беловатая, без пятен. Боковая линия светлее боков тела. Вертикальные плавники серые с темными пятнами, иногда образующими поперечные ряды, грудные и брюшные плавники светлее, брюшной часто одной окраски с брюшком. В зависимости от места обитания окраска трески сильно изменяется. Наиболее известной цветовой вариацией трески является красноватая или оранжевая (aberr. erythrina), называемая у наших берегов турянкой. Окраска эта неустойчива, принимает ее мелкая треска при обитании среди зарослей ляминарий и при перемене места обитания и при выдерживании в аквариуме исчезает. Мелкая треска, держащаяся у берегов, иногда (Jeffers, l. с.) бывает окрашена в темный цвет подобно Gadus morhua maris-albi и Gadus morhua ogac. Размеры наичаще 50—80 см, в среднем около 75 см. максимально до 1.8 м. В июле 1940 г. в Баренцовом море в трал была поймана треска длиной 169 см, весом 40 кг, возраст ее был 24 г.
Распространение. Северная часть Атлантического океана. У берегов Европы от Бискайского залива до Нордкапа; у берегов Сев. Америки от мыса Гаттераса до Гудсонова и Девисова проливов, у юго-западных и юго-восточных берегов Гренландии, вокруг Исландии и, повидимому, к северу от нее почти до Ян-Майен (до 69°13' с. ш., 8°23' в. д., где найдены отолиты), Фарерских, Шетландских, и Оркнейских о-вов, в Северном море; в Баренцовом море по западному побережью Шпицбергена почти до северной; оконечности, на Медвежинском мелководье до широты о. Надежды, в центральной части до 77° с. ш., у Новой Земли до п-ова Адмиралтейства и, весьма вероятно, севернее; в Карском море до Карской губы. Возможно, до устья р. Пясины, где небольшая треска была найдена в желудке омуля (Остроумов, Тр. Полярной ком., 30, 1937 : 53), если это действительно треска. Заходит в пределы распространения своих nationes и близких географически подвидов — в Белое и Балтийское моря и в Гудсонов пролив.
Биология. Атлантическая треска, как это видно из ее распространения, несколько менее теплолюбива, чем сайда и пикша, но значительно более, чем навага. Встречается она при температуре от 0 до 16° С, оптимальными же температурами для нее являются 4—7° С; прибрежные (фиордовые) формы встречаются при несколько более низких температурах; в Баренцовом море в наибольшем количестве встречается при температуре от 1 до 5° С. Живет атлантическая треска, как правило, в воде нормальной океанической солености; случаи захождения ее в пресную воду единичны, но известны неоднократно. Несколько случаев захождения трески в пресную воду известно у берегов. Сев. Америки и Европы. В р. Эльбе поймана треска 30 см длиной (в 15 км выше Гамбурга, рыба эта сильно отличается от типичной трески). 3 трески пойманы в р. Луге в 1 км от устья в начале января 1931 г. Держится обычно на банках или по подводным склонам в пределах континентальной ступени на глубине 10—250, наичаще 40—100 м, предельно до 500 м. Нерест происходит на протяжении почти всего ареала, но основные нерестилища расположены вдоль берегов Норвегии, главным образом на банках у Лофотенских о-вов, где треска во время нереста скопляется в больших количествах. Менее интенсивно нерест происходит в Северном море, у восточных берегов Великобритании и на банках посередине моря, вокруг Ирландии и у западных берегов Великобритании, у Фарерских о-вов, у южных и западных берегов Исландии, у берегов Нью-Фаундленда, у западных берегов Гренландии (до 67° с. ш.), в норвежских фиордах, у берегов Мурмана от Финмаркена до Святого Носа, в некоторые годы в западной части Баренцова моря (у Медвежьего о-ва). Возможно, нерест происходит и у берегов Новой Земли, где непосредственно он наблюдаем не был, но найдена молодь трески. По мнению Расса, молодь эта занесена сюда из других мест. Плодовитость трески чрезвычайно велика, от 570 до 9300 тыс. икринок. Количество икринок у трески размерами 100 см достигает 3—4 млн., у крупных рыб количество икринок достигает 9 млн. и более. Икра в яичнике созревает не вся сразу и выметывается не одновременно. Нерест в основном происходит в пределах или около 100-метровой изобаты, местами в Северном море и вокруг Великобритании на меньших глубинах (20—80 м и даже менее). Нерест не происходит за пределами 200-метровой изобаты, у западных берегов Шотландии и Ирландии — глубже 100 м. Нерест океанической формы трески у Лофотенских о-вов происходит при температуре 4—6° и даже 7° С, однако местные прибрежные (фиордовые) формы у берегов Гренландии и западной части Мурмана мечут икру при значительно более низких температурах (1—2° С и даже при отрицательных от — 0.4° С). Нерест происходит в основном в феврале—апреле, достигая максимальной интенсивности большей частью в марте, в отдельные годы может наступать ранее или же сильно затягиваться, вообще же известны случаи нереста трески почти в любое время года, начиная с осени (августа—сентября) и кончая летом (июнь—июль). Продолжительность развития икры 20—35 дней. Молодь трески встречается повсеместно почти по всему ареалу ее обитания, причем относительно некоторых мест трудно сказать, произошла ли она из икры, выметанной здесь же, или занесена в виде пелагических личинок или мальков. Молодая треска совершает сезонные миграции к берегам и от берегов. Такие же ограниченные миграции, как показали опыты мечения, совершает и взрослая треска в Северном море, у Фарерских о-вов, в Каттегате и Бельтах, у берегов Новой Шотландии, Новой Англии и в ряде других мест — треска фиордовая и мурманская береговая, хотя известны отдельные случаи и отдаленных миграции помеченных в некоторых из этих мест рыб. Треска, мечущая икру у Лофотенскпх о-вов, после нереста, как показали опыты мечения, совершает отдаленные миграции, проникая в струях Североатлантического течения далеко на север, в Баренцово море до берегов Новой Земли. Эта треска составляет обычно основу нашего трескового промысла, так как мурманская местная треска значительно малочисленнее. К берегам Мурмана лофотенская треска подходит в мае—июне, вскоре после появления ее в южной части Баренцова моря на Финмаркенской — Рыбачьей банке. Осенью треска начинает уходить обратно и в ноябре из береговых вод исчезает, продвигаясь далее в западные районы Баренцова моря. На Медвежинской банке она держится круглый год, являясь основой зимнего промысла. Приходящая после нереста в Баренцево море треска отличается меньшей жирностью, чем местная мурманская. Отдаленные миграции, как установлено точно так же многочисленными опытами мечения, совершает крупная треска между берегами Исландии и Гренландии. Численность трески, приходящей к берегам Гренландии, в Баренцево море и у берегов Норвегии, вообще не постоянна, а подвержена периодическим колебаниям. Половой зрелости в разных местах своего ареала достигает не при одном и том же возрасте: в Северном море начинает впервые метать икру на 4—5-м году, по достижении длины в среднем 65—75 см, у Лофотенских о-вов (Rollefsen, l. с.) — в 6—15, наичаще в 10—11 лет, при длине 60—110 см. Предельным возрастом трески следует считать 24 года, а возможно и более. По достижении половой зрелости нерест происходит ежегодно. Пища взрослой трески крайне разнообразна, но основой питания ее является рыба — сельдь, мойва, песчанка, молодь и мелкие тресковые. Кроме того, питается пелагическими Euphausiidae и Hyperiidae и в меньшей степени придонными ракообразными (Schizopoda, Amphipoda, Decapoda). Молодь, ведущая пелагический образ жизни, питается планктоном (Calanus). В центральных и западных районах Баренцова моря с октября по апрель наибольшее значение в питании имеет сельдь. С февраля по июнь в тех же районах и на прибрежных мелководьях питается мойвой. Молодь трески и пикши служит пищей в течение круглого года: в конце весны — в западных промысловых районах; в это же время, а также в начале осени, и с конца декабря и в январе — в центральных районах; осенью — в восточных промысловых районах и на Новоземельском мелководье. Сайка имеет большое значение в питании трески в восточных промысловых районах Баренцова моря, на Новоземельском мелководье и отчасти по северному и южному склонам Гусиной банки в осенне-зимние месяцы. Euphausiidae и Hyperiidae наибольшее значение в питании трески имеют летом. Донные ракообразные и ряд моллюсков и червей служат пищей, в восточных промысловых районах. Главное значение в откорме имеют центральные и восточные промысловые районы, западные — меньше. В западных районах средние индексы наполнения желудков невелики (35—173), в центральных выше (86—173), наиболее высоки в восточных (170— 224). В наших промысловых районах в больших концентрациях треска, по исследованиям, требующим впрочем выяснения причин этого явления, как оказывается, интенсивно не питается. Интенсивное питание происходит в непромысловых районах и в непромысловое время, когда треска держится рассеянно. Стаями треска держится на подходах к кормным районам; во время нереста почти не питается. Рост трески в различных местах ее широкого ареала неодинаков; наиболее медленно растет прибрежная (фиордовая) треска. Пределы колебаний средних размеров половозрелой и близкой к половой прелости атлантической трески из Баренцева моря таковы: 4-годовалые — 35.0—38.4 см, 5-годовалые — 41.8—44.9 см, 6-годовалые — 50.7—55.2 см, 7-годовалые 60,1—64.0 см, 8-годовалые — 69.7—73.0 см, 9-годовалые — 80.6—82.5 см, 10-годовалые — 89.7—91.4 см. Возрастной состав, размеры и уловы атлантической трески не постоянны, а колеблются, находясь в зависимости от численности поколения («урожайности») ее в отдельные годы. Поколения урожайных лет вступают в промысел через 5—7 лет и являются преобладающими в уловах в течение нескольких лет. В Баренцовом море урожайными были поколения 1919, 1929, 1930 и 1933 гг., не урожайным — поколение 1931 г. Содержание жира в печени трески также испытывает существенные колебания из года в год. Так, например, у трески, ловимой с берегов Норвегии, относительный вес печени значительно изменялся в период 1900—1911 гг., будучи минимальным в 1903 г., максимальным в 1900 и 1907 гг. Подобные периодические изменения испытывает жирность (вес печени в процентах к весу тела) и в Баренцовом море. Наибольшее количество жира в печени содержит крупная треска длиной свыше 70 см, жирность которой в среднем колеблется от 4.0 до 9.2%, у более мелкой — от 2.5 до 6%.
Хозяйственное значение имеет очень большое. По величине улова среди тресковых стоит на первом месте. Ежегодные уловы у берегов Европы, Гренландии и Сев. Америки (Нью-Фаундленд) за последние годы превышает 10 млн. ц; до первой мировой войны вылавливалось ежегодно свыше 6 млн. ц. Наш траловый флот в период времени с 1934 по 1938 г. вылавливал ежегодно от 558 (1934 г.) до 1750 (1937 г.) тыс. ц. Кроме того, довольно значительные количества трески вылавливались прибрежным промыслом.
Сравнительные заметки. Атлантическая треска в пределах своего ареала образует несколько форм (рас), ближе еще не изученных и морфологически друг от друга почти не различимых, но характерных неодинаковыми темпом роста и размерами и обособленностью нерестилищ. Наиболее широко распространенной и мощной является форма трески, главные нерестилища которой расположены около Лофотенских о-вов, в небольшом количестве она нерестится и в Мотовском заливе. Характеризуется, она большим количеством позвонков (по Дементьевой и Танасийчук, в среднем 52.5 позвонка), жаберных тычинок (в среднем 24.9) и лучей в плавниках (19.9 во II D). У наших берегов лофотенская треска преобладает в южной части Баренцова моря. В наиболее чистом виде она встречается весной в западных районах (Финмаркенская — Рыбачья банка), в особенности в моменты подхода рыбы. В центральных районах в более чистом виде держится в летние месяцы и поздней осенью, в остальное время здесь встречается в смешанном виде; на Медвежинской банке держится круглый год. Количество ее у наших берегов в разные годы неодинаково, в более теплые годы она приходит сюда в больших количествах, в холодные — в меньших. На встречаемость у берегов Мурмана, кроме пришлой лофотенекой, еще местной, здесь же и размножающейся трески указывалось еще первыми исследователями (Брейтфус и Гебель, l. с.) позднее самостоятельность этой формы трески отрицалась (Есипов, l. c.), но в последние годы получены новые данные (Дементьева и Танасийчук, l. c.), подтверждающие обособленность этой еще ближе не изученной формы. Более подробные сведения о биологии мурманской трески приводит Т. И. Глебов (1941), который также указывает, что среди мурманской трески в последнее время различают две формы.
Мурманская треска нерестится у берегов Мурмана, главным образом в Мотовском заливе, в меньшем количестве и у восточного побережья (до Рынды); в некоторые годы нерест ее происходит в значительных количествах. Характеризуется она меньшими, чем лофотенская треска, количествами позвонков (в среднем 51.8, без последнего считаемого нами позвонка), жаберных тычинок (в среднем 23.7), лучей в плавниках (в среднем II D 19.4), большей жирностью и весом, пониженным темпом роста. Больших миграций не совершает. В небольшом количестве эта треска постоянно встречается у берегов Мурмана, примешиваясь ко всем подходящим с моря косякам. По сообщению Дементьевой, летом и осенью, как показали опыты меченая, мурманская треска продвигается на восток (Гусиная банка и Новоземельское мелководье), где она примешивается к пришлой лофотенской треске. Весной неполовозрелая треска этой формы держится у южных берегов Мотовского залива, у о. Кильдина и к востоку от него; зимой держится разреженно в глубинных частях заливов и на мелководье. В открытой части Баренцова моря в чистом виде не найдена, в некоторых прибрежных районах преобладает над лофотенской. Мурманская треска более холодолюбива, чем лофотенская. Максимум подхода мурманской трески к берегам приходится на гидрологическую зиму, максимум лофотенской — на гидрологическое лето. Мурманская треска, несомненно, представляет самостоятельную, ближе еще не изученную форму (natio) атлантической трески; возможно, что она идентична с беломорской зимней треской. Есть данные, указывающие на существование еще формы трески малопозвонковой и многотычинковой, нерестящейся, повидимому, в северных фиордах Норвегии. Далеко на восток в Баренцово море эта треска не заходит, в некоторые годы является основой летнего промысла на Медвежинской банке, в южной части Баренцова моря входит, повидимому, в состав летних косяков трески и большого значения в уловах здесь не имеет. Форма трески, подобная или даже идентичная мурманской, встречается в Троньемском фиорде; в заливе Св. Лаврентия точно также встречается треска с меньшим количеством позвонков и лучей (J. Schmidt, l. c.). Шпицбергенская треска отличается карликовыми размерами и некоторыми внешними отличиями (Ehrenbaum, l. c.). Вообще в заливах и фиордах атлантическая треска образует ближе не изученные локальные формы с ограниченными миграциями и местным нерестом. Кроме того, неоднородна треска и на протяжении Атлантического океана; в западной часта его она имеет большее количество позвонков и лучей во II D, чем в восточной; у трески, держащейся вокруг Великобритании и Ирландии, количество позвонков и лучей во II D еще меньше. В настоящее время трудно сказать, чем являются все эти перечисленные выше формы трески. Во всех этих случаях необходимо помнить, что в зависимости от условий развития икры и личинок количество позвонков у трески, как показал Dannevig (l. c.), значительно изменяется. Более, резко обособлены географически и лучше изучены формы, которые образует атлантическая треска в Белом и Балтийском морях и в оз. Могильном на о. Кильдене.
*la1. Gadus morhua morhua n. hiemalis Taliev — Беломорская зимняя треска (табл. XIII).
— Gadus callarias hiemalis Талиев, Изв. Отд. прикл. ихт., XI, 2, 1931 : 113, фиг. 2 (Кандалакшский залив).
— Gadus morhua hiemalis Европейцева, Расовый анализ беломорской трески, изд. Ленингр. Гос. унив., 1937 (биометрическая характеристика, питание, нерест, темп роста).
— Gadus callarias Ильин и Певзнер, Сборн., посвящ. Книповичу, 1939 : 320—322 (биометрика).

I D 12—13, II D 18—20, III D 18—20, I A 20—22, II A 17—23, sp. br. 21—22, vert. 52—55. По данным Н. В. Европейцевой, I D 12—16 (14.3), II D 17—21 (19.5), III D 17—21 (19.0), I А 19—25 (21.6), II А 16—21 (18.6), sp. br. 20—26 (23.3), vert. 51—54 (52.3) (без последнего позвонка, считаемого нами). Голова сравнительно короткая и узкая, длина ее 25.2—27.2% длины тела, ширина межглазничного промежутка 14.9—18.5% длины головы. Усик обычно короче продольного диаметра глаза, реже равен ему. Хвостовой стебель удлиненный и низкий, длина его 10.8—13.2% длины тела, высота 4.5—5.0% той же длины. Тело и непарные плавники невысокие. Лучи 1-го спиного плавника в прижатом к спине состоянии обычно заходят за начало 2-го спинного плавника на 1/7—l/3 его длины. Рожкообразные выросты у переднего края плавательного пузыря обычно как у типичной формы. Окраска перитонеума от свинцово-черного до черноватого. В процентах к длине тела: антеанальное расстояние 44.6—49.3, антедорсальное расстояние 27.9—32.1, длина грудного плавника 13.3—14.7, длина брюшного плавника 10.4—11.5, высота 1-го спинного плавника 11.0—13.1, высота 1-го анального 10.0—12.2. В процентах к длине головы: длина рыла 40.5—42.8, продольный диаметр глаза 19.7—23.6, длина верхней челюсти 40.5—42.8, длина нижней челюсти 48.4—52.2, окраска, как у типичной формы. Обычные размеры 20—40 см, наибольшие до 50 см. (8 экз.).
Форма очень близкая, морфологически почти не отличимая от атлантической трески. Отличается лишь биометрически (по средним) длиной грудных и брюшных, 2-го спинного и 1-го анального плавников, более длинным усиком. Краниологически точно так же неотличима. Повидимому, идентична с мурманской прибрежной треской. По сообщению Дементьевой, по ее исследованиям, эта треска по всем признакам ближе всего именно к мурманской прибрежной форме трески. Наиболее существенны отличия от типичной формы в темпе роста и вообще в карликовости этой трески. Б. С. Ильин и В. И. Певзнер, сравнив эту сомнительную форму с экземплярами мурманской трески (из Полярного, Ейна-губы, Урицы и из-под Базаров Рыбачьего пол.), обнаружили различия лишь в длине головы, диаметре глаза и в росте.
Распространение. Белое море, откуда подходит к Кандалакшскому заливу. В Кандалакшском заливе встречается в наибольшем количестве осенью и зимой; летом она отсюда уходит. Подход к Кандалакшскому заливу начинается с июня и позднее — в зависимости от места, так как к местам, расположенным ближе к устью залива, подход наблюдается раньше. В Белом море, повидимому, совершает значительные миграции, возможно выходя даже за пределы его. К Канину Носу, где была ловима треска с характерным для этой формы темпом роста, что отчасти подтверждает высказанные выше соображения об идентичности ее с мурманской прибрежной формой трески.
Биология. Нерестится частично или целиком в Белом море. Известны только 2 близкие к нересту (IV стадии зрелости) самки в возрасте 5 лет, пойманные 11 апреля 1936 г. в губе Педунихе на глубине 6 м. Самец (не совсем типичный) с половыми продуктами в V стадии зрелости был пойман в 1935 г. в губе Педунихе. Летом молодь держится в прибрежной полосе совместно с однолетними Gadus morhua maris-albi, поедая вместе с ними Harpacticus, бентонических Crustacea, личинок креветок, Amphipoda, Brachyura и др. Более крупные рыбы, размерами 15—25 см (2—3-годовалые) поедают преимущественно Nereis, Caprella, затем рыб (сельдь, мойву и др.), Decapoda. Рыбы размерами 25—50 см (4-годовалые и более) питаются преимущественно рыбой — мойвой (а не колюшкой, как Gadus morhua maris-albi), затем Nereis pelagica, Amphipoda и Decapoda. Весной питается пищей, сходной с пищей крупных (25—50 см) Gadus morhua maris-albi. Отличается замедленным по сравнению с мурманской треской и несколько более по сравнению с Gadus morhua maris-albi ростом. По данным обратного расчисления по чешуе, собранной в 1933 г., Н. В. Европейцева дает следующие величины роста и прироста:

Хозяйственное значение сравнительно небольшое, но большее, чем Gadus morhna maris-albi, так как главная масса вылавливаемой в Кандалакшском заливе трески приходится на эту форму. Улов обеих форм трески в Кандалакшском заливе за период с 1895 до 1915 г. не превышал 100 центнеров, в последние годы этот кустарный промысел почти прекратился. Ловят преимущественно женщины, главным образом на удочку (Дексбах, Рыбн. хоз. СССР, 1925, 8—9 : 13).
*lb. Gadus morhua callarias Linne — Балтийская треска (табл. XI).
— Gadus callarias Linne, Syst. nat., ed. X, 1758 : 251 (ex parte: Балтийское море); Pallas, Zoogr. rosso-asiat., III, 1811 : 182 (Балтийское море у берегов Курляндии, Лифляндии, Эстонии).
— Gadus morhua Кесслер, Рыбы Спб. губ., 1864 : 61 (ex parte: иногда у Кронштадта, 1 экз. в Невской губе).
— Gadus callarias Smitt, Scand. fish., I, 1893 : 473 (ex parte: Балтийское море); J. Schmidt, C.-R. Trav. Lab. Carlsberg, XVIII, 6, 1930 : 15 (ex parte: Балтийское море к востоку от о. Борнгольм).
— Gadus morhua callarias Берг, Рыбы пресных вод, II, 1933 : 744 (Балтийское море к востоку от о. Борнгольм до Невской губы, в р. Луге в 1 км от устья).
Распространение, биология. Malmgren, Kritisk Öfvers. Finnl, fisk.-fauna, 1863 : 27 (Ботнический залив до Улеаборга); Möbius u. Heincke, Fische Ostsee, 1883 : 233 (биология); G. Schneider, Acta Soc. Fauna Flora Fennica, XX, 1, 1900 : 24; ibid., XXII, 4, 1901 : 43; ibid., XXII, 2, 1902 : 67 (питание, зараженность паразитами); Sandmann, Rapp. Proces-Verb., V, 1906 : 41, pl. IV (Финский и Ботнический заливы, нерест, размеры); Hessle, Meddel. Kungl. Lantbruksstyrels, 243, (2, 1923), 1923 : 8 (берега Швеции, нерест, питание); 278, (7, 1929), 1929 : 3, 35 (берега Швеции, опыты мечения); Kändler, Kieler Meeresforsch., II, 2, 1938 : 272 (зап. часть моря к востоку от о-ва Борнгольм, весенний и осенний нерест, сравнение весенне- и осенненерестующей трески); Берг, Изв. Всесоюзн. научно-исслед. инст. озерн. и речн. рыбн. хоз., XXIII, 1940 : 23 (Финский зал., размеры); Ziegik, Arch. Hydrob. Rybact., Suwalki, XI, 1938 : 51; Глебов, Рыбн. хоз., 1946, 4—5 : 9 (Балтийское море к югу от о. Готланда, траловый лов).
I D 14—16, II D 16—20, III D 17—20, I А 18—22, II A 18—21, sp. br. 18—23, vert. 53. Количество позвонков только по одному экземпляру; по J. Schmidt'у, 50—56 позвонков. Голова сравнительно короткая и узкая, длина ее 25.4—27.8% длины тела, ширина в межглазничной области 16.2—21.0% длины головы. Усик обычно короче, реже несколько более продольного диаметра глаза. Хвостовой стебель удлиненный и низкий, длина его 9.5—12.4, высота 4.1—4.8% длины тела. Тело и непарные плавники невысокие. Лучи 1-го спинного плавника в прижатом к спине состоянии обычно заходят за начало 2-го спинного плавника, изредка достигают лишь начала его. Плавательный пузырь наичаще несколько более крупных размеров и менее эластичен, чем у типичной формы, рожкообразные выросты у переднего края его у взрослых обычно очень длинные, свернуты в клубочек (фиг. 27, с). В процентах к длине тела: антеанальное расстояние 45.7—48.2, антедорсальное расстояние 29.5—33.3, длина грудного плавника 13.6—14.9, длина брюшного плавника 9.9—12.0, высота 1-го спинного плавника 11.2—13.6, высота 1-го анального 9.4—12.1. В процентах к длине головы: длина рыла 35.4—39.0, продольный диаметр глаза 15.9—20.5, длина верхней челюсти 40.7—44.5, длина нижней челюсти 48.8—52.9, длина усика 13.8—18.6. Окраска оливковая или зелено-серая с частыми сероватыми, коричневатыми или коричневыми пятнами и вообще более пестрая, чем атлантической трески; часто окраска бывает более темной. Обычные размеры 40—50 см, в восточной части моря мельче, 30—45 см. Вес в Финском зал. в среднем 0.5 кг, в исключительном случае 3—4 кг. Встречающиеся иногда в Финском заливе крупные особи длиной почти до 100 см и весом до 7.75 кг относятся, повидимому, к случайно заходящей атлантической треске.
Распространение. Балтийское море к востоку от о. Борнгольма, весьма обычна в средней его части и до середины Финского и Ботнического (прол. Кваркен) заливов, реже встречается в северо-восточной части Ботнического (у Улеаборга) и в восточной части Финского заливов, в последнем даже в Невской губе, но единично.
Биология сходна с биологией прибрежных форм атлантической трески. Как показали опыты мечения, держится локально, совершая очень ограниченные миграции. Все помеченные ловились, по крайней мере в течение года, на расстоянии 1—2 морских миль от места мечения. В середине северной части Балтийского моря концентрируется, повидимому, особенно вокруг глубин; наилучшие уловы обычны на склонах этих глубин или в тех местах у берегов, где близки глубины. К югу от о. Готланд (55°50' с. ш., 19° в. д.) в 1941 г. наибольшие скопления были в придонных слоях воды с температурой 4—5° С. Предпочитает каменистые грунты, на мягких илистых и песчаных грунтах встречается реже. Характерной особенностью этой формы трески является то, что она и живет и размножается при пониженной солености; уже в западной части Балтийского моря, к востоку от о. Борнгольма, соленость снижается до 13—18‰ и даже ниже; в центральной и восточной частях Балтийского моря она еще ниже (около 10—12‰), однако нерест происходит и здесь. Для нереста подходит ближе к берегу, в некоторых местах еще с осени, и держится здесь до мая. В западной части моря нерест начинается с января и заканчивается в мае, наиболее интенсивно происходит в марте. В восточной части моря икрометание происходит в меньшей степени, продолжается и летом; основной период его приходится на более позднее время. В глубокой западной части моря к востоку от о-ва Борнгольма нерест длится с марта по сентябрь включительно и наблюдается два периода его — весенний, растянутый с апреля по июль с наибольшей интенсивностью в июне, и более короткий осенний, заканчивающийся в сентябре, с наибольшей интенсивностью в середине этого месяца. Нерест здесь происходит на склонах глубин, на глубине от 70 до свыше 100 м при придонной температуре воды в сентябре 4.3—4.6° С и содержании солей от 12.9 до 17.8‰. У о-ва Готланд, Аландских о-вов и против Стокгольма нерест с наибольшей интенсивностью происходит в апреле—июне, в Ботническом зал. — в июле и августе. Зрелой становится уже в трехгодовалом возрасте. В средней части Балтийского моря созревает при длине менее 40 см в трех четырехгодовалом возрасте. Количество самок всегда больше количества самцов. Икринки и личинки найдены над глубинами 100—300 м. В Ботническом зал. растет быстрее, чем в более южных частях моря. Питается ракообразными и рыбой; на глубинах пища состоит из Mesidothea entomon, Mysis oculata, Pontoporeia temorata, Lumpenus lampetriformis, сельди, на небольшой глубине — Gammarus locusta, рыбы (сельдь, бельдюга, колюшка, песчанка).
Хозяйственное значение имеет сравнительно небольшое, к западу от пролива Бельт за последние годы вылавливалось около 170 тыс. ц (1936 г.). В частности, в этом году у берегов Дании было выловлено 19.3, Швеции — 59.3, Германии — 45, Польши — 9.3 (в 1932 г. — 38.1), Латвии 29.7 (в 1939 г. — 54.6), Эстонии 6.7 (в 1939 г. — 27.7). Литвы — 6.9 тыс. ц (в 1939 г.) (По опубликованным данным перечисленных стран.). В Финском зал. промыслового значения не имеет. Добывается главным образом ярусами, отчасти удёбным ловом и сетями. У открытых берегов моря лов начинается в марте, в период, подхода к берегам для нереста, и продолжается до зимы. В наибольшем количестве ловится с июня по октябрь. Зимой ловится на больших глубинах в отдалении от берегов.
Сравнительные заметки. Внешне и краниологически почти не отличается от атлантической трески, имея лишь несколько меньшее количество лучей во 2-м спинном плавнике (по J. Schmidt'у, в среднем около 17.5, в то время как у океанической трески среднее количество их обычно свыше 19), меньшее количество жаберных тычинок и меньшие размеры, но существенно отличается более длинными и мягкими, свернутыми в клубочек рожкообразными выростами плавательного пузыря. Есть указания на наличие в Балтийском море формы трески с укороченным черепом и крутым профилем головы.

*lc. Gadus morhua kildinensis Derjugin — Кильдинская треска (табл. XI, LII).
Gadus callarias kildinensis Дерюгин, Тр. Петрогр. общ. ест., LI, 1, 1920 : 26 (оз.. Могильное на о-ве Кильдине); Тр. Петергофск. инст., 2, 1925 : 78 (измерения, биология); Расс, Раб. Мурм. биол. ст., III, 1929 : 25 (измерения 2 экз.); Есипов, Гидробиол. журн. СССР, IX, 4—6, 1930 : 131 (сравнение с атлантической треской).
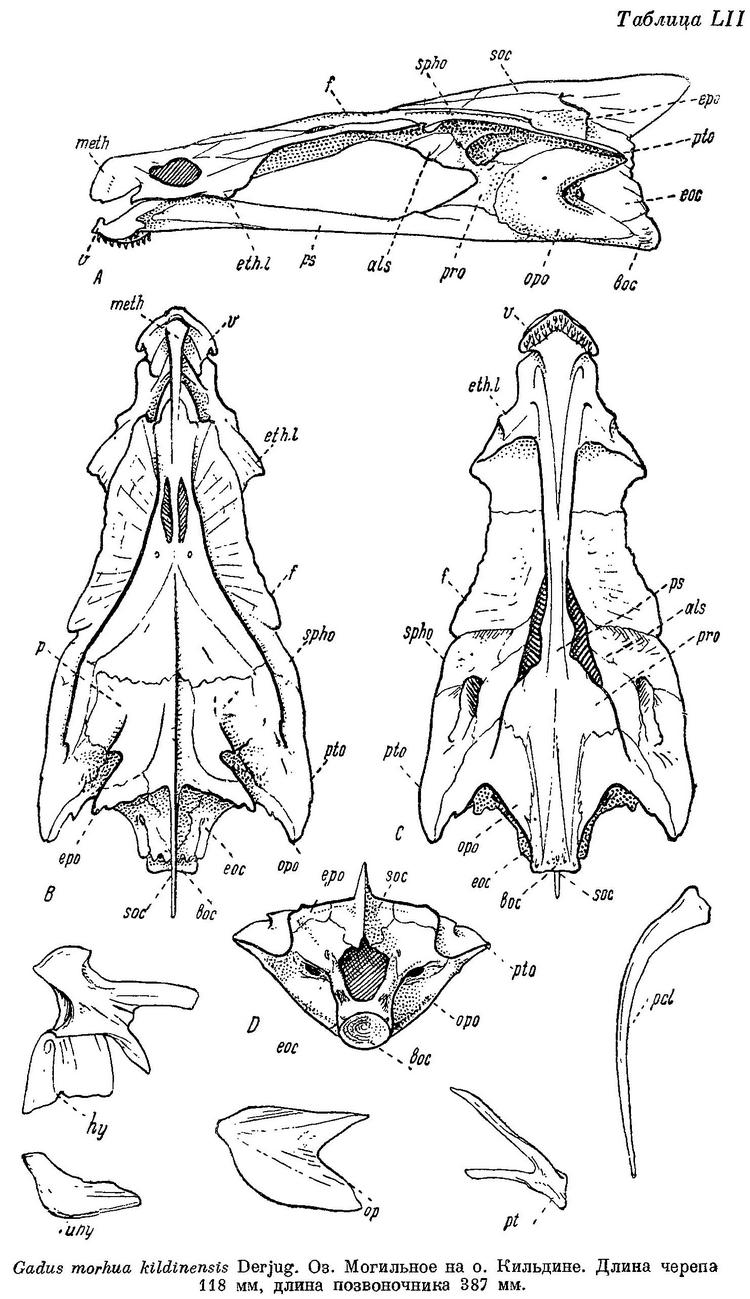
I D 12—14, II D 17—20, III D 17—20, I А 18—20, II А 17—20, sp. br. 20—24, vert. 54. По Есипову (17 экз.), I D 12—14 (12.8), II D 16—20 (18.4), III D 17—20 (18.3), I А 18—22 (20.1), II А 17—20 (18.3), sp. br. 21—24 (22.0), vert. 51—52 (51.8), без последнего позвонка, считаемого нами. Голова небольшая, но довольно широкая, длина ее 24.4—26.9% длины тела, ширина межглазничного промежутка 17.5— 22.0% длины головы. Хвостовой стебель удлиненный, длина его 11.9—13.3%, высота 4.2—4.8% длины тела. Лучи 1-го спинного плавника в прижатом к спине состоянии едва заходят за начало 2-го спинного плавника. Рожкообразные выросты у переднего края плавательного пузыря у взрослых обычно очень длинные, чаще всего свернуты в клубочек (фиг. 27, g, h, i). Перитонеум свинцово-серебристый или свинцово-черный. В процентах в длине тела: антеанальное расстояние 44.1—49.1, антедорсальное расстояние 27.7—31.5, длина грудного плавника 13.2—14.4, длина брюшного плавника 9.4—11.2, высота 1-го спинного плавника 11.2—12.0, высота 1-го анального 10.7—11.5. В процентах к длине головы: длина рыла 34.0—37.7, продольный диаметр глаза 16.0—20.3, длина верхней челюсти 40.0—42.1, длина нижней челюсти 48.8—50.3, длина усика 14.5—17.7. Окраска темная и очень пестрая. Основной цвет верхней части тела коричневатый с часто расположенными темнокоричневыми круглыми пятнами; такие же пятна имеются и на спинных и хвостовом плавниках; брюшные и анальные плавники темные с мелкими черными точками, грудные несколько темнее. Известные наибольшие размеры 79.7 см. (8 экз.).
Paспространение. Оз. Могильное на о. Кильдине к западу от входа в Кольский залив.
Биология этой формы крайне своеобразна. Живет в небольшом замкнутом озере с максимальной глубиной около 17.5 м, с соленостью придонных слоев воды (ниже 10 м) от 22 до 32‰ и сильно опресненными, почти пресными поверхностными слоями. Держится в придонных слоях воды, но известны случаи поимки на глубине 1.5—2 м, т. е. в воде с соленостью около 1—5‰. Большой интерес представляет ближе еще не изученный нерест этой трески, так как пелагическая икра ее должна находиться или в опресненных поверхностных слоях воды или на глубине метров 10. Самки со зрелой икрой и самцы с развитыми молоками были находимы в середине февраля; очевидно, нерест происходит в марте. Мальки в 7.3 и 9.5 мм длиной пойманы в конце июня. 140 экз. трески длиной не более 7 см были пойманы неводом в начале сентября. Питается ракообразными. Рост этой формы трески, повидимому, близок к росту атлантической; в 5 лет она имеет размеры в среднем около 60.7 см, в 6 лет — 65.1 см.
Хозяйственного значения не имеет, так как запасы ее в небольшом озере незначительны.
Сравнительные заметки. Произошла эта форма трески, несомненно, от Gadus morhua morhua, причем, судя по одному черепу и величине межглазничного промежутка у 8 измеренных рыб, череп ее несколько более широк, чем у атлантической трески. Значительно сильнее отличается, кильдинская треска по форме черепа и по ряду других признаков от широкоголовых форм (Gadus morhua maris-albi, Gadus morhua ogac, Gadus morhua macrocephalus).
Рыбы пресных вод СССР и сопредельных стран. Л.С.Берг 1948 г.
1. Gadus morhua Linne. — Атлантическая треска
Gadus morhua Linne, Syst. nat., ed. X, 1758, p. 252 (Атлантический океан у берегов Европы).
Gadus callarias Linne, 1. с. (частью: Атлантический океан у берегов Европы). — Smitt, Scand. fish., I, 1892, p. 472 (частью: Аглантический океан у берегов Европы и Сев. Америки). — Михин, Изв. Отд. прикл. ихтиол., III, вып. 2, 1925, стр. 191 (Мурман). — Суворов и Чудинов, Изв. Отд. прикл. ихтиол., VI, вып. 2, 1927, стр. 233 (турянка, morpha erythrina; Мал. Оленье на Мурмане). — J. Schmidt, С. R. trav. lab. Carlsberg, XVIII, № 6, 1930 (частью: Атлантический океан у берегов Европы и Америки). — Талиев, Изв. Ихтиол. инст., XI, вып. 2, 1931, стр. 137 (Белое море у Поньгомы). — Есипов, Тр. Инст. по изуч. Севера, вып. 48, 1931, стр. 75—91 (Баренцево море на меридиане о. Кильдина, 50 экз.; турянка из губы Порчниха, биометрика, 114 экз.; морфологически турянка не отличается от трески из Баренцова моря на меридиане Кильдина), цветн. рис. турянки. — Дементьева и др., Расовый состав трески Баренцова моря. Доклады 1-й сессии Океаногр. инст., № 2, 1932. — Ряд работ по биологии трески зап. Мурмана в Сборн. научно-пром. работ на Мурмане. М.—Л., 1932. — Сынкова, Изв. Ихтиол. инст., XIII, вып. 2, 1932, стр. 89 (Новая Земля; биометрика, возраст). — Гурвич, там же, стр. 101 (Новая Земля на север до Крестовой губы).
новоземельская треска Есипов, Тр. Аркт. инст., VII, 1933, стр. 71—81, рис. (биометрика).
Gadus morhua Берг, Рыбы пресных вод СССР, II, 1933, стр. 744, рис. — Есипов, Пром. рыбы Баренц. м., Л., 1937, стр. 76.
треска Ю. Марти, Тр. Полярн. инст. рыбн. хоз., III, 1939, стр. 3—37 (Мурман, биология). — А. Дубровский, Природа, 1941, № 6, стр. 74 (Карское море у берегов Новой Земли: зал. Кротова под 72° с. ш.; Маточкин шар, 29 X 1939, длина 1 м). — Маслов, Тр. Полярн. инст. рыбн. хоз., VIII, 1944, стр. 3—184 (Баренцово море; биология).
Gadus morhua morhua Световидов, Трескообразные, 1948, стр. 165, табл. Х, фиг. 2 (рыба), табл. LI (череп), табл. LXXI, фиг. 2 (позвоночник).
Формула по данным Т. Ф. Дементьевой, на основании изучения 2000 экз. из Баренцова моря и Мотовского залива. Средние для разных проб: I D 13.8—14.8, II D 19.2—20.3, III D 18.8—19.4, I А 21.3—22.0, II А 18.5—19.3, жаберных тычинок на 1-й дуге в Баренцевом море 24.1—25.0, в Мотовском заливе (156 экз.) 23.6, позвонков (считая с уростилем) для 1500 экз. из Баренцова моря 51—55 (55), средн. 53.3—53.5, для 422 экз. из Мотовского залива 51—55, средн. 53.1—53.2. Среднее количество лучей во II D больше 18.0. Длина (абс.) в среднем около 75 см. В июле 1940 г. в Баренцевом море поймана треска весом 40 кг, длиною 169 см, возрастом 24 года (Рыбн. хоз., 1940, № 10, стр. 29). Мелкая треска, живущая среди зарослей ламинарий и называемая турянкой, принимает красноватую окраску (aberr. erythrina).
Берега Европы от северной оконечности зап. Шпицбергена и п-ова Адмиралтейства на северном острове Новой Земли и западной части Карского моря (Летом 1932 г. один экземпляр трески обнаружен А Н. Пробатовым в Карской губе (Матер. по обслед. Карской губы. М., 1934, стр. 94) и в довольно большом количестве в Югорском шаре. В 1940 г. на западном берегу Карского моря треска ловилась на север вплоть до становища Архангельского (севернее Маточкина шара; И. Д. Агапов).) до Бискайского залива; западная часть Балтийского моря, заходит в Белое море; берега Сев. Америки от Гудсонова пролива и Гренландии до Виргинии. Отмечены, как исключение, случаи захождения этой трески в пресную воду в Дании (Колдинг-о, Оденсе-фьорд). Шнакенбек упоминает о поимке 11 декабря 1930 г. трески в Эльбе, в 15 км выше Гамбурга (Јool. Anz., v. 94, 1931, p. 17—32), но экземпляр этот (длиною 30 см), согласно описанию, настолько отличается от обычной трески, что признать его за Gadus morhua невозможно.
Образует ряд форм.
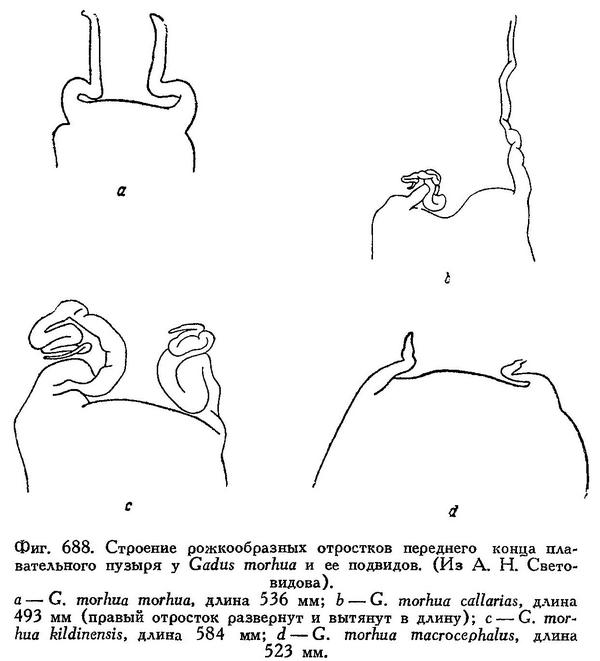
la. Gadus morhua morhua natio hiemalis Taliev. — Беломорская зимняя треска
(Gadus callarias hiemalis Талиев, Изв. Ихтиол. инст., XI, вып. 2, 1931, стр. 113, рис. 2; Кандалакшский залив Белого моря, зимою. — Gadus morhua hiemalis Европейцева, Расовый анализ беломорской трески, Л., 1937, стр. 4 и сл. — Gadus callarias Б. Ильин и Певзнер, Сборн., посвящ. Книповичу, М., 1939, стр. 321—322; Gadus callarias hiemalis есть синоним Gadus callarias. — Gadus morhua morhua n. hiemalis Световидов, Трескообразные, 1948, стр. 171, табл. XIII, фиг. 1).
Форма, с трудом отличимая от мурманской прибрежной трески (турянки). Отличается главным образом меньшей величиной (абс. длина в среднем 30 см, но попадаются особи до 50 см), а также более медленным темпом роста. Жаберных тычинок на 1-й жаберной дуге 20—26, средн. у 151 экз. 23.3. Позвонков 52—55, средн. у 136 экз. 53.3. Белое море, зимою и осенью в Кандалакшском заливе. Возможно (Европейцева), что эта же треска попадается у Канина носа.
lb. Gadus morhua callarias Linne
(Gadus callarias Linne, 1. c.; частью: Балтийское море. — Pallas, Zoogr. rosso-asiat., III, 1811, p. 182; Балтийское море у берегов Курляндии, Лифляндии, Эстонии. — Gadus morhua Кесслер, Рыбы СПб. губ., 1864, стр. 61; частью: на восток доходит иногда до Кронштадта; экземпляр в 23 см добыт в Невской губе. — Polish Cod Ziecik, Archiw. hydrobiol. i rybactwa, XI, Suwalki, 1938, p. 51—69; Данцигский залив, биометрика, 100 экз.; II D 15—19 (20), средн. 16.9; Балтийское море у берегов Дании, биометрика, 100 экз., II D 16—20, средн. 17.9. — Gadus morhua callarias Берг, Рыбы пресных вод СССР, II, 1933, стр. 744, Балтийское море; Изв. Инст. озерн. и речн. рыбн. хоз., XXIII, 1940, стр. 23; Финский залив на восток до Кронштадта, Копорская и Лужская губы. — Световидов, Трескообразные, 1948, стр. 173, табл. XI, фиг. 1, цельная рыба и фиг. 27с на стр. 163, рожкообразные выросты плавательного пузыря; Балтийское море).
Балтийская треска. (При устье Наровы — пикша (неправильно).) Во II D в среднем меньшее количество лучей, чем у атлантической трески: 16.9—17.9 (пределы 14—22). Среднее количество позвонков 52.5—53.4 [пределы 51—55 (56)]. Жаберных тычинок меньше, 18—23. Рожкообразные выросты плавательного пузыря у взрослых обычно длиннее, чем у атлантической трески, и свернуты в клубочек. Окраска более пестрая, чем у атлантической трески. Длина (абс.) обычно всего 40—50 см, в Финском заливе 30—45 см (вес в Финском заливе в среднем 0.5 кг, как исключение, 3—4 кг). (О крупных экземплярах, возможно принадлежащих к типичной антлантической треске, см.: Берг, 1940, стр. 23—24). Балтийское море к востоку примерно от о. Борнгольма (см.: J. Schmidt, 1. с., р. 15) и вплоть до Невской губы. В начале января 1931 г. в р. Луге, в районе дер. Остров около 1 км выше устья, были пойманы три крупных трески; одна из них длиной 753 мм (абс.) доставлена в Институт озерного и речного рыбного хозяйства; формула этого экземпляра: I D 15, II D 19, III D 18, I А 17, II А 18. (А. Н. Световидов (1948, стр. 174) высказывает предположение, не принадлежат ли эти крупные особи к случайно зашедшей атлантической треске.) Вообще же в восточной части Финского залива треска встречается очень редко.
lc. Gadus morhua kildinensis Derjugin
(Gadus callarias kildinensis Дерюгин, Тр. Петрогр. общ. ест., т. 51, вып. 1, 1920, стр. 27; оз. Могильное на Кильдине; Тр. Петергофск. инст., № 2, 1925, стр. 78—82, табл. — Есипов, Гидробиол. журн., IX, № 4—6, 1930, стр. 131. — Gadus morhua kildinensis Световидов, Трескообразные, 1948, стр. 175, табл. XI, фиг. 2, табл. LII, череп).
Кильдинская треска. Жаб. тыч. 20—24, средн. 22, т. е. меньше, чем у океанической. Антеанальное расстояние в среднем 45% длины всего тела, т. е. меньше, чем у океанической. Рожкообразные выросты плавательного пузыря у взрослых обычно очень длинные, обычно свернуты в клубочек. Окраска темная, очень пестрая. Длина до 70 см (шестилетки). Оз. Могильное на о. Кильдине на зап. Мурмане. Озеро это замкнутое, поверхностные слои воды имеют соленость 1—5‰, придонные до 32‰. Нерест, повидимому, в марте.
1d. Gadus morhua maris-albi Derjugin
(Gadus callarias maris-albi Дерюгин, Тр. Петрогр. общ. ест., т. 51, вып. 1, 1920, стр. 27; Белое море. — Михин, Изв. Отд. прикл. ихтиол., III, вып. 2, 1925, стр. 191; биометрика. — Талиев, там же, XI, вып. 2, 1931, стр. 106, рис. 3. — Gadus morhua maris-albi Берг, Рыбы пресных вод СССР, II, 1933, стр. 746, рис. 717. — Европейцева, Расовый состав беломорской трески, Л., 1937, стр. 7 и сл.; биометрика, биология. — Gadus maris-albi Б. Ильин и Певзнер, Сборн., посвящ. Книповичу, М., 1939, стр. 318—320; пертуй, биометрика, темп роста. — Gadus morhua maris-albi Световидов, Трескообразные, 1948, стр. 176, табл. XII, фиг. 1, табл. LIII, череп; Кандалакшский залив, Соловецкие остроща).
Широкоголовая беломорская треска. Эта треска относится к группе тихоокеанских (Gadus morhua macrocephalus): она имеет широкую голову, ширина межглазничного промежутка 18.4—24.1% длины головы (Световидов). Жаберных тычинок у 184 экз. (18) 19—24, средн. 21.4. Позвонков у 183 экз. 50—54, средн. 52.2. Средняя длины (абс.) разных проб от 213 до 289 мм, но попадаются особи до 60 см (Европейцева). Белое море, преимущественно в Кандалакшском заливе, также у Соловецких островов. Нерест в марте—апреле и позже.
le. Gadus morhua macrocephalus Tilesius
[Gadus macrocephalus Tilesius 1810, Берингово море. — Gadus morhua macrocephalus Андрияшев, Исследования морей СССР, вып. 25, 1937, стр. 350; Берингово море на север до бухты Провидения, длина до 1 м. — Полутов, Рыбн. хоз., 1946, № 6, стр. 37; Авачинская губа. — Световидов, Трескообразные, 1948, стр. 179, табл. XIII, фиг. 2, табл. LV, череп. — треска Авачинского залива Полутов, Изв. Тихоокеанск. инст. рыбн. хоз., XXVIII, 1948, стр. 103; биология. — треска Петрова-Тычкова, там же, стр. 127 (зап. Камчатка; биометрика)].
Тихоокеанская треска. I D 12—16, средн. 14.3; II D 14—21 (22), средн. 17.8, III D 16—21, средн. 18.3, I А (16) 17—23, средн. 20.1, II А 16—21 (22), средн. 18.4; жаб. тыч. 17—24, средн. 19.7; позвонков без последнего (49) 50—55, средн. 53.1 (данные М. А. Петровой-Тычковой по 222—228 экз. с западного берега Камчатки). Голова широкая: ширина межглазничного промежутка 20.3—25.3% длины головы. Голова большая: длина ее 26.7—28.6% длины всего тела. Позвонков 51—56, средн. 54.2. Рожкообразные отростки плавательного пузыря короткие, загнутые к серединной линии. Хвостовой стебель сравнительно низкий, 3.5—4.5% длины тела. Мельче атлантической, особенно мелка треска Японского моря. В общем средняя (абс.) длина от 45 до 90 см. Северная часть Тихого океана на север до бухты Провидения, на юг до Порт-Артура и зал. Тояма-ван на Хондо, а по американскому побережью до берегов Орегона.
Очень близка к тихоокеанской треске Gadus morhua ogac Richardson 1836 (см. Световидов, 1948, стр. 178), которая встречается у берегов зап. Гренландии до 73.5° с. ш. и в Гудсоновом заливе. Это мелкая прибрежная форма, аналогичная беломорской Gadus morhua maris-albi.