(Clupeidae Cuvier, 1816) Семейство Клупеиды, Сельдевые, Family Clupeidae Cuvier, 1816 (Herrings, Shads, Sardines, Menhadens) 63 рода 205 видов
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
Семейство Сельдевые (Clupeidae)
Жирового плавника нет. Половой диморфизм чаще всего не выражен в период нереста. Пелагические морские, проходные, редко пресноводные и солоноватоводные формы. Семейство объединяет около 200 видов, множество подвидов и форм. В наших водах наиболее обычны представители 20 видов.
Декоративное рыбоводство. А.М.Кочетов 1991 г.
Семейство Сельдевые (Clupeidae)
Представители этого семейства с детства хорошо знакомы каждому человеку с гастрономической стороны. Семейство состоит из 6—7 подсемейств с примерно 50 родами и 190 видами. В промысловом отношении это одно из трех важнейших семейств мировой ихтиофауны. У входящих в него представителей рыло не выступает вперед, рот конечный или полуверхний умеренной величины. Окраска, как правило, серебристая с зеленовато-синей спинкой. Брюшной киль зачастую состоит из зазубренных чешуек, зубы на челюстях слабые или отсутствуют вовсе, а плавательный пузырь соединен специальным каналом с желудком. Есть нижние и верхние межмышечные косточки, нет боковой линии (иногда 2—5 чешуй), голова голая.
Наибольшим разнообразием отличаются субтропические и тропические морские виды. Держатся они большими косяками в толще воды; предельный размер 80 см, чаще 20—40 см, самцы мельче, ярче и стройнее самок. Созревают производители в возрасте 2—6 лет. Пелагофилы нередко совершают многокилометровые нерестовые миграции, склонны к ночным икрометаниям. Плодовитость колеблется от сотен до сотен тысяч (2—3 порции) икринок диаметром около 1,5 мм (после набухания до 4 мм). Инкубационный период у тепловодных видов равен 10—24 ч (при температуре воды 22 °С). Корм молоди — микроводоросли, коловратки, науплии циклопа, артемии и т. п. Основу питания взрослых рыб составляет планктон. Из перспективных с точки зрения аквариумного рыбоводства тропических сельдей нужно отметить амазонских шипоносых сардинок (Rhinosardina), аргентино-уругвайских манфудий (Ramnogaster), индийских корик (Corica), бирманских гудузий (Gudusia), центральноамериканских доросом (Dorosoma) и новогвинейских флювиалоз (Fluvialosa). К числу необычных черт следует отнести отсутствие спинного и брюшных плавников у длиннохвостой индокитайской раконды (Raconda russelliana), сильный брюшной киль у гайано-бразильского пилобрюха (Pristigaster cayanus), нижний или полунижний рот, мускулистый зобовидный желудок, а также клейкую икру (диаметром 0,75 мм) у вегетарианцев — доросом. Условия содержания: dH do 30°, рН 7,2—8,5, t 15—25 °С, аэрация, фильтрация, подмена воды, аквариум объемом не менее 100 л на стаю рыб мелких видов, для крупных рыб нужен аквариум объемом в 500 л.
В фауне СССР семейство представлено 6 родами и 14 видами. Тело рыб покрыто чешуей (брюшко с килеватыми чешуйками), голова голая, рот обычно конечный, в боковой линии 2—5 чешуй, хвост сильно выемчатый. При содержании рыб воду лучше подсаливать (5—18 ‰).
Семейство Конготриссовые (Congothrissidae)
Семейство было описано Максом Полом в 1964 г. От всех сельдевидных оно отличается наличием только четырех лучей жаберной перепонки. Окраска оливково-серая, полупрозрачная, с блестящей продольной полоской. Созревают они в 8-12 месяцев. Пелагофилы. Плодовитость до 1000 икринок. Инкубационный период длится 15 ч (при температуре воды 24 °С). Корм - микропланктон. Рыбы нежные. Условия содержания: dH 2-12°, рН 6,0-7,5, t 20-26°С, аэрация, фильтрация, подмена воды, аквариум объемом не менее 50 л.
Конготрисса Госсе - Congothrissa gossei (Poll, 1964), Заир, 8 см. Сходную биологию имеют танганьикские столотриссы и лимпотриссы (Stolothrissa, Limnothrissa), заирские микротриссы и одаксотриссы (Mickrothrissa, Odaxothrissa), a также тумбайские наннотриссы (Nannothrissa parva, N. stewarti); размер 6—15 см.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Семейство Сельдевые (Clupeidae)
Сельдевые рыбы имеют сжатое с боков
или вальковатое тело, обычно
серебристое, с темно-синей или зеленоватой
спиной. Спинной плавник один, обычно в
средней части спины, грудные
расположены у нижнего края тела, брюшные —
в средней трети брюха (иногда
отсутствуют), хвостовой плавник выемчатый.
Очень характерно отсутствие прободенных чешуи боковой линии на теле,
бывающих только в числе 2—5 сразу за
головой. Вдоль средней линии брюха у
многих тянется киль из приостренных
чешуи. Зубы на челюстях слабые или
отсутствуют. Плавательный пузырь
соединяется каналом с желудком, а от
переднего конца пузыря отходят два отростка,
проникающие в ушные капсулы черепа.
Есть верхние и нижние межмышечные
косточки.
Сельдевые — стайные планктоноядные
рыбы; большая часть видов морские, часть
проходные, немногие — пресноводные.
Широко распространены от субантарктики до Арктики, но количество родов и
видов велико в тропиках, убывает в
умеренных водах, а в холодных водах
распространены единичные виды. Большей
частью это мелкие и некрупные рыбы,
менее 35—45 см, только немногие
проходные сельди могут достигать длины 75 см.
Всего насчитывается около 50 родов и 190
видов сельдевых. Это семейство дает около
20% мирового улова рыб, занимая по
величине улова, наряду с анчоусовыми,
первое место среди семейств рыб.
В этом большом и важном семействе
различают 6—7 подсемейств, частью
принимаемых некоторыми учеными в качестве
особых семейств.
Подсемейство Сельди-круглобрюшки (Dussumierinae)
Сельди-круглобрюшки отличаются от
других сельдей тем, что брюхо у них
закругленное и вдоль его средней линии
нет килевых чешуи. Рот небольшой,
конечный. Челюсти, нёбо и язык усажены
мелкими многочисленными зубами. В этой
группе насчитывают 7 родов с 10 видами,
распространенными в тропических и
субтропических водах Тихого, Индийского
и западной части Атлантического
океанов. Среди сельдей-круглобрюшек
различают две группы форм (родов): более
крупных многопозвонковых (48—56
позвонков) рыб, достигающих длины 15—35 см (Dussumieria, Etrumeus), и более
мелких малопозвонковых (30—46
позвонков) рыб, 5—11 см длины (Spratelloides,
Jenkinsia, Echirava, Sauvagella, Gilchristella).
Виды остальных родов круглобрюхих
сельдей — мелкие селедочки, живущие
в бухтах и эстуариях, у берегов
Восточной Африки, Мадагаскара и Индии.
Подсемейство Шпротоподобные, или Обыкновенные сельди (Glupeinae)
Шпротоподобные, или обыкновенные
сельди, представляют собой группу
важнейших для человека сельдевых рыб,
включающую северных морских сельдей,
сардин, сардинелл, шпротов, тюлек и
другие роды. Сюда входят 12 родов с 40—45
видами.
Виды трех родов — Морские сельди
(Clupea), Шпроты (Sprattus) и
аргентинские сельди — Мандуфии (Ramnogaster)—
распространены в умеренных и холодных
водах северного и южного полушарий;
Сардины-Пильчарды (Sardina), Сардины-Сардинопсы (Sardinops) и Тюльки
(Clupeonella) — в умеренно теплых морях;
Сардинеллы (Sardinella), Сардинки и
Селедочки (Harengula, Herclotsichthys), Maчуэлы (Opisthonema) и остальные (Lile,
Rhinosardinia) — в тропических водах.
Сардинами называют виды трех родов
морских сельдевых рыб — сардину-пильчарда (Sardina), сардину-сардинопса
(Sardinops) и сардинелл (Sardinella). Для
этих трех родов характерны удлиненные,
выступающие в виде лопасти два задних
луча анального плавника и наличие двух
удлиненных чешуи — «крылышек» — при
основании хвостового плавника. Кроме
того, у сардины-пильчарда и сардинопсов
на жаберной крышке имеются радиально
расходящиеся бороздки. Настоящие
сардины (пильчард и сардинопсы)
распространены в умеренно теплых и
субтропических морях, сардинеллы — в
тропических и отчасти субтропических водах.
Сардины достигают длины 30—35 см,
в промысловых уловах обычно бывают
13—22 см длины.
Все сардины — морские стайные рыбы,
живущие в верхних слоях воды; питаются
планктоном, выметывают плавучую
икру. Икринки сардин имеют большое кругожелтковое пространство, а в желтке
есть маленькая жировая капля. Сардины
имеют большое практическое значение,
замещая в теплых водах морских сельдей.
Европейская сардина (пильчард) и сардинопсы распространены в умеренно
теплых и субтропических морях северного и
южного полушарий, главным образом при
температуре от 7 — 10 до 20°С, отсутствуя
только в атлантических водах Америки.
Сардинеллы, напротив, распространены
в тропических водах Азии, Африки,
Восточной Америки при температуре воды
от 15 до 30°С, заходят в умеренно
теплые и субтропические моря Южной
Европы, преимущественно при температуре
воды выше 20°С, но отсутствуют в
тихоокеанских водах Америки.
Селедочками и сардинками называют
мелких, до 15—20 см длины, тропических
сельдевых рыб со сжатым с боков
серебристым телом и чешуйным килем на брюхе.
Они населяют прибрежные воды индо-западнотихоокеанской биогеографической
области и Центральной Америки. Нет их
у восточных берегов Атлантического
океана. По строению эти рыбки близки к
сардинеллам. На переднем крае плечевого
пояса под жаберной крышкой у них также
имеются две выступающие вперед
закругленные лопасти. Последние два луча
анального плавника слегка удлинены, не
образуя, однако, выступающей лопасти.
Икринки у них, как и у сардин,
плавучие, с большим кругожелтковым
пространством, с маленькой жировой каплей
в желтке. В отличие от сардин у них нет
удлиненных чешуи при основании
хвостового плавника. Тело у них сжато с
боков, серебристое; позвонков 40—45.
Подсемейство Гологлазые сельди, или Сельди-гологлазки (Pellonulinae)
Подсемейство содержит 14 родов и
свыше 20 видов тропических,
преимущественно пресноводных сельдевых рыб Америки
(8 родов), Индо-Малайского архипелага,
отчасти Индии и Австралии. Жирового
века на глазах у представителей этого
подсемейства нет или оно едва развито,
брюхо обычно сжатое с боков, рот
маленький. У некоторых видов австралийских
родов (Potamalosa, Hyperlophus) на спине
между затылком и спинным плавником
имеется зубчатый киль из ряда щитков
(чешуи). Большинство видов этой
группы — мелкие рыбки, менее 10 см длины.
Подсемейство Пузанковые сельди (Alosinae)
Подсемейство содержит самых крупных
по размерам сельдевых рыб. Большинство
видов этой группы анадромные проходные, часть солоноватоводные, некоторые
пресноводные. В этой группе сельдевых
рыб 4 рода с 21 видом, обитающие в
умеренно теплых и в меньшей мере
субтропических и тропических водах северного
полушария. Пузанковые сельди имеют
сжатое с боков брюхо с шиповатым чешуйным килем вдоль его медиальной линии;
у них большой рот, задний конец верхней
челюсти заходит за вертикаль середины
глаза; на глазах есть жировые веки.
Сюда относятся алозы, гильзы и гудузии.
Алозы распространены в умеренно теплых
прибрежных морских, солоноватых и
пресных водах Восточной Америки и Европы;
гильзы и гудузии живут у берегов и
частично в пресных водах Восточной Африки,
Южной и Юго-Восточной Азии.
К подсемейству пузанковых сельдей
обычно относят также особую группу
сельдевых рыб, близких к американской
менхэден (Brevoortia). По-видимому,
правильнее их выделять в особую группу или
подсемейство гребнечешуйных
сельдей, относя сюда американских
менхэден, мачету и западноафриканскую
бонгу.
Подсемейство Гребнечешуйные сельди (Brevoortiinae)
Отличаются от всех других сельдевых
чешуей с гребенчатым задним краем и двумя
рядами увеличенных чешуи, или щитков,
вдоль средней линии спины, от затылка до
начала спинного плавника. Характерно
для них также наличие 7 лучей в брюшных
плавниках. Они близки к пузанковым
сельдям по форме сжатого с боков
высокого тела, с зубчатым чешуйным килем
вдоль брюха, по наличию медиальной
вырезки в верхней челюсти, по
отсутствию зубов на челюстях у взрослых
особей.
По строению икринок менхэден
отличаются от алоз, а близки к сардинам:
икринки у них содержат жировую каплю
в желтке и являются пелагическими, а не
полупелагическими. В отличие от
пузанковых сельдей гребнечешуйные —
морские рыбы, живущие и размножающиеся
в море при солености не ниже 20‰.
К гребнечешуйным сельдям относятся три
рода: менхэден, очень близкая к ней мачета и бонга.
Подсемейство Тупорылые, или Зобатые сельди (Dorosomatinae)
Тупорылые, или зобатые, сельди,
имеющие короткое, высокое, сжатое с боков
тело, с брюшным пильчатым килем из
чешуи, представляют своеобразную группу.
В отличие от всех прочих сельдевых
рыло у них почти всегда выступающее,
тупо закругленное; рот маленький,
нижний или полунижний; желудок короткий,
мускулистый, напоминающий зоб у птиц.
Анальный плавник довольно длинный, от
18—20 до 28 лучей; брюшные плавники
расположены под началом спинного или
ближе спинного к переднему концу тела,
в них 8 лучей. Почти у всех видов имеется
темное «плечевое» пятно сбоку, позади
верха жаберной крышки; у многих, кроме
того, имеется 6—8 узких темных продольных полос вдоль боков. У большинства
родов и видов последний (задний) луч
спинного плавника вытянут в длинную
нить; только у видов двух родов (Anodontostoma, Gonialosa) он не вытянут. Это
илоядные и питающиеся фитопланктоном
рыбы заливов, эстуариев, рек тропических
и отчасти субтропических широт, не
представляющие большой пищевой ценности
вследствие костлявости. Тем не менее
во многих районах их заготовляют в
пищу, преимущественно в сушено-вяленом
виде и в виде консервов. Всего в этой
группе 7 родов с 20—22 видами.
Тупорылые сельди (или сельди-тупорылки)
распространены в водах Северной и
Центральной Америки (род Dorosoma, 5
видов), Южной и Юго-Восточной Азии и
Западной Океании (Меланезии) (роды Nematalosa, Anodontostoma, Gonialosa,
всего 7 видов), Восточной Азии (роды Konosirus, Clupanodon, Nematalosa, 3 вида),
Австралии (роды Nematalosa, 1 вид, и
Fluvialosa, 7 видов). У более северных
видов — японского коносира и
американской доросомы — позвонков 48—51, у
остальных 40—46.
Подсемейство Килегорлые, или Пилобрюхие, сельди (Pristigasterinae)
Эта группа чисто тропических родов
сельдевых рыб характеризуется сильно
сжатым с боков, приостренным по брюшному
краю телом, с пиловидно-зазубренным
брюшным килем из чешуи,
простирающимся и вперед, на горло. Рот почти у всех
верхний или полуверхний. Анальный
плавник у них длинный, содержащий
более 30 лучей; брюшные плавники
малы (у Pellona и Ilisha) или отсутствуют
(у остальных родов). В эту группу
входят 8 родов с 37 видами.
По внешнему виду разные роды пилобрюхих сельдей представляют различные
ступени специализации. Наименее
специализированы и несколько напоминают по
внешнему виду алоз или гильз упомянутые уже
рыбы родов Пеллона (Pellona) и Илиша
(Ilisha). (см. Pristigasteridae)
Остальные 6 родов пилобрюхих сельдей
лишены брюшных плавников. Очень
своеобразен из них Пристигастер (Pristigaster). (см. Pristigasteridae)
Из остальных 5 родов пилобрюхих
сельдей три рода американские,
встречающиеся либо только у тихоокеанского
побережья Центральной Америки (род Рliоsteostoma), либо представленные одним
видом в тихоокеанских водах и одним-двумя видами — в атлантических (роды
Odontognathus, Neoopisthopterus). Один
род (Opisthopterus) представлен тремя
видами у тихоокеанского побережья
Панамского перешейка и Эквадора и двумя
видами в Индийском океане и в
юго-западной части Тихого океана, у берегов
Индии, Индокитая и Индонезии.
Обитатели бездны. Уильям Кроми 1971 г.
К гребневикам принадлежит и ненасытный морской крыжовник Pleurobranchia. Выросши, этот пират формой и размерами (около 2,5 сантиметра) становится похож на ягоду крупноплодного крыжовника. На нижней стороне его тела расположены рот и два щупальца, которые в десять раз длиннее «ягоды». Щупальца имеют огромное количество тонких, как волоски, ответвлений. Они покрыты узловатыми хватательными клетками, которые при прикосновении к ним выделяют клейкое вещество. Движимая вперед рядами гребневидных пластинок, находящихся по бокам, как у старинного колесного парохода, плевробранхия процеживает воду с помощью этой чудовищно уловистой рыболовной снасти. От нее добыча не уйдет. Жертва начинает отчаянно вырываться, но чем энергичнее она борется, тем крепче ее сжимают хватательные клетки. Мускульная сердцевина щупалец сокращается, подтягивая добычу к вытянутым «губам» животного. Эти хищники хватают молодь рыб, в два раза большую их самих. В желудках некоторых экземпляров находили до пяти мальков сельди.
Наиболее характерны морские стрелки, принадлежащие к типу щетинкочелюстных (Chaetognatha). Они живут только как планктон; в любой части Мирового океана можно встретить какую-либо разновидность животных этого типа. Наиболее распространенными — их вы наверняка увидите в любой пробе воды — являются представители рода Sagitta. Длиной около 2,5 сантиметра, эти прозрачные животные напоминают крохотные стеклянные палочки с двумя комплектами боковых плавников и горизонтальным хвостом в виде наконечника стрелы. Существа эти имеют два маленьких черных глаза, и с их помощью они могут смотреть во все стороны, не поворачивая головы. Они могут смотреть даже сквозь свое прозрачное туловище. По обеим сторонам головы у них прочные, искривленные щетинки, выполняющие роль мощных челюстей, которые делают этих животных сущими драконами мира планктона. Такой крохотный дракон лежит в воде неподвижно до тех пор, пока в «радиусе его действия» не окажется какая-нибудь жертва. Тогда, помогая себе мощными толчками хвоста, он устремляется вперед. Морская стрелка в мгновение ока может покрыть расстояние в пять-шесть раз большее ее собственной длины. Хватая жертву своими сильными щетинками, она пожирает молодь сельди величиной с самое себя, рачков-копепод и даже своих сородичей. Подобно гребневикам, «стеклянные» черви хорошо различимы лишь тогда, когда в их прозрачных желудках находится полупереваренная пища.
Весенней порой в бухте Кейп-Код в одном кубическом метре воды может оказаться до 2000 копепод. Сельдь, макрель и другие существа, проплывая сквозь такие скопища, поглощают животных в невероятных количествах. Так, в желудке одной лишь сельди было обнаружено ни много ни мало 60 000 копепод.
СЕЛЬДЬ И МЕНХАДЕН
Типичный житель открытого моря, где обитают многие виды рыб, маленькая сельдь, пожалуй, самая многочисленная рыба в приповерхностных слоях воды. Атлантическая сельдь и ее тихоокеанская сестра Clupea pallasi передвигаются огромными многомиллиардными косяками, которые подчас достигают длины в несколько миль и такой же ширины. Иногда кажется, что море битком набито их серебристыми, длиной сантиметров в 30 телами.
Это изобилие делает семейство сельдевых, куда входят менхаден и сардины, чрезвычайно важным для экономики моря и экономической жизни людей. Плавая близ поверхности, сельдь питается копеподами, птероподами, червями-стрелками и иными планктонными организмами. В свою очередь такие хищники, как треска, макрель, тунец, акулы, морские птицы и усатые киты, пожирают ежегодно столько сельди, что ею можно было бы загрузить состав, который опоясал бы земной шар. Люди каждый год добывают приблизительно 11 миллиардов сельдей. Вместе с сардинами и менхаденом они составляют четвертую часть всей рыбы, вылавливаемой человеком в соленых водах.
Рыбаки Новой Англии ежегодно доставляют в порты около 67 миллионов килограммов молодой сельди длиной от 7,5 до 12,5 сантиметра, которая выдается за консервированные «сардины». Этот невинный обман восходит к 1875 году, когда Россия вела военные действия (Очевидно, автор имеет в виду русско-турецкую войну, которая, правда, относится не к 1875, а к 1877—1878 гг.) и один ловкий упаковщик с залива Мэн продал некоему импортеру из Нью-Йорка несколько ящиков сельди под видом русских сардин, которые невозможно было достать ни за какие деньги.
Большеголовый, длиной в 30 сантиметров сородич сельди, менхаден служит объектом промысла, крупнейшего в США, если оценивать его по весу выловленной рыбы. Это наиболее распространенная рыба, которая водится по всему Атлантическому побережью страны от залива Мэн до Флориды, но мясо ее жирно и не слишком вкусно. Менхаден используется для приготовления удобрений и пищевых концентратов для откорма скота, а также для изготовления масляных растворителей для красок и лаков. Этот промысел — самый старый в Америке, он восходит к колониальному периоду, когда индейцы учили поселенцев использовать менхадена как удобрение для посевов злаковых.
Подобно другим пелагическим рыбам (то есть обитающим в открытом океане), сельдь и менхаден обладают удивительно обтекаемой формой. Рыбы настолько превосходно приспособлены для быстрого и маневренного передвижения в воде, что люди веками копировали их форму, конструируя подводные лодки, торпеды и иные предметы, которые должны двигаться в воде, создавая минимальное сопротивление. Поскольку сжать воду невозможно, рыба, чтобы продвигаться вперед, вынуждена «распихивать» ее в стороны. Животное делает это, рассекая массу воды более или менее заостренной головой, а затем расталкивая ее в стороны расширенной частью, после чего вода плавно скользит по конусообразному хвосту, создавая минимальные завихрения, лишь незначительно замедляющие движение.
Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Часть 2. (Acipenseriformes — Polynemiformes). Г.У.Линдберг, М.И.Легеза 1965 г.
XXXIV. Сем. CLUPEIDAE — СЕЛЬДЕВЫЕ
Боковая линия в виде прободенных чешуй на теле отсутствует, развита лишь в самой передней части в виде короткого канала с порами от первых 2 (у Sardina), до 5 (у Cluреа) чешуях сразу за головой. Отверстопузырные. Рот окаймлен посредством предчелюстных (praemaxillaria) и верхнечелюстных (maxillaria) или только посредством предчелюстных (у большинства Dorosomatinae). В верхней челюсти 1 или 2 дополнительные косточки — надчелюстные (supramaxillaria). Спинной плавник посередине тела, иногда (у Pristigasterinae) над анальным. Брюшные плавники на брюхе, иногда отсутствуют (у некоторых Pristigasterinae). Брюхо закругленное или сжатое с боков, брюшные килевые чешуи есть или отсутствуют (у Dussumieriinae), чешуя циклоидная или у некоторых (Brevoortia, Ethmalosa и Ethmidium) на заднем крае гребенчатая.
Около 50 родов и до 190 видов. 6 подсемейств (Световидов, 1952а: 91), из которых в водах Японского моря не представлено только 1 — Pellonulinae.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ СЕМ. CLUPEIDAE
1 (4). Брюшных килевых чешуй нет. Брюхо закругленное. (Подсем. Dussumieriinae). Верхнечелюстная кость не заходит за передний край глаза. Грудные плавники значительно короче половины расстояния от их начала до основания брюшных.
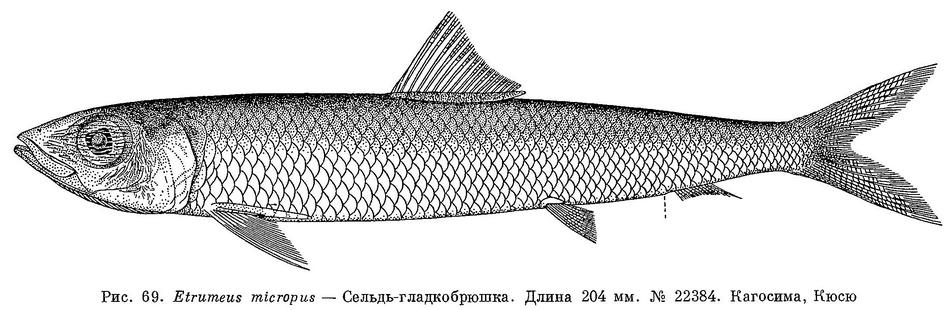
2 (3). Брюшные плавники расположены заметно позади заднего края основания спинного плавника. Лучей жаберной перепонки 14—19. Жировое веко полностью закрывает глаз. На боках тела нет широкой серебристой полосы. Длина половозрелых от 150 до 250 мм (рис. 69). 1. Etrumeus Bleeker.
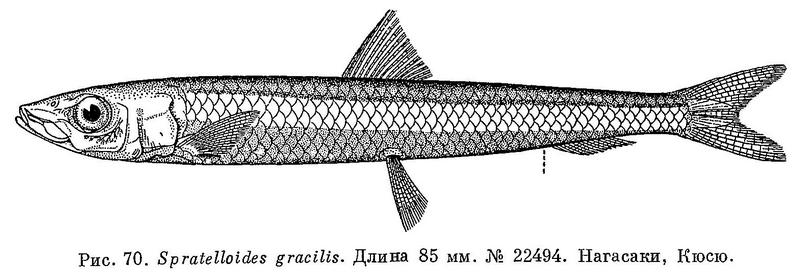
3 (2). Брюшные плавники расположены под основанием спинного плавника. Лучей жаберной перепонки 6—7. Жировое веко слабо развито. На боках тела широкая серебристая полоса. Длина половозрелых от 50 до 110 мм (рис. 70). (Род Dussumieria Cuvier et ValencienDes, 1847, отличается от рода Spratelloides большим числом лучей в анальном плавнике (15—17 вместо 9—13) и в жаберной перепонке (15—20 вместо 6), а также более задним положением спинного, расположенного ближе к хвостовому плавнику, а не к краю рыла. Этот род представлен к югу от Тайваня.) 2. Spratelloides Bleeker.
4 (1). Брюшные килевые чешуи есть, обычно хорошо развиты, реже слабые и менее заметные. Брюхо сжатое с боков и заостренное в виде киля, а если закругленное, то верхнечелюстная заходит за передний край глаза, а грудные плавники заметно длиннее половины расстояния от их начала до основания брюшных.
5 (18). Брюшные килевые чешуи не простираются на горло. Анальный плавник менее чем с 30 лучами; начало его далеко позади заднего конца спинного. Брюшные плавники под основанием спинного плавника.
6 (15). Последний луч спинного плавника не удлинен. Надчелюстных (дополнительных) костей 2.
7 (14). Верхняя челюсть без заметной медиальной вырезки. Сочленение нижней челюсти с черепом впереди вертикали заднего края глаза или под ней. (Подсем. Clupeinae).
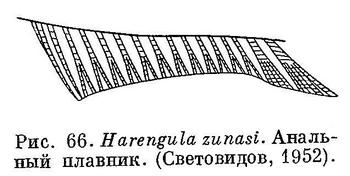
8 (11). Последние 2 луча анального плавника не удлинены в виде лопасти или удлинены очень слабо (рис. 66). Удлиненных чешуй (alae) при основании хвостового плавника на его лопастях нет.
9 (10). Грудные далеко не достигают начала спинного, (рис. 71). По следние 2 луча анального не удлинены. 3. Clupea Linne.

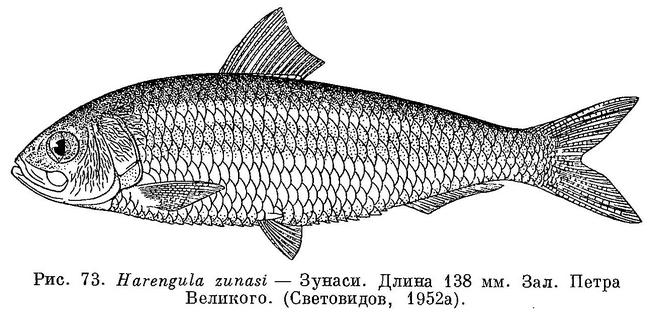
10 (9). Грудные достигают и даже заходят за начало спинного (рис. 73). Последние 2 луча анального плавника слегка удлинены, но не
образуют выдающейся свободной лопасти (рис. 66). 4. Harengula Valenciennes.
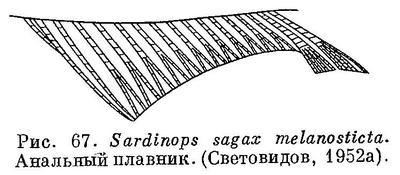
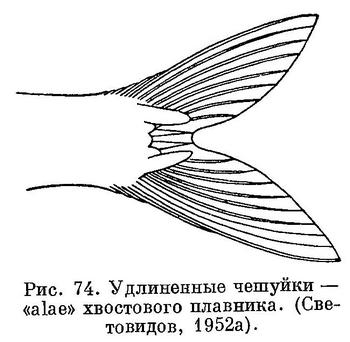
11 (8). Последние 2 луча анального плавника сильно удлинены и образуют отчетливо выступающую лопасть (рис. 67). Удлиненные чешуи (alae) при основании хвостового плавника имеются. (рис. 74).
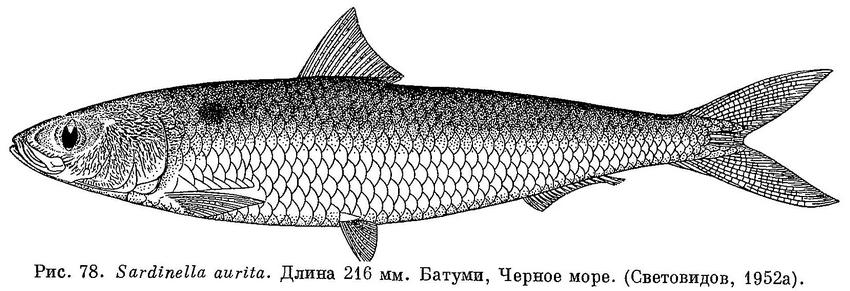
12 (13). Нет радиальных бороздок на крышечной кости. На боку тела позади жаберной крышки 1 пятно (рис. 78). 5. Sardinella Valenciennes.
13 (12). Есть радиальные бороздки на крышечной кости. На боку тела позади жаберной крышки несколько (до 8) пятен, расположенных в ряд (рис. 80). 6. Sardinops Hubbs.

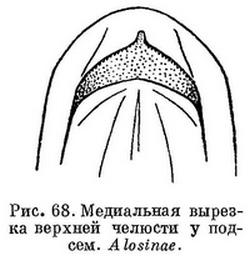
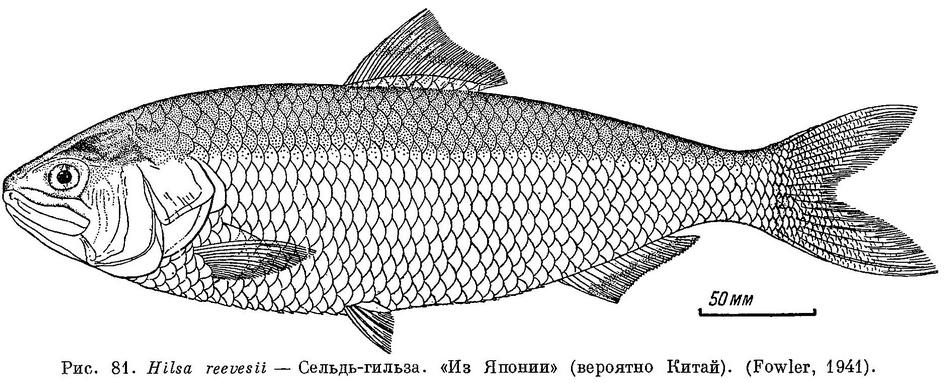
14 (7). На верхней челюсти есть медиальная вырезка (рис. 68). Сочленение нижней челюсти с черепом за вертикалью заднего края глаза. (Подсем. Alosinae; рис. 81). 7. Hilsa Regan.
15 (6). Последний луч спинного плавника вытянут в длинную нить. Надчелюстных (дополнительных) костей 1. (Подсем. Dorosomatinae).
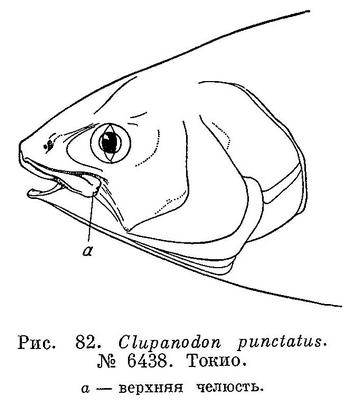
16 (17). Верхнечелюстная кость обычной формы; кость уплощена, а в задней части расширена и не изогнута резко книзу. Предчелюстная кость заметно не достигает конца верхнечелюстной; длина задней части верхнечелюстной, от вертикали конца предчелюстной, примерно равна длине предчелюстной (рис. 82). 8. Clupanodon Lacepède.
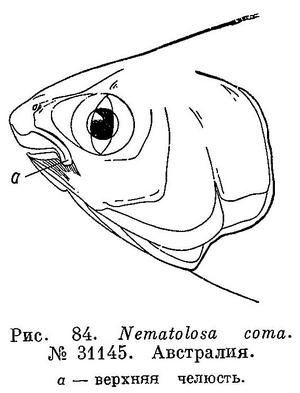
17 (16). Верхнечелюстная кость не совсем обычной формы; кость не уплощена, узкая, в задней части не расширена и резко изогнута книзу. Предчелюстная кость, также резко изогнутая книзу, простирается почти до конца верхнечелюстной кости; благодаря этому изгибу хорошо виден задний конец dentale (зубной кости нижней челюсти; рис. 84). 9. Nematolosa Regan.
18 (5). Брюшные килевые чешуи простираются на горло. Анальный плавник длинный, более чем с 30 лучами. Нижняя челюсть выдается вперед верхней. (Подсем. Pristigasterinae).
19 (20). Брюшные плавники имеются; они малы и расположены впереди вертикали начала спинного плавника, который почти целиком впереди анального. 10. Ilisha Richardson.
20 (19). Брюшных плавников нет. Начало спинного плавника впереди начала анального, но значительная часть основания спинного расположена позади начала анального. (Род Opisthopteris Gill, 1861, также не имеющий брюшных плавников, отличается более задним положением спинного плавника; его начало находится позади начала анального (Matsubara, 1955 : 190). Распространен у берегов Китая и южнее.) 11. Pristigaster Cuvier.
Рыбы северных морей СССР. А.П.Андрияшев 1954 г.
XI. Сем. CLUPEIDAE — СЕЛЬДЕВЫЕ
Regan, Ann. Mag. Nat. Hist., (8), XIX, 1917 : 297. — Берг, Система рыб, 1940 : 223. — Берг, Рыбы пресн. вод СССР, I, 1948 : 110. — Световидов. Сельдевые, Фауна СССР, Рыбы, II, 1, 1952 : 91 (монографич. сводка).
Тело покрыто чешуей, голова голая. Боковая линия не развита, имеется лишь 2—5 передних пор. Каналы системы боковой линии на голове сильно развиты, заключены в костные каналы и имеют вторичные ответвления во многих покровных костях черепа, жаберной крышки, нижней челюсти. Плавательный пузырь соединен с желудком, его передние выросты через occipitale laterale входят в ушную капсулу (в prooticum и обычно в pteroticum). Рот не выдвижной, край верхней челюсти образуют maxillaria и praemaxillaria, реже только последнее (большинство Dorosomatinae). Есть 1 или 2 supramaxillaria. Жаберные перепонки свободны от межжаберного промежутка. Жаберные тычинки длинные, тонкие. Жирового плавника нет. Брюшные плавники на брюхе, реже отсутствуют. Спинной плавник у взрослых впереди вертикали анального
(у рода Raconda спинной отсутствует). Хвостовой плавник с глубокой выемкой. Postcleithrum прикрепляется к наружной стороне cleithrum. Сошник и мезэтмоид парные. Парапофизы не приращены к телам позвонков. Есть нормальные яйцеводы. Имеются нижние и верхние ребра; дистальная часть последних хрящевая, проксимальная — соединительнотканного происхождения. Есть верхние и нижние межмышечные косточки. Известны с нижнего мела.
Около 50 родов и 190 видов (по Световидову) в тропических и умеренных, реже в арктических морях. Многие заходят в реки, некоторые постоянно живут в пресной воде. В мировом промысле сельдевые занимают первое место; уловы их достигают до 63 млн. ц в год (37 % мирового улова всех рыб) (Т.С. Расс. Мировой промысел водных животных. 1948 : 20.). В северных морях один род. При характеристике сельдей широко использована монография А. Н. Световидова, рукопись которой была любезно представлена мне автором до ее опубликования.
Фауна СССР. Рыбы. Том II, вып.1. Сельдевые (Clupeidae). А.Н.Световидов 1952 г.
КРАТКИЙ ИСТОРИЧЕСКИЙ ОЧЕРК СИСТЕМЫ СЕМЕЙСТВА
Отношение семейства сельдевых к близким семействам и положение его в системах, предложенных рядом авторов, в зависимости от признаков, принятых за основу в этих системах, и положения в системах отряда, к которому относились сельдевые, различно. Не останавливаясь на этом детально, укажем, что обзор систем рыб с конца XVII в. по XX в. дан Леннбергом (Lönnberg, 1924 : 43—62), позднейших работ — Л. С. Бергом (1940 : 88—95), где интересующиеся могут найти необходимые сведения. Здесь отметим только, что в новейшей системе рыб, предложенной Бергом (1940 : 221—227), сем. Clupeidae относится к самому обширному из 18 подотрядов отряда Clupeiformes — подотр. Clupeoidei, который разделяется следующим образом:

Объем сем. Clupeidae как у авторов раннего периода ихтиологии, так и у позднейших точно так же различен. Объем семейства первыми авторами принимался крайне широко, со включением ряда близких и далеко стоящих от него семейств как отряда Clupeiformes современных авторов, так и других отрядов.
Не вдаваясь в детали, укажем, что Кювье (Cuvier, 1829 : 316) относил к сельдевым представителей семейств Elopidae, Magalopidae, Engraulidae, Chirocentridae, Hyodontidae, Notopteridae и Osteoglossidae отряда Clupeiformes современных авторов и ряда семейств (Polypteridae, Amiidaey, Lepidosteidae, Sudidae и др.), относимых в настоящее время к другим отрядам.
В значительно меньшем объеме рассматривал сельдевых (fam. Clupeoidei) Мюллер (Müller, 1843 : 324), включивший в него помимо родов, относимых современными авторами к сем. Clupeidae, некоторые роды, рассматриваемые в настоящее время как относящиеся в основном к другим семействам Clupeiformes (Engraulidae, Hyodontidae, Elopidae, Alepocephalidae). Семейства Polypteridae и Lepidosteidae Мюллер (Müller, 1846 : 202) в замечательной для своего времени системе рыб относит не к сельдевым, а к другому подклассу (Ganoidei).
Приблизительно в таком же объеме рассматривалось сем. Clupeidae Гюнтером (Günther, 1868 : 381), который делит его на следующие группы - I Engraulina, II Chatoessina, III Clupeina, IV Dussumieriina, V Albulina, VI Elopina, VII Chanina. Из этих групп в настоящее время к сем. Clupeidae относятся лишь группы Chatoessina, Clupeina и Dussumieriina. Группа Engraulina является самостоятельным, близким к Clupeidat семейством, Albulina и Elopina — более резко отграниченными семействами подотряда Clupeoidei, Chanina — семейством, относящимся к отряду Clupeiformes, но к особому подотряду Chanoidei.
Преобладающее большинство последующих авторов сем. Clupeidae принимало в значительно меньшем объеме, в объеме группы Clupeina Гюнтера, или включало в него и группы Chatoessina и Dussumieriina. Остальные группы Гюнтера рассматривались в качестве семейств отр. Clupeiformes (Isospondili, Malacopterygii s. str. auct.).
В большем объеме сем. Clupeidae рассматривалось лишь немногими авторами; из числа их отметим Буланже (Bulenger, 1904: 562—565) и Гудрича (Goodrich, 1909 : 391—393), включивших в сем. Clupeidae из ныне живущих сельдевых группы Engraulina и Chanina Гюнтера, причем группа Dussumieriina включена ими в подсем. Engraulinae, a Chatoessina — в подсем. Clupeinae. Всех ныне живущих сельдевых оба автора делят на подсемейства Engraulinae, Clupeinae и Chaninae.
У большинства других авторов разногласия существовали лишь в отношении групп Chatoessina и Dussumieriina.
Джордэн и Эверманн (Jordan a. Evermann, 1896 : 417) включают группу Dussumieriina в сем. Clupeidae, которое они разделяют на подсемейства Dussumieriinae, Clupeinae и Pristigasterinae. Группа Chatoessina рассматривается ими в качестве самостоятельного семейства (Dorosomidae; цит. раб. : 415). Эта группа как самостоятельное семейство (Dorosomatidae) принималась также Джордэном и Херре (Jordan а. Herre, 1906 : 623). Ригэн (Regan, 1917b : 297), наоборот, считает, что выделение сем. Dorosomidae в самостоятельное или даже в группу внутри семейства искусственно. Позднее Джордэн (Jordan, 1923 : 120—122) принимает сем. Clupeidae в более узком смысле, выделяя из него сем. Dussumieriidae и Dorosomidae. Последние два семейства в качестве самостоятельных рассматриваются и рядом других авторов (McCulloch, 1929 : 36, 41; Fowler, 1941 : 561). Берг (1940 : 223, 224), наоборот, включает эти семейства в Clupeidae, которое он разделяет на подсем. Dussumieriini, Clupeini и Dorosomatini. Не останавливаясь здесь на этом вопросе подробнее, укажем, что оба упомянутых семейства имеют основания для выделения их самое большее в ранг подсемейства.
Классификация сельдевых внутри семейства разработана слабо, как это отчасти видно из сказанного. Подсемейства, как указывалось выше, различали Джордэн и Эверманн и Л. С. Берг. Из других авторов, разделявших сем. Clupeidae, следует отметить Расса (1936 : 299), который по типам строения икринок выделил среди сельдевых северной части Атлантического океана три группы: 1) Clupea, Sprattus, Pomolobus (сюда он относит также Spratelloides = Stolephorus), 2) Alosa, Caspialosa, 3) Sardina, Sardinella, Clupeonella, Brevoortia. Отмечая, что это деление не стоит в согласовании с группировками, установленными для рассматриваемых родов Ригэном (Regan, 1917с : 377) на основании наличия или отсутствия в верхней челюсти медиальной вырезки, Т. С. Расс считает свое деление более обоснованным. В подтверждение большей обоснованности своих группировок им приведены лишь особенности строения икринок, однако признак, положенный в основу группировок Ригэна, довольно существен. Действительно, род Brevoortia весьма своеобразен, но медиальная вырезка у него все же есть, так же как она имеется и у Pomolobus. Вообще оба эти рода по ряду признаков гораздо ближе ко второй группе Т. С. Расса и менее к тем, в которые они им помещены. Что же касается Spratelloides, относимого в первую группу, то этот род принадлежит к подсем. Dussumieriinae, принимаемому многими, как указывалось выше, даже за особое семейство.
СИСТЕМА СЕМЕЙСТВА И ПРИНЦИПЫ КЛАССИФИКАЦИИ
Морфология представителей сем. Clupeidae изучена еще слабо; достаточно резких и существенных признаков для выделения подсемейств известно мало, причем большинство их — признаки внешнеморфологические. На основании ряда морфологических особенностей семейство провизорно может быть разделено на следующие 6 подсемейств: I Dussumieriinae, II Clupeinae, III Pellonulinae, IV Alosinae, V Dorosomatinae, VI Pristigasterinae.
Принципы классификации сельдевых, как, впрочем, и преобладающего большинства семейств рыб, за исключением немногих, точно так же не разработаны. Некоторые соображения о примитивности или высоте организации высказывались, как будет указано ниже, в отношении лишь отдельных форм видов или самое большее немногих видов сельдевых и притом на основании лишь единичных морфологических особенностей. Классификация же, как неоднократно подчеркивал автор (1946 : 183; 1948 : 19), должна основываться на комплексе морфологических особенностей, функционально связанных с наиболее характерной биологической особенностью рассматриваемой систематической группы и выработанных в результате взаимодействия организма со средой. Наичаще комплекс морфологических особенностей, положенных в основу системы, бывает связан с движением и обитанием в различных частях водной среды, реже с питанием, иногда с размножением изучаемой систематической группы. Изменение комплекса морфологических особенностей обычно сопровождается изменением их в процессе онтогенеза рассматриваемой группы. Поэтому особенно важно учитывать и данные онтогенеза, которые обычно являются исходной точкой при построении системы.
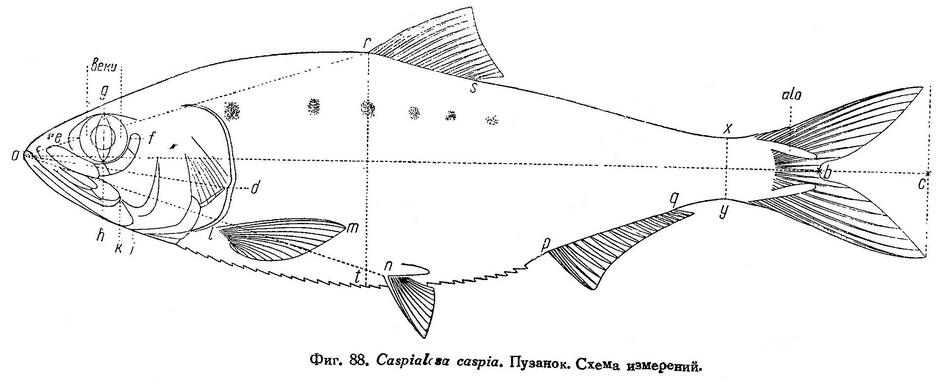
Одной из особенностей развития личинок и строения взрослых сельдевых следует считать различие в положении спинного плавника. Как известно, личинки сельдевых по сравнению со взрослыми, подобно личинкам и малькам многих костистых рыб, характеризуются сильно удлиненной передней частью тела, вследствие чего брюшные, анальный и спинной плавники у них отнесены далеко назад и, что самое существенное и свойственно только сельдевым и некоторым близким к ним семействам отр. Clupeiformes, спинной плавник у них расположен не над брюшными плавниками, как у взрослых, а значительно далее назади, между брюшными и анальным или почти над анальным (у Engraulidae). В процессе метаморфоза личинок передняя часть тела у них укорачивается вследствие сложного перемещения плавников. Спинной плавник при этом перемещается над миотомами вперед, а брюшные плавники назад, поэтому взаимное расположение обоих плавников становится таким, как у взрослых. Одновременно с этим анальный плавник перемещается вперед (Lebour, 1921; Ford, 1930). У взрослых сельдевых спинной плавник расположен над или несколько позади от брюшных в непосредственной к ним близости. При этом у разных видов семейства и видов одного и того же рода и даже форм вида положение спинного, брюшных и анального плавников неодинаково: у одних они расположены ближе к переднему концу тела, у других — дальше. Обусловлено это тем, что у разных родов и видов плавники закладываются в различном удалении от переднего конца тела, не над одними и теми же миотомами, а в дальнейшем степень перемещения плавников у разных родов и видов неодинакова (Lebour, 1921; Ford, 1930). Перемещение спинного плавника заканчивается к концу метаморфоза личинок и неоформившихся мальков, и у вполне сформировавшихся мальков и у молоди он расположен уже, как у взрослых. В дальнейшем, по мере роста молоди и взрослых рыб, как это видно на примере каспийской тюльки (Clupeonella delicatula caspia), положение анального и брюшного плавников несколько изменяется. (На каждый класс размеров приходится по 10 рыб. Как молодь, так и взрослые рыбы происходят из одного места (от западных берегов средней части Каспийского моря) и пойманы приблизительно в одно время. Такие же цифры рассматриваемых признаков получены автором и для тюльки из северной части моря перед дельтой Волги.)

Спинной плавник, как видно из приведенных примеров, занимает постоянное положение как у взрослых, половозрелых рыб, так и у рыб, еще не достигших половой зрелости. Что касается анального и брюшного плавников, то положение их на протяжении жизни рыбы претерпевает перемещение, судя по всему, в связи с половым созреванием вследствие растяжения брюшной стенки развивающимися половыми продуктами. Увеличение антеанального и антевентрального расстояний происходит по достижении рыбами размеров 70—80 мм, что приблизительно соответствует половому созреванию. В дальнейшем антеанальное расстояние остается примерно постоянным, увеличиваясь в незначительной степени, на доли процента. Антевентральное расстояние изменяется несколько в большей степени, повидимому, в связи с различным растяжением брюшной стенки вследствие разной степени наполнения полости тела половыми продуктами, у исследованных крупных рыб отчасти уже выметанными. К такому же выводу о постоянстве антедорсального расстояния, характеризующего положение спинного плавника у взрослых черноморско-азовских проходных сельдей (Alosa kessleri pontica), пришла Майорова (1939 : 7), а для черноморского анчоуса (Engraulis encrasicholus ponticus), относящегося к близкому сельдевым семейству того же подотряда, С. М. Малятский и отчасти Александров (1927 : 45, 46). Точно так же и относительно атлантической сельди (Clupea harengus harengus) установлено, что у взрослых рыб длиной свыше 10 см положение спинного, брюшных и анального плавников остается с возрастом неизменным (Heincke, 1898 : 113). Таким образом, у взрослых сельдевых положение спинного, а в известной мере также анального и брюшного плавников следует считать практически неизменным.
Положение спинного плавника у сельдевых несомненно имеет важное филогенетическое значение, и ряд авторов, устанавливая филогенетические отношения некоторых Clupeidae (Малятский, 1930 : 69) и близких к ним Engraulidae (Fage, 1911 : 25; 1920 : 31; Александров, 1927 : 83) и кладя в основу своих построений антедорсальное расстояние (у Engraulidae) или постдорсальное (у Clupeidae), имеют в сущности дело с положением спинного плавника. Помимо значительного перемещения при развитии личинок и различного положения у взрослых сельдевых спинной плавник имеет неодинаковое положение и у других, в особенности более примитивных семейств отряда. У наиболее примитивных из Clupeiformes, подотрядов Lycopteroidei (сем. Lycopteridae, нижний мел Забайкалья, Монголии и сев. Китая; Берг, 1940 : 218) и Leptolepidoidei (сем. Leptolepidae, от верхнего триаса до среднего мела; Берг, там же : 219; Woodward, 1895 : 500—530; 1919 : 121—139; Rayner, 1937 : 46—74), спинной плавник отнесен далеко назад, причем у Lycopteridae начало его расположено над анальным или слегка впереди его, а у более высокоорганизованных Leptolepidae спинной плавник расположен по середине между анальным и брюшным. У более примитивных, чем сельдевые, современных семейств Elopidae, Megalopidae (оба семейства известны с нижнего мела) и Albulidae (известны с палеоцена), относимых вместе с сельдевыми к одному и тому же подотряду Clupeoidei, спинной плавник расположен так же, как у Clupeidae. Следовательно, некоторые особенности в положении спинного плавника, которые у ископаемых семейств отряда являются конечной стадией, у современных семейств, в частности у сельдевых, свойственны только личинкам и являются лишь онтостадией. Однако есть основание считать, что личиночная стадия у более примитивных семейств Clupeoidei, сем. Elopidae и Albulidae, заканчивается при более крупных размерах и длится значительно дольше, чем у сем. Clupeidae. В частности, личиночная стадия Albulidae, повидимому, в некоторой степени даже аналогична личиночной стадии угрей. Личинка Albula была описана в качестве особого рода Esunculus (Kaup, 1856 : 143, табл. XVI, фиг. 3) и имеет значительно более крупные размеры, чем личинки сем. Clupeidae (Fitch, 1950).
Одной из наиболее характерных биологических особенностей сельдевых является их пелагический образ жизни, причем по образу жизни их следует считать скорее неритическими, чем океаническими. Большинство из сельдевых, ведя пелагический образ жизни, живет в море; значительная часть является проходными или пресноводными — речными или озерными. Сельдевые, живущие в море, имеют разную степень подвижности — одни из них совершают значительные по протяженности миграции, другие менее подвижны и совершают лишь ограниченные миграции. Одни из сельдевых живут на некотором отдалении от берегов, другие держатся у берегов, в заливах, бухтах и пр.
Сопоставление положения плавников на теле с образом жизни дает возможность установить известную функциональную связь между рассматриваемыми признаками и биологическими особенностями сельдевых.
По положению спинного плавника роды и виды подсем. Clupeinae располагаются в следующий ряд (табл. 1, рис. 1).
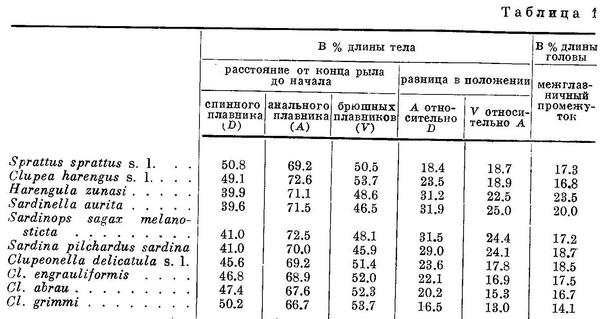
Примечание. За длину тела в табл. 1—8 принято расстояние от конца рыла до конца средних лучей хвостового плавника. Это сделано как для единообразия всех рассматриваемых величин, так и вследствие большей точности измерения длины тела до конца средних лучей хвостового плавника, чем до конца лопастей его.
Соответственно изменению положения спинного плавника относительно переднего конца тела закономерно изменяется положение брюшного и анального плавников, причем положение спинного и брюшного плавников на протяжении ряда претерпевает изменение в одном направлении, изменение же положения анального плавника — в обратном.
Каковы же причины различного положения спинного, анального и брюшного плавников? Об этом отчасти уже сказано выше. Прежде
всего различие это обусловлено неодинаковым положением плавников у личинок. В дальнейшем, в процессе метаморфоза личинок, спинной, анальный и брюшной плавники у разных представителей семейства перемещаются на неодинаковое расстояние. В подсем. Clupeinae, по исследованиям Форда (Ford, 1930) и Лебаур (Lebour, 1921), у Sprattus и Clupea, имеющих почти одинаковую удаленность спинного плавника от переднего конца тела, этот плавник перемещается вперед на равное число миотомов (4—5). У Sardina, стоящего в середине подсемейства, по исследованиям тех же авторов, спинной плавник, расположенный к переднему концу тела значительно ближе, перемещается вперед уже на 10 миотомов. Следовательно, более близкое положение к переднему концу тела спинного плавника у родов, расположенных в середине подсемейства, обусловлено большим перемещением его вперед, чем у родов, расположенных в начале и, вероятно, в конце подсемейства. Анальный плавник наиболее далеко отнесен назад у Clupea. Соответственно этому у личинок этого рода он перемещается вперед всего лишь на 2 миотома. У личинок Sprattus и Sardina анальный плавник перемещается вперед на большее число миотомов (на 4—5 у первого и на 5—6 у второго), вследствие чего и во взрослом состоянии этот плавник у них расположен ближе к переднему концу тела, чем у Clupea. Таким образом, различие в положении рассматриваемых плавников обусловлено неодинаковым перемещением его в процессе метаморфоза личинок, а большее различие родов подсемейства по положению спинного плавника по сравнению с различием в положении брюшных плавников — большим перемещением первого, чем последних.
Соответственно изменению положения относительно переднего конца тела спинного плавника положение анального и брюшных плавников изменяется не во всех случаях закономерно. Так, анальный плавник у Clupea отнесен несколько более назад по сравнению со стоящими дальше от него родами, брюшные плавники несколько более удалены от переднего конца тела у Clupea и Sardinops. Строгая закономерность, параллельная изменению положения спинного планника относительно переднего конца тела, наблюдается в изменении положения анального плавника относительно спинного и брбшного относительно анального. При этом разница в положении анального плавника относительно спинного, по мере уменьшения расстоянии от переднего конца тела до спинного плавника и увеличения до анального, на протяжении ряда естественно увеличивается (табл. 1). По этой же причине разница в положении брюшных плавников относительно анального увеличивается. Изменение положения брюшных плавников относительно спинного закономерного характера не имеет. Разница в положении анального плавника относительно спинного здесь и в дальнейшем изложении выражается разностью между расстоянием обоих плавников от переднего конца тела. Равным образом разница в положении брюшных плавников относительно анального выражается разностью между расстояниями этих плавников от переднего конца тела. Точнее говоря, разница во взаимном положении этих плавников представлена виде проекции расстояний между ними на продольную ось тела, выраженную в процентах длины тела.
Параллельно с изменением положения спинного плавника, с уменьшением расстояния от него до переднего конца тела, ширина головы (межглазничный промежуток) увеличивается. В дальнейшем с увеличением расстояния от начала спинного плавника до переднего конца тела ширина головы уменьшается. Следует отметить, что и здесь Clupea и Sardinops несколько уклоняются от общего закономерного характера изменения этого признака на протяжении подсемейства. Что касается длины головы и числа лучей в анальном и спинном плавниках, характеризующих размеры (длину основания) этих плавников и изменяющихся строго закономерно в других рассматриваемых ниже группах сельдевых, то в подсем. Clupeinae эти особенности такого строгого параллельного характера не имеют и поэтому в таблице не приведены. Однако и здесь с изменением положения спинного плавника ближе к переднему концу тела число лучей в анальном плавнике в общем уменьшается, а число лучей в спинном плавнике, наоборот, увеличивается. Размеры головы точно так же, как и в рассматриваемых ниже группах сельдевых, с увеличением ее ширины уменьшаются.
Кроме того, параллельно с изменением положения спинного плавника относительно переднего конца тела и разницы в положении плавников относительно друг друга изменяется и степень развития каналов системы боковой линии на голове. Наиболее развиты каналы системы боковой линии на голове у родов со спинным плавником, расположенным ближе к переднему концу тела и с наибольшей разницей в положении плавников относительно друг друга, в частности у Sardina (Третьяков, 1950, рис. 5; Wolfahrt, 1937, рис. 1, 10), Sardinops и Sardinella. Значительно менее развиты эти каналы у родов со спинным плавником, отнесенным далее назад, и с меньшей разницей в положении плавников относительно друг друга, например у Clupea (Bamford, 1941, рис. 9), и еще менее у Sprattus и Clupeonella (Третьяков, 1938, рис. 3; 1950, рис. 2). Посредством каналов системы боковой линии, как установлено, рыба ориентируется в течениях, воспринимает неподвижные (например берега и пр.), а также движущиеся предметы, в том числе, что крайне важно для стайных сельдевых, рыб и даже мелкие пищевые объекты (Пучков, 1941 : 262—266; Wunder, 1936 : 53—67).
Ископаемые виды рода Clupeonella по основным из рассматриваемых особенностей более примитивны, чем современные, причем Богачев (1938 : 47) отмечает, что один из известных понтических видов (Clupeonella vexata) имеет значительное сходство с современным Clupeonella engrauliformis. Однако голова у Clupeonella vexata значительно крупнее, чем даже у наиболее большеголового из современных видов (Clupeonella grimmi). У другого ископаемого вида (Clupeonella binagadensis), провизорно относимого тоже к роду Clupeonella, длина головы еще больше. По расстоянию спинного плавника от конца рыла, составляющему около 49—50% длины тела, Clupeonella vexata близок к наиболее примитивному из современных видов.
Помимо изменения ширины в межглазничной области, ширина головы изменяется и в других ее частях, почему выше, ссылаясь на изменение межглазничного промежутка, говорилось об изменении ширины головы вообще. Изменение ширины головы в других ее частях у видов рода Clupeonella видно из данных измерений ширины черепа:

Изменение всех рассматриваемых признаков, за исключением, как указывалось, числа лучей в плавниках и длины головы, происходит строго закономерно, причем численные значения одних из них (положение спинного плавника и брюшных относительно переднего конца тела, число лучей в анальном плавнике) по направлению от начала ряда к середине убывают, возрастая вновь к концу ряда, других (положение анального плавника относительно переднего конца тела и относительно спинного, брюшных относительно анального, межглазничный промежуток, число лучей в спинном плавнике), наоборот, возрастают к середине ряда и убывают вновь к концу его. Иными словами, спинной и брюшные плавники от начала ряда к середине его перемещаются к переднему концу тела, претерпевая перемещение в обратном направлении от середины к концу ряда. Анальный плавник, наоборот, в направлении от начала ряда к середине от переднего конца тела и от спинного плавника удаляется, перемещаясь вновь к переднему концу тела и к спинному плавнику от середины к концу ряда, равно как и брюшные плавники по отношению к анальному. Ширина головы в направлении от начала ряда к середине увеличивается, длина уменьшается; в направлении от середины ряда к концу ширина головы, наоборот, уменьшается, а длина увеличивается.
Роды, стоящие в первой и второй половинах ряда, существенно различаются тем, что у родов первой половины (роды Sprattus, Clupea) анальный плавник имеет нормальное строение, а у родов второй (роды Sardinella, Sardinops, Sardina, Clupeonella) два последних луча анального плавника удлинены и выдаются в виде лопасти. Род Harengula занимает промежуточное положение между этими двумя группами, так как два последних луча анального плавника у него удлинены слабо, не образуя лопасти.
Роды, расположенные н середине ряда (Harengula, Sardinella, Sardinops, Sardina), характерны развитием у большинства их пигментных темных пятен на боках тела, совершенно отсутствующих у родов, расположенных в начале и конце ряда. У большинства видов рода Harengula, как и у изображенного на табл. V Harengula zunasi, пятен на теле нет; лишь у некоторых видов (Harengula humeralis, Harengula thrissina, Harengula clupeola, Harengula majorina и др.) с каждой стороны тела есть по темному пятну за жаберной крышкой и у немногих из них — сзади него иногда по ряду неясно выраженных пятен на боках тела. Нет пятен на теле и у большинства видов рода Sardinella, и лишь некоторые из них имеют, как и Sardinella aurita, по темному пятну за жаберной крышкой. У Sardinops с каждой стороны на боках тела всегда имеется от нескольких до 15 расположенных в ряд темных пятен; ниже этого ряда у Sardinops sagax melanosticta изредка есть второй и иногда третий ряд из меньшего числа более светлых и неясных пятен. Ряд таких же темных пятен, как и у Sardinops, есть и у Sardina, но эти пятна на теле сверху бывают не всегда заметны, но обнаруживаются по удалении чешуй.
Следовательно, в развитии темных пигментных пятен на боках тела наблюдаются все переходы от наибольшей степени развития их у рода Sardinops, расположенного в середине ряда, к родам, стоящим в начале и конце ряда и не имеющим пятен совершенно. Отсутствие или наличие удлиненных лучей в анальном плавнике имеет переходный характер у рода Harengula, расположенного также по середине ряда.
Кроме того, роды, стоящие в середине ряда, связаны между собой некоторыми и другими морфологическими особенностями. У Sardinella, Sardinops и Sardina на хвостовом плавнике у основания лопастей есть удлиненные чешуи (ala), отсутствующие у Sprattus, Clupea и Harengula, стоящих в начале ряда, и у Clupeonella, стоящего в конце. Расположенные рядом роды Harengula и Sardinella имеют двулопастные кожистые выросты на вертикальной части ключицы, a Sardinops и Sardina — радиальные бороздки на крышечной кости.
Таким образом, изменение рассматриваемых морфологических особенностей на протяжении ряда происходит не прямолинейно (в сторону возрастания одних и убывания других), а циклически, представляя восходящую спираль. На протяжении этой спирали рассматриваемые морфологические особенности претерпевают закономерное изменение, причем удлинение двух последних лучей анального плавника имеет нарастающий характер; все остальные особенности (положение плавников относительно переднего конца тела и друг друга, пигментация и пр.) возрастают к середине ряда, убывая вновь к концу его и давая в начале и в конце сходные особенности.
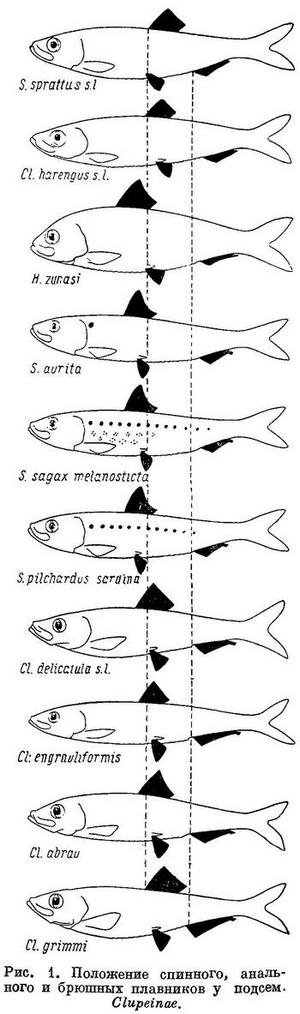
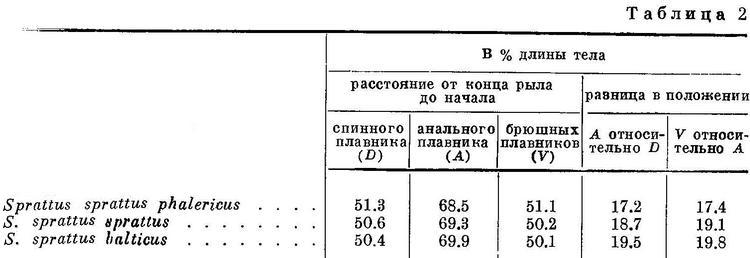
Это закономерное изменение рассматриваемых признаков стоит в тесной связи с изменением биологических особенностей родов, видов и форм подсем. Clupeinae, большинство представителей которого, за исключением нескольких пресноводных видов, является морскими рыбами. Роды (Sardinops, Sardina, Sardinella и в меньшей степени Harengula), у которых спинной и брюшные плавники расположены наиболее близко к переднему концу тела, а анальный плавник наиболее отнесен назад, ведут и наиболее подвижный образ жизни. Многие из них, в частности Sardinops и Sardina, совершают миграции большой протяженности. Роды, стоящие в начале и в конце ряда, с отнесенными назад спинными и брюшными плавниками, с расположенным ближе к переднему концу тела анальным плавником и наименее отстоящими анальным плавником от спинного и брюшными от анального менее подвижны. Из видов рода Clupeonella наиболее подвижный образ жизни ведет Clupeonella delicatula, каспийская форма которого (Clupeonella delicatula caspia) совершает миграции из южной части моря в северную и частично заходит даже в реки; в море эта форма держится ближе к берегам. Азовская форма этого вида (Clupeonella delicatula delicatula) совершает меньшие передвижения. Clupeonella engrauliformis держится в некотором отдалении от берегов в южной и средней частях Каспийского моря, иногда подходит и в прибрежную зону. Clupeonella abrau, населяющий небольшое пресноводное озеро, совершает более ограниченные миграции. Наконец, Clupeonella grimmi живет главным образом в южной части Каспийского моря и отчасти в средней, держится в открытом море в галистатической области, никогда не подходя к берегам, и на значительно больших глубинах, чем другие виды рода. Clupea совершает в общем меньшие по протяженности миграции, чем Sardinops, Sardina и другие роды, расположенные в середине ряда. Clupea harengus pallasi, Clupea harengus pallasi n. suworowi, Clupea harengus pallasi n. maris-albi и Clupea harengus membras живут и размножаются ближе к берегам, чем Clupea harengus harengus. Последний размножается на больших глубинах, часто в отдалении от берегов, на банках среди моря. Среди локальных форм Clupea harengus harengus есть такие, которые совершают значительные миграции (например мурманская сельдь), но у многих форм, в частности у некоторых в Северном море, миграции ограниченные. Формы Sprattus sprattus живут и размножаются в море, но не у самых берегов, а в некотором отдалении от них. Sprattus sprattus balticus ведет более прибрежный образ жизни, так как живет и размножается в заливах и фиордах; Sprattus sprattus phalericus держится и размножается в море, в большем отдалении от берегов.
Таким образом, изменение комплекса рассматриваемых морфологических особенностей, как видно из сказанного, стоит в тесной связи с образом жизни, в частности с движением, причем наиболее прогрессивные формы совершают большие по протяженности миграции.
Термины «прогрессивность» и «примитивность» понимаются здесь и далее в смысле большей или меньшей близости по рассматриваемым морфологическим особенностям к личинке и к стоящим ниже в системе семействам отряда. Вообще же по комплексу рассматриваемых морфологических особенностей каждый вид и форма наилучшим образом приспособлены к условиям, в которых они обитают, и к тому образу жизни, который они ведут. Вместе с тем наиболее прогрессивные по рассматриваемым морфологическим особенностям роды и виды подсемейства Clupeinae и, как будет видно в дальнейшем, виды родов других подсемейств, а также и формы ряда видов являются более прогрессивными и по биологическим особенностям. Критериями биологической прогрессивности и данном случае следует считать помимо протяженности миграций размеры ареала и большую численность вида в случае монотипических родов или большое число видов у политипических родов.
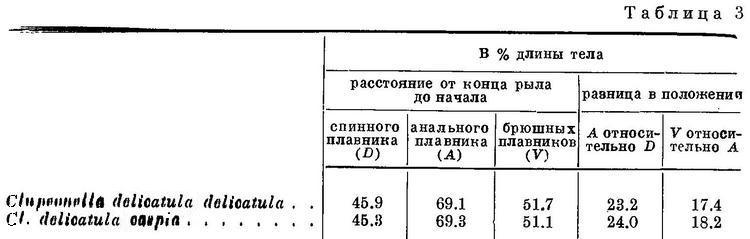
Наиболее широкий ареал из подсем. Clupeinae занимают более прогрессивные по рассматриваемым морфологическим особенностям — Clupea harengus (2 подвида со многими формами в умеренных частях Атлантического и Тихого океанов и в прилегающих морях), Harengula (около 19 видов в тропических частях Тихого, Индийского и западной части Атлантического океанов), Sardinella (около 16 видов в тропических и отчасти в субтропических частях Индийского, Атлантического и западной части Тихого океанов), Sardinops sagax (5 субтропических подвидов по обе стороны северной части Тихого океана и в южном полушарии у берегов Южн. Америки, Австралии, Новой Зеландии и южной Африки) и Sardina pilchardus (2 субтропических подвида и северной части Атлантического океана, в Средиземном и Черном морях. Все они отличаются или большой численностью, составляя основу мирового промысла (Clupea harengus, Sardina pilchardus и особенно Sardinops sagax), или, не имея большой численности и хозяйственного значения, большим количеством видов (роды Harengula и Sardinella). Следует отметить, что большим количеством видов отличаются тропические роды, субтропические и умеренные являются монотипическими, но отличаются, как указывалось, большой численностью. (Большая или меньшая численность обусловливает в общем большее или меньшее хозяйственное значение, если данный вид или форма его достаточно освоены промыслом. Поэтому характеристику численности во всех случаях см. «Хозяйственное значение».) Более примитивные по комплексу рассматриваемых признаков роды и виды занимают менее обширный ареал, совершают меньшие миграции и не имеют такой большой численности. Наименьший ареал в рассматриваемом подсемействе имеет род Clupeonella, а из видов его Clupeonella grimmi, несколько больший — Clupeonella engrauliformis и самый большой — Clupeonella delicatula, отличающийся и наибольшей численностью из всех видов рода и представленный двумя подвидами. Значительно больший, приблизительно такой же, как Sardina pilchardus, ареал занимает Sprattus sprattus. Численность этого вида меньше численности Sardina pilchardus.
Помимо рассматриваемых особенностей для суждения о высоте или примитивности организации сельдевых существенное значение иногда имеет степень развития зубов. У наиболее примитивных из современных Clupeoidei, сем. Elopidae и Albulidae, зубы развиты значительно сильнее, чем у сем. Clupeidae. У представителей сем. Clupeidae зубы имеются, причем не у всех только на челюстных костях, реже на сошнике и отчасти на нёбных костях и мезоптеригоиде, между тем как у сем. Elopidae и Albulidae зубы есть, кроме того, и на парасфеноиде (Берг, 1940 : 222; Boulenger, 1910 : 548). В пределах подсем. Clupeinae степень развития зубов не имеет закономерного изменения соответственно изменению положения плавников и длины и ширины головы, что вполне понятно, так как степень развития зубов стоит в связи с характером питания, рассматриваемые же особенности — с движением. Однако в пределах наиболее изученных родов и видов у более примитивных по рассматриваемым признакам зубы развиты сильнее. В частности, из подвидов Clupea harengus зубы на сошнике сильнее всего развиты у Clupea harengus harengus, более примитивного по положению плавников и по другим признакам, чем Clupea harengus pallasi и его формы (Clupea harengus pallasi n. maris-albi, Clupea harengus pallasi n. suworowi), у которых зубы на сошнике развиты менее. Равным образом у рода Sprattus зубы на сошнике имеются у некоторых видов из южного полушария (Sprattus antipodum, Sprattus muelleri), откуда ведет свое происхождение этот род; у вида из северного полушария (Sprattus sprattus) зубов на сошнике нет. Следует отметить, что у молоди некоторых видов зубы развиты сильнее, чем у взрослых.
Соответственно положению спинного плавника изменяется положение анального и брюшных плавников и у подвидов в пределах некоторых видов подсем. Clupeinae. Естественно, что степень различия рассматриваемых признаков между подвидами менее степени различий их у видов.
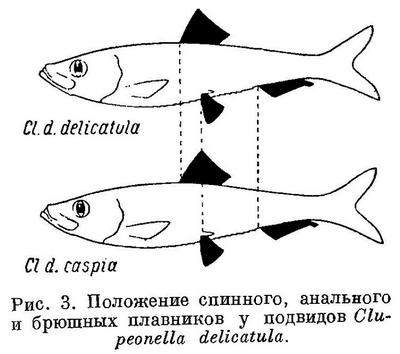
Положение анального и брюшных плавников относительно переднего конца тела, точно так же как первого относительно спинного и второго относительно анального у подвидов Sprattus sprattus и Clupeonella delicatula соответственно положению спинного плавника относительно переднего конца тела, изменяется таким же образом, как и у родов и у видов подсем. Clupeinae (табл. 2 и 3, рис. 2 и 3).
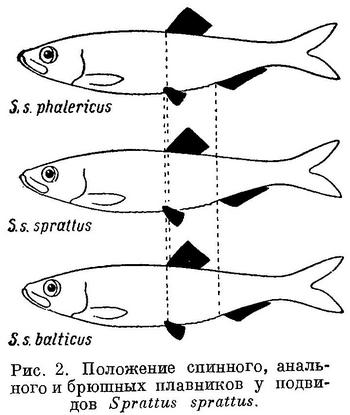
Изменение положения этих плавников у Clupea harengus, имеющего наибольшее количество подвидов, представлено в табл. 4 и на рис. 4. Следует отметить, что положение анального плавника у подвидов Clupea harengus изменяется в том же направлении, что и спинного и брюшных. Однако вследствие того, что на протяжении ряда положение анального плавника относительно переднего конца тела изменяется в большей степени, чем спинного и брюшных, разница в положении анального плавника относительно спинного также уменьшается, равно как и брюшных относительно анального плавника.
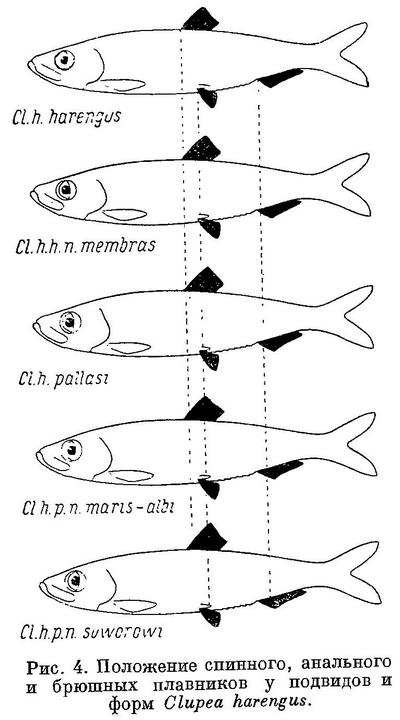
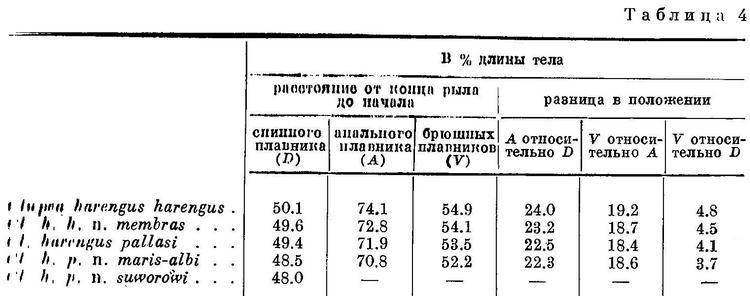
Spraltus sprattus balticus, Clupeonella delicatula caspia и Clupea harengus harengus, имеющие наибольшую разницу в положении анального плавника относительно спинного и брюшных относительно анального, совершают, как указывалось, и большие миграции, чем другие подвиды этих видов, у которых разница в положении этих плавников меньше. У подвидов Sprattus sprattus и Clupeonella delicatula положение анального плавника изменяется в обратном направлении, чем спинного и брюшных, у подвидов и форм Clupea harengus положение всех плавников изменяется в одном направлении.
Изучение изменения признаков, положенных в основу системы у подсем. Clupeinae, проведено на достаточно большом количестве рыб каждого вида и более низких систематических форм, что устраняет влияние индивидуальной изменчивости. Средние значения признаков каждого вида и форм отдельных видов, приведенные в табл. 1—7, основываются в среднем на 50 экз., по которым произведено описание этих видов и форм в систематической части, где указано количество исследованных рыб. Исключение составляют лишь немногие виды и формы, по которым исследовано меньшее количество экземпляров. Средние значения признаков Sprattus sprattus s. l., Clupea harengus s. l. и Clupeonella delicatula s. l. в табл. 1, Alosa caspia s. l. и др. в табл. 5 представляют среднее из средних значений их для всех форм данных видов. Отсутствие подобных материалов по другим подсемействам, встречающимся в преобладающей своей массе вне пределов наших вод, не даёт возможности остановиться на рассмотрении и их таким же образом. Поэтому в дальнейшем мы остановимся на рассмотрении лишь некоторых родов и полиморфных видов, встречающихся, в наших водах, а из подсемейств — лишь Dorosomatinae и Pristigasterinae, для которых имеющиеся литературные данные позволяют установить закономерное изменение числа лучей в анальном и спинном плавниках у родов в пределах обоих подсемейств.
По положению спинного и прочих плавников, по окраске и некоторым другим особенностям виды рода Alosa располагаются следующим образом (табл. 5, рис. 5). Как видно из этой таблицы и рисунка, по положению спинного и других плавников, по ширине и длине головы, а также по степени развития темных пигментных пятен на боках тела виды рода Alosa располагаются в ряд, который, повидимому, является частью общего ряда подсем. Alosinae. Изменение всех рассматриваемых признаков происходит в общем закономерно, за исключением некоторых из них у Alosa alosa и Alosa sapidissima, что обусловлено в основном малым количеством исследованных рыб (4 экз. первого и 2 экз. второго). При этом следует отметить, что изменение положения всех плавников относительно переднего конца тела происходит в одном направлении, как у форм Clupea harengus, в отличие от подсем. Clupeinae и других видов этого подсемейства, у которых анальный плавник претерпевает перемещение в обратном направлении, чем спинной и брюшные. Однако в отличие от форм Clupea harengus расстояния между анальным и спинным и брюшными и анальным плавниками, наоборот, увеличиваются. С увеличением размеров головы, как у подсем. Clupeinae, ширина ее увеличивается.
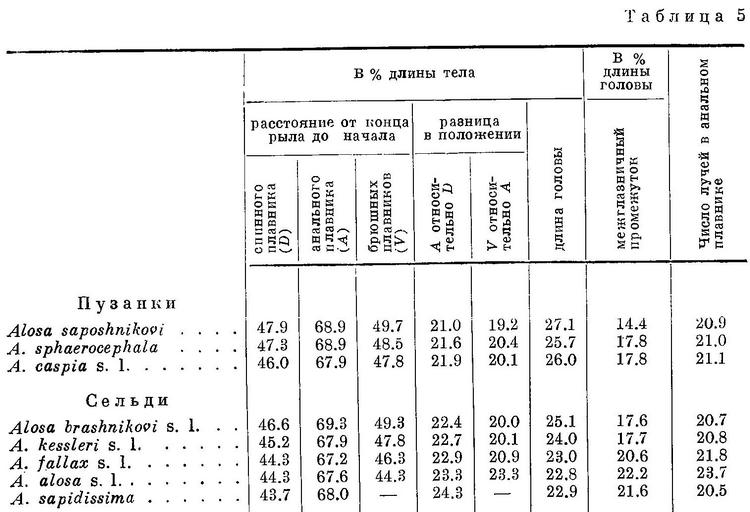
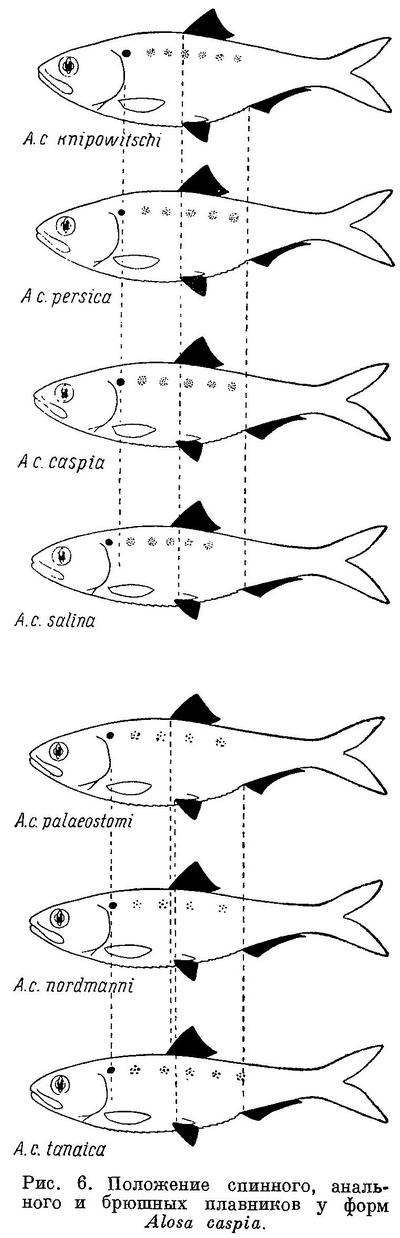
Виды рода Alosa делятся на две группы: на пузанков и сельдей. Виды из группы пузанков (Alosa saposhnikovi, Alosa sphaerocephala и Alosa caspia) по всем рассматриваемым особенностям являются более примитивными, чем виды из группы сельдей, из последних средиземноморско-атлантические виды (Alosa fallax, Alosa alosa, Alosa sapidissima) — наиболее высокоорганизованными. В связи с различием в форме тела пузанков и сельдей в положении спинного и анального плавников относительно переднего конца тела между обеими группами наблюдается некоторое захождение и параллельный характер изменения положения плавников в каждой группе, проявляющийся, как будет указано ниже, и у каспийских и у черноморско-азовских форм Alosa caspia. Однако разность в положении плавников относительно друг друга имеет непрерывный возрастающий характер за исключением разности в положении брюшного плавника относительно анального у Alosa caspia, что, невидимому, обусловлено наличием у него немигрирующих форм, локализованных в южной части моря.
У всех видов рода Alosa за жаберной крышкой сверху с каждой стороны тела имеется по темному пятну. Позади этого пятна у некоторых видов на боках тела есть еще по нескольку таких же пятен, расположенных в ряд. Из группы пузанков у Alosa saposhnikovi имеется только одно пятно за жаберной крышкой; у Alosa sphaerocephala, кроме того, бывает изредка несколько пятен и на боках тела, которые у Alosa caspia встречаются значительно чаще и в большем количестве. Далее, из сельдей у большинства форм Alosa brashnikovi есть только пятно за жаберной крышкой, но у некоторых наиболее специализированных форм этого вида иногда (Alosa brashnikovi nirchi) или обычно (Alosa brashnikovi grimmi) имеется, кроме того, по ряду расплывчатых темных пятен на боках тела. Из форм Alosa kessleri у одних (Alosa kessleri volgensis, мелкая форма Alosa kessleri pontica) имеется только по пятну за жаберной крышкой, у других (Alosa kessleri kessleri, крупная форма Alosa kessleri pontica) изредка есть по ряду из нескольких пятен и на боках тела. Наконец, у Alosa fallax, Alosa alosa и Alosa sapidissima на боках тела есть постоянно по ряду пятен, причем под этим рядом иногда имеется еще 1—2 ряда из меньшего числа пятен. Наличие пятен на боках тела у каспийско-черноморских сельдей ставилось в связь с сезоном, полом, зрелостью половых продуктов. Однако позднее (Дехтярева, 1940 : 61) установлено, что пятна на боках тела есть и у молоди тех видов, у которых они имеются во взрослом состоянии, и, следовательно, наличие их зависит не от указанных причин.
Как и у некоторых видов и подвидов подсем. Clupeinae, существенное значение, наряду с рассматриваемыми особенностями, имеет степень развития зубов. Берг (1913 : 12) на основании того, что у молоди видов Alosa зубы развиты сильнее, чем у взрослых, считает виды с хорошо развитыми зубами примитивными, виды с менее развитыми зубами — более высокоорганизованными. Мнение это стоит в полном согласии с предлагаемой системой рода за немногими лишь исключениями (Alosa kessleri kessleri, черноморско-азовские формы Alosa caspia). Из группы пузанков зубы наиболее сильно развиты у Alosa saposhnikovi, наименее — у всех форм Alosa caspia; Alosa sphaerocephala занимает промежуточное положение между этими видами. Далее, из группы сельдей зубы наиболее развиты у всех форм Alosa brashnikovi. У Alosa kessleri зубы развиты менее, особенно у Alosa kessleri volgensis, иногда почти незаметные; наиболее развиты они у Alosa kessleri kessleri. Наконец, у средиземноморско-атлантических видов Alosa fallax, Alosa alosa и Alosa sapidissima зубы наименее развитые из всех видов рода. У молоди этих видов, как и у упомянутых выше, зубы более развитые (ясно заметные на ощупь), чем у взрослых. Следует отметить, что у молоди зубы наиболее развиты у тех видов (Alosa saposhnikovi, Alosa brashnikovi и др.), у которых они сильнее всего выражены во взрослом состоянии, у молоди же видов со слабыми зубами они менее заметны (Дехтярева, 1940 : 58).
Изменение признаков, положенных в основу системы видов рода Alosa, в обоих группах, пузанков и сельдей, находятся в такой же связи с особенностями их образа жизни, как и у подсем. Clupeinae. Alosa saposhnikovi и Alosa sphaerocephala, у которых спинной и другие плавники наиболее отнесены назад и наименьшая разность в положении плавников относительно друг друга, всю жизнь проводят в море, не заходя в реки. Оба эти вида, судя по всему, совершают миграции в основном из средней части Каспийского моря в северную. Alosa caspia совершает более отдаленные миграции, в особенности его северокаспийские формы, идущие на нерест из южной части моря в северную, причем все черноморско-азовские формы и отчасти Alosa caspia caspia заходят в реки. Из сельдей наименее протяженные миграции совершают формы Alosa brashnikovi, большинство которых всю жизнь проводит в южной части Каспийского моря, и лишь две формы (Alosa brashnikovi brashnikovi, Alosa brashnikovi agrachanica) идут для размножения в северную часть его, не заходя в реки. Черноморско-азовская форма (Alosa brashnikovi maeotica), подобно двум последним, совершает миграции значительной протяженности из Черного моря и северо-западную часть его и в Азовское море. Проходные виды, у которых спинной и другие плавники расположены ближе к переднему концу тела и разность в положении плавников относительно друг друга наибольшая, совершают наиболее отдаленные миграции. Каспийские формы Alosa kessleri (Alosa kessleri volgensis, Alosa kessleri kessleri) из южной части моря идут для икрометания в реки северной части, черноморско-азовская (Alosa kessleri pontica) — из Черного моря в реки северо-западной части его и Азовского моря. Средиземноморско-атлантические виды (Alosa fallax, Alosa alosa, Alosa sapidissima) являются также проходными. Протяженность миграций их в море не известна, но в отношении Alosa sapidissima можно считать установленным, что эти миграции превосходят по протяженности таковые у Alosa kessleri.
Наиболее прогрессивные виды как из пузанков, так и из сельдей имеют и более широкий ареал, отличаясь вместе с тем и большей численностью.
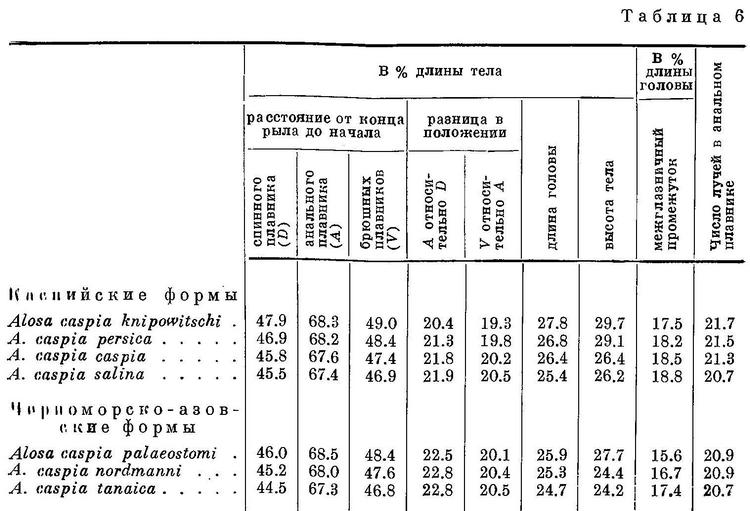
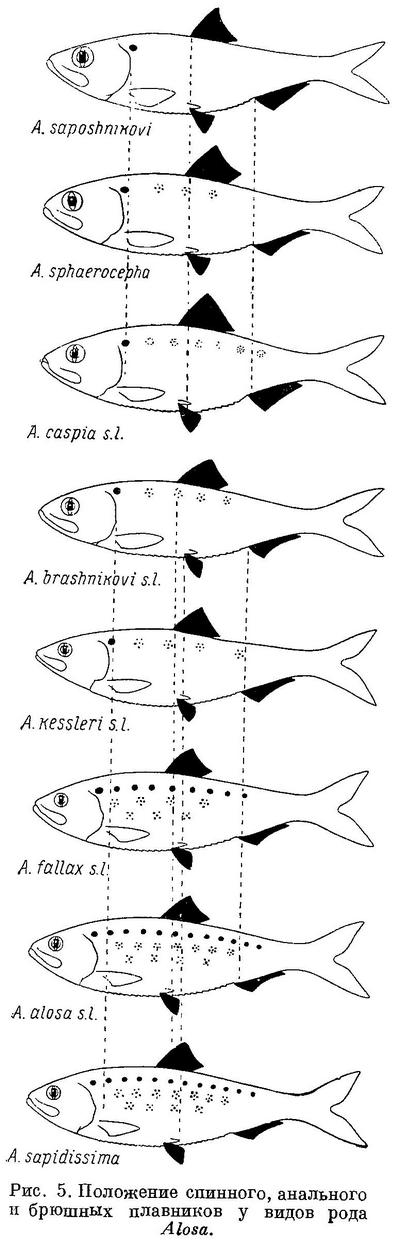
Изменение признаков, положенных в основу классификации, рассматривалось у видов рода Alosa. У подсем. Clupeinae, как указывалось выше, оно наблюдается и у подвидов в пределах видов. То же самое можно, установить у некоторых видов рода Alosa. Так, например, у Alosa caspia такая же зависимость, как и у видов рода, наблюдается в отношении почти всех рассматриваемых признаков, причем, как и у видов из группы пузанков и сельдей Alosa, изменение их, в частности изменение положении плавников, у каспийских и черноморско-азовских форм имеет некоторое захождение и параллельный характер. Однако в каждой из групп изменение положения плавников имеет характер закономерного перемещения их к передней части тела (табл. 6, рис. 6). При этом все признаки изменяются сходным образом, как и у видов рода. В сходной взаимосвязи находятся рассматриваемые признаки и с образом жизни форм Alosa caspia. Среди каспийских форм спинной и анальный плавники расположены далеко назад, разность в положении их относительно друг друга меньше и лучей в анальном плавнике больше у форм, живущих о размножающихся в южной части моря (Alosa caspia knipowitschi, Alosa caspia persica). У форм, зимующих в южной части моря, а для размножения идущих в северную часть моря (Alosa caspia caspia, Alosa caspia salina), спинной и анальный плавники расположены ближе к переднему концу тела, разность в положении их относительно друг друга больше и лучей в анальном плавнике меньше. Черноморско-азовские формы Alosa caspia соответственно в общем более близкому расположению плавников к переднему концу тела и пр., чем у каспийских форм, ведут в общем и более подвижный образ жизни, причем все они для размножения заходят в низовья рек, в озера и лиманы, и ни одна из них не мечет икру в море, между тем как большинство каспийских форм размножается в море, в заливах и ильменях или в устьях рек. Наименьшие по протяженности миграции из всех черноморско-азовских форм Alosa caspia совершает Alosa caspia palaeostomi, локализованный в южной части моря, наибольшие — Alosa caspia nordmanni и Alosa caspia tanaica; первые с мест зимовок от южных берегов Крыма и западных Черного моря идет в реки северо-западной части его, второй от южного кавказского побережья — в р. Дон и лиманы дельты Кубани. Помимо указанных признаков как у каспийских, так и у черноморско-азовских форм Alosa caspia сходным образом изменяется, высота тела, находящаяся также в зависимости от миграционных особенностей. Как видно из рис. 6 и табл. 6, у форм, совершающих миграции большей протяженности в северные части моря, тело значительно ниже, в особенности у каспийских, чем у форм, локализованных в южных частях моря. Что касается темных пятой на боках тела, то у черноморско-азовских форм они встречаются реже и в менее выраженной форме, чем у каспийских. Следует отметить, что у черноморско-азовских форм и других видов рода Alosa (Alosa kessleri pontica) пятна на боках тела развиты менее.
У Alosa kessleri рассматриваемые признаки изменяются непрерывно на протяжении всех форм вида (табл. 7, рис. 7), за исключением длины головы, изменение которой, как и у форм Alosa caspia, имеет параллельный характер соответственно делению форм вида на черноморско-азовских и каспийских (табл. 7, рис. 7). Что касается степени развития зубов, то они менее всего развиты у Alosa kessleri volgensis, более — у Alosa kessleri pontica и Alosa kessleri kessleri. Таким образом, степень развития зубов в данном случае не стоит в связи с изменением других признаков, что вполне понятно, так как эта особенность связана не с движением, а с питанием.
Судя по всем рассматриваемым признакам, за исключением степени развития зубов, наиболее высокоорганизованной формой вида следует считать Alosa kessleri kessleri. Это стоит в полном согласии и с тем, что пятна на боках тела из обеих каспийских форм имеет только эта. По более отнесенным назад плавникам, большим размерам и большей ширине головы и пр. наиболее примитивной формой Alosa kessleri следует считать Alosa kessleri pontica. При этом крупная форма Alosa kessleri pontica, имеющая изредка на боках тела темные пятна, соответствует биологически и отчасти морфологически Alosa kessleri kessleri, у которой пятна на теле бывают чаще, а мелкая форма — Alosa kessleri volgensis. Более высокой степени развития рассматриваемых признаков у Alosa kessleri kessleri по сравнению с Alosa kessleri volgensis и крупной формы Alosa kessleri pontica по сравнению с мелкой соответствуют особенности в распространении и биологии, подобные отмеченным у более высокоорганизованных видов рода Alosa и форм Alosa caspia. Alosa kessleri kessleri и крупная форма Alosa kessleri pontica заходят для нереста высоко вверх по течению рек, в то время как мелкая форма Alosa kessleri pontica и Alosa kessleri volgensis высоко в реки не поднимаются и нерестуют в нижнем течении, последняя иногда частично, даже в предустьевом пространстве Волги.
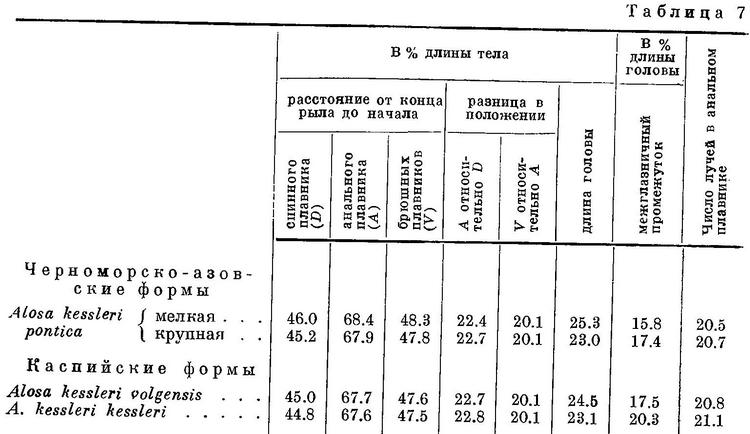
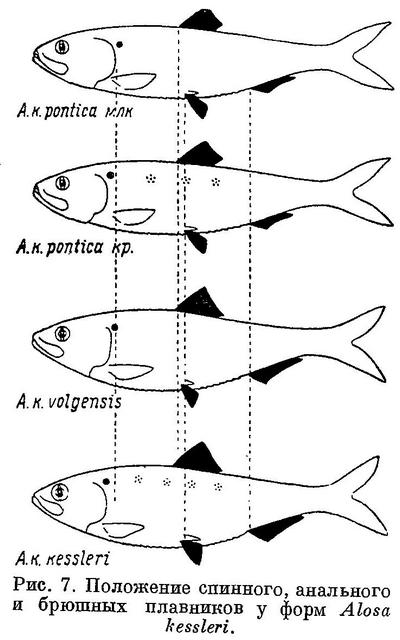
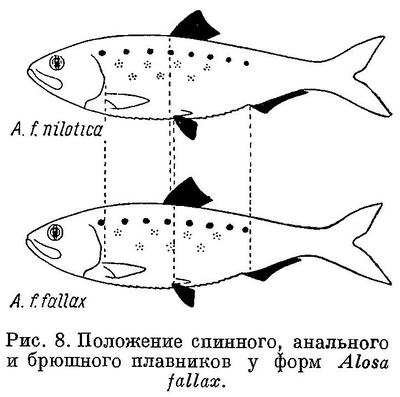
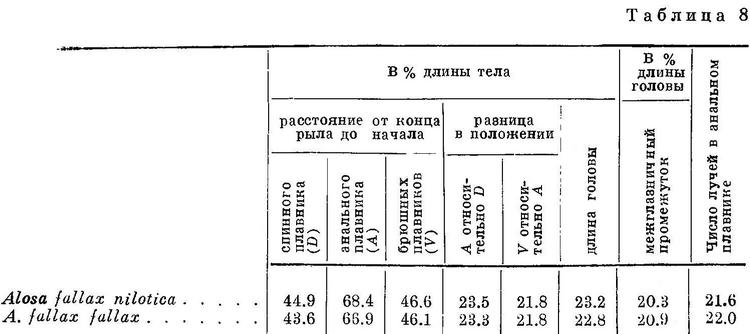
Из всех форм Alosa fallax и Alosa alosa представилась возможность исследовать лишь некоторые и притом на немногочисленном материале. Исследуемые признаки и этих форм обоих видов имеют в общем такие же закономерные изменения, как и у других видов рода (табл. 8, рис. 8).
Alosa brashnikovi, наиболее богатый формами из всех видов рода, обладающий, кроме того, некоторыми своеобразными биологическими адаптациями, крайне изменчив в основных, связанных с этими адаптациями морфологических особенностях. Формы этого вида, расположенные соответственно положению спинного плавника, почти не обнаруживают какой-либо закономерности в изменении других рассматриваемых признаков, почему фактический материал здесь не приводится. Однако следует отметить, что и здесь у форм, идущих для нереста в северную часть Каспийского моря (Alosa brashnikovi brashnikovi и Alosa brashnikovi agrachanica), спинной и анальный плавники расположены ближе к переднему концу тела, межглазничный промежуток шире, чем у других форм. Вместе с тем, у Alosa brashnikovi kisselevitshi, ведущей наиболее пелагический образ жизни, но живущей и размножающейся в южной части Каспийского моря, спинной плавник расположен еще ближе к переднему концу тела, а межглазничный промежуток так же широк, как у обеих упомянутых форм. Наиболее отнесены плавники назад и уже голова у южнокаспийских форм (Alosa brashnikovi orientalis, Alosa brashnikovi grimmi, Alosa brashnikovi nirchi), ведущих придонный образ жизни.
Морфологические особенности форм Alosa brashnikovi обусловлены приспособлениями их к различным условиям существования. Основным фактором образования форм этого вида является, повидимому, приспособление их главным образом к различному характеру питания, крайне разнообразному у форм этого вида, вследствие чего у них и наблюдаются большое разнообразие и различия в строении жаберных тычинок, рта и частей черепа, связанных с питанием. Наиболее резко обособленная форма вида, Alosa brashnikovi grimmi, является вместе с тем по образу жизни и характеру питания наиболее придонной из всех форм вида. Питается эта форма преимущественно бычками, иногда даже моллюсками (Cardium), реже тюлькой, в связи с чем жаберных тычинок у ней наименьшее количество, а на конце нижней челюсти у взрослых рыб имеется мозолистое утолщение, развивающееся, повидимому, вследствие соприкосновения нижней челюсти с грунтом при захватывании пищи. Весьма характерно, что у молодых рыб, длиной до 20 см, мозолистого утолщения на нижней челюсти нет. Повидимому, по этой же причине отсутствует у Alosa brashnikovi grimmi и бугорок на конце нижней челюсти, имеющийся почти постоянно у всех других форм вида. Вследствие механического воздействия грубой пищи жаберные тычинки искривляются, на конце утолщаются, расплющиваются, разветвляются и обламываются, а тычинки на нижнем конце жаберной дуги редуцируются и отсутствуют даже у молодых, благодаря чему первая тычинка у этой формы отстоит далеко от основания языка. Окраска у Alosa brashnikovi grimmi белесая, а на боках тела с каждой стороны, кроме пятна за жаберной крышкой, обычно имеется по ряду расплывчатых серых пятен, почти сливающихся в продольную полосу. Близкая к Alosa brashnikovi grimmi и ведущая подобно ей придонный образ жизни Alosa brashnikovi nirchi имеет иногда на нижней челюсти точно такое же, но небольшое мозолистое утолщение; окраска у этой формы бледная; на боках тела, помимо пятна за жаберной крышкой, иногда имеется по ряду расплывчатых серых пятен. Наиболее резко уклоняющаяся от этих форм, а в особенности от Alosa brashnikovi grimmi, Alosa brashnikovi kisselevitshi имеет самое большое количество жаберных тычинок из всех форм вида, причем тычинки у этой формы обычно заостренные, не утолщенные и не разветвленные на концах, первая тычинка расположена близко от основания языка. Держится эта форма в поверхностных слоях воды, питается тюлькой, креветками, бычками. Вследствие различий в характере питания наблюдаются изменения и в строении частей скелета головы Alosa brashnikovi grimmi. Часть черепа у этой формы в области прикрепления hyomandibulare, а посредством него и нижней челюсти (именно передний конец каждого sphenoticum) укорочена и сзади закруглена и не изогнута крючкообразно, как у других форм вида (например у Alosa brashnikovi brashnikovi, Alosa brashnikovi orientalis, Alosa brashnikovi kisselevitshi). Сходное строение имеет передний конец каждого sphenoticum и у Alosa brashnikovi nirchi. У этих форм наблюдаются существенные различия и в строении жаберных тычинок, подробно описанные в диагнозах.
Таким образом, из этих примеров видно, что наиболее специализированными формами Alosa brashnikovi следует считать Alosa brashnikovi grimmi и затем Alosa brashnikovi nirchi, приспособившихся к совершенно несвойственному сельдевым придонному образу жизни и преимущественно бентоядному питанию. Эта биологическая специализация сопровождается рядом глубоких морфологических изменений и появлением пятен на боках тела, свойственных, как указывалось выше, наиболее высокоорганизованным видам и формам. Спинные же и другие плавники расположены ближе к переднему концу тела, и разница в положении их относительно друг друга больше у форм, идущих для размножения в северную часть Каспийского моря (Alosa brashnikovi brashnikovi и Alosa brashnikovi agrachanica), чем у форм, постоянно живущих и размножающихся в южной части моря (Alosa brashnikovi sarensis, Alosa brashnikovi autumnalis), а среди последних менее всего у ведущих придонный образ жизни (Alosa brashnikovi grimmi, Alosa brashnikovi nirchi). Среди южнокаспийских форм Alosa brashnikovi спинной и анальный плавники расположены ближе всего к переднему концу тела и разность в положении этих плавников относительно, друг друга больше у наиболее пелагической формы (Alosa brashnikovi kisselevitshi).
Естественно, что наиболее прогрессивные по рассматриваемым морфологическим особенностям формы Alosa caspia и Alosa brashnikovi, идущие для икрометания в северную часть Каспийского моря, занимают и более широкий ареал, чем формы, локализованные в южной части моря. Вместе с тем эти формы характерны и большей численностью.
У подсем. Dorosomatinae основание анального плавника несколько более удлиненное и плавник имеет большее количество лучей, чем у рассмотренных выше подсем. Clupeinae и отчасти Alosinae, спинной плавник, наоборот, в общем немного короче и с меньшим количеством лучей. У родов подсемейства по мере увеличения числа лучей в анальном плавнике число лучей в спинном плавнике уменьшается (табл. 9). Количество лучей в анальном и спинном плавниках в табл. 9 и 10 приведено по работам, цитированным при описании обоих подсемейств в систематической части. У родов Dorosoma, Gonialosa и Signalosa, имеющих наибольшее число лучей в анальном плавнике и наименьшее в спинном, за жаберной крышкой сверху с каждой стороны тела есть по большому темному пятну. У родов Clupanodon, Nematalosa и Anodontostoma с наименьшим числом лучей в анальном плавнике и наибольшим в спинном, кроме пятна за жаберной крышкой, имеются ряды темных более мелких пятен вдоль рядов чешуи на верхней половине тела. Последние роды имеют более короткое расстояние от конца рыла до начала спинного плавника (соответственно перечисленным выше родам 42.2, 44.8 и 45.3%) и более длинное до начала анального (соответственно 66.7, 68.8 и 71.1%) и брюшных (44.0, 45.0, 46.1%), чем у Dorosoma (до начала спинного 47.2, анального 57.0,
брюшного 57.9%). Отсутствие материалов по некоторым родам подсемейства не дает возможности расположить их соответственно положению плавников, но, судя по всему, роды Clupanodon, Nematalosa и Anodontostoma, имеющие наиболее развитую пигментацию на теле, должны, как и сходные роды в подсем. Clupeinae, занять среднюю часть ряда. Представители подсем. Dorosomatinae — проходные, пресноводные и отчасти морские рыбы, держащиеся в заливах и эстуариях. Образ жизни их изучен мало, и поэтому в настоящее время затруднительно установить связь между рассматриваемыми морфологическими и биологическими особенностями.
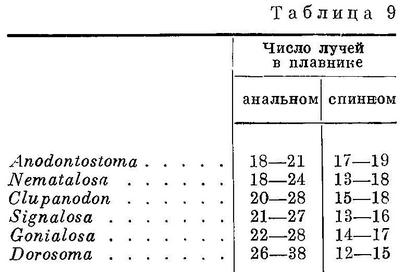

Подсем. Pristigasterinae характерно еще более удлиненным основанием анального плавника с большим числом лучей в нем и слабым развитием брюшных и спинного плавников, причем оба последних плавника у некоторых родов отсутствуют. По мере увеличения числа лучей в анальном плавнике число их в спинном, как это отмечено и у подсем. Clupeinae и Dorosomatinae, уменьшается (табл. 10), и, наконец, у рода Raconda, имеющего наибольшее число лучей в анальном плавнике, спинной плавник вообще отсутствует. Параллельно с этим брюшные плавники, имеются лишь у родов Ilisha, Pellona и Chirocentrodon, редуцируются и у остальных родов отсутствуют. Роды подсем. Pristigasterinae в таблице расположены соответственно увеличению числа лучей в анальном плавнике и уменьшению числа их в спинном. Возможно, что в зависимости от величины антедорсального расстояния расположение их будет иное и даст, как в подсем. Clupeinae, возрастание одних и убывание других к середине ряда и соответственно убывание и возрастание к концу. Биология представителей подсем. Pristigasterinae не изучена, и поэтому установить взаимосвязь между рассматриваемыми морфологическими особенностями и образом жизни не представляется возможным.
В заключение естественно задать вопрос, каково значение различий в положении плавников, числа лучей в них, ширины головы и других признаков, которые, как указывалось, теснейшим образом связаны с биологическими особенностями — с протяженностью миграций и жизнью на разных глубинах и на разной отдаленности от берегов?
Функция плавников, в частности непарных, выяснена еще недостаточно. По наблюдениям Васнецова (1941 : 503), спинной плавник, расположенный, как у преобладающего большинства сельдевых, на уровне или немного сзади центра тяжести, служит для сохранения прямолинейности движения и для фиксации положения середины тела при поворотах. Анальный плавник, расположенный у всех костистых рыб в задней части тела сзади уровня центра тяжести тела, служит для подъема и опускания передней, части тела, так как при помощи его соответственно изменяется положение задней части тела в обратном направлении. Удаление его влечет за собой затруднение или неспособность к опусканию головы вниз. Такую же роль выполняет и спинной плавник, расположенный сзади уровня центра тяжести тела рыбы.
Более детально этот вопрос не изучен. В частности, не выяснено значение различия во взаимном положении спинного и анального плавников, разница в положении которых по отношению друг к другу изменяется, как указывалось, более закономерно и, имеет большую абсолютную разность между крайними значениями ее, чем между крайними положениями этих плавников относительно передней части тела. На основании того, что изложено выше, об этом можно только догадываться и высказывать предположения. У представителей подсем. Clupeinae, совершающих миграции наибольшей протяженности и живущих в основном в более поверхностных слоях воды (Sardinops, Sardina, Sardinella), анальный плавник отнесен назад, спинной еще в большей степени вперед и поэтому разница во взаимном расположении этих плавников наиболее значительна (см. табл. 1). У представителей того же подсемейства, не совершающих таких миграций и живущих в основном на больших глубинах (Sprattus, Clupea, Clupeonella), наоборот, анальный плавник расположен ближе к переднему концу тела, спинной более удален от него, чем у упомянутых выше родов, и поэтому разница во взаимном расположении их значительно меньше.
Равным образом и у форм Sprattus sprattus и Clupeonella delicatula разница в положении спинного и анального плавников относительно друг друга увеличивается параллельно уменьшению расстояния спинного плавника от конца рыла. О биологических отличиях этих форм сказано выше.
Большее значение положения спинного и анального плавников относительно друг друга, чем относительно переднего конца тела, нагляднее видно у видов рода Alosa. Несмотря на то что положение анального плавника у видов этого рода в отличие от подсем. Clupeinae изменяется в том же направлении, что и спинного, тем не менее разница во взаимном положении этих плавников, по мере уменьшения расстояния спинного плавника от конца рыла, как указывалось, увеличивается (табл. 5). Виды, имеющие менее отнесенные назад спинной и анальный плавники и, что самое существенное, большую разницу во взаимном положении их, совершают более отдаленные миграции. Наибольшую разницу в положении этих плавников относительно друг друга имеют виды из группы сельдей, наименьшую — виды из группы пузанков, среди последних наибольшую Alosa caspia, некоторые формы которого заходят для икрометания в реки, среди первых — проходные виды. Об этом свидетельствуют также и формы некоторых видов Alosa, в частности Alosa caspia (см. табл. 6): у черноморско-азовских форм, заходящих для размножения в реки, расстояние между началом спинного и анального плавников больше, чем у каспийских, а среди последних расстояние это больше у форм, идущих для икрометания в северную часть моря, чем у форм, живущих в южной части моря, не выходящих за пределы его.
Наконец, еще более убедительный пример большего значения положения спинного и анального плавников относительно друг друга, чем относительно переднего конца тела, представляют формы Clupea harengus (табл. 4). Изменение положения анального плавника по отношению к переднему концу тела у форм этого вида в отличие от видов подсем. Clupeinae и форм других видов его имеет такой же характер, как и изменение положения спинного. Однако разница во взаимном положении этих плавников по мере уменьшения расстояния от конца рыла до начала спинного плавника не увеличивается, как у всех рассматриваемых выше сельдевых, а уменьшается, что стоит в полном соответствии с размерами миграций форм Clupea harengus. Clupea harengus harengus, имеющий наибольшую разницу в положении этих плавников, совершает, как указывалось выше, и большие по размерам миграции, чем другие формы, у которых разница во взаимном положении этих плавников меньше. К сходному выводу автор пришел и относительно тресковых, у которых виды, имеющие большие промежутки между спинными и анальными плавниками, являются более подвижными.
Таким образом, из приведенных примеров видно, что большее значение и большую связь с другими рассматриваемыми морфологическими особенностями, а также и с биологическими имеет положение спинного и анального плавников на теле не столько по отношению к переднему концу тела, сколько относительно друг друга. Виды и формы, имеющие большую разницу во взаимном положении спинного и анального плавников, совершают более отдаленные миграции и являются более подвижными.
Еще менее изучено значение изменения числа лучей в спинном и анальном плавниках, характеризующее длину основания их. Из всех рассматриваемых примеров можно сделать заключение, что увеличение числа лучей в анальном плавнике идет параллельно уменьшению числа их в спинном. Особенно наглядно видно это у тех из рассматриваемых групп, где число лучей в плавниках велико — в подсем. Dorosomatinae и особенно Pristigasterinae. Судя по характеру изменения числа их у представителей подсем. Clupeinae, можно полагать, что оно также теснейшим образом связано с движением, так как число лучей в анальном плавнике менее всего у видов, совершающих большие по протяженности миграции, однако у видов Alosa — наоборот. В частности, такую же зависимость между размерами анальных плавников, в особенности первого анального плавника, и подвижностью обнаружена автором у тресковых (Световидов, 1948 : 27). Наиболее подвижный образ жизни ведут виды, имеющие более короткий первый анальный плавник и располагающиеся в середине ряда подсем. Gadinae. Виды, живущие на больших глубинах и ведущие менее подвижный образ жизни, имеют длинный первый анальный плавник.
Функция брюшных плавников менее ясна, чем спинного и анального. По указанию Васнецова (1941 : 50), эти плавники являются рулями глубины. Вместе с тем, по исследованиям того же автора, при удалении брюшного плавника тело рыбы несколько смещается в сторону, что указывает на роль этих плавников в фиксации середины тела на месте при поворотах. Значение же положения брюшных плавников на теле рыбы относительно переднего конца тела и тем более относительно непарных плавников совершенно не изучено. Судя по более близкому положению брюшных плавников к переднему концу тела и большему удалению от анального у рыб, совершающих отдаленные миграции, можно полагать, что эти плавники имеют отношение и к движению.
В теснейшей связи с рассматриваемыми морфологическими и биологическими особенностями сельдевых находятся и размеры головы, значительно более широкой у видов и форм отдельных видов с более отнесенным вперед спинным плавником, большей разницей в положении относительно него анального и совершающих большие миграции. Это вполне понятно, так как среди пелагических рыб многие из совершающих отдаленные миграции, например кефали, имеют торпедообразную форму тела с расширенной головой. Что касается длины головы, не имеющей, повидимому, функциональной связи с движением, то увеличение ее в связи с уменьшением ширины, можно полагать, обусловлено коррелятивной изменчивостью.
Степень развития сети каналов системы боковой линии на голове у видов подсем. Clupeinae находится также в связи с движением, так как они более развиты у видов, совершающих отдаленные миграции, чем, у видов с меньшими размерами миграций.
Таким образом, параллельный характер изменения рассматриваемых морфологических особенностей находится в теснейшей связи с биологическими особенностями, с размерами совершаемых ими миграций.
Эта взаимная связь выражается, с одной стороны, в положении спинного, анального и брюшных плавников на теле как относительно переднего конца его, так в основном относительно друг друга, в различном числе лучей в спинном и анальном плавниках, характеризующем их длину, в большей или меньшей ширине головы и у некоторых сельдевых и высоте тела, характеризующими в совокупности различную степень приспособления к движению. С другой стороны, как это видно на примере подсем. Clupeinae, эта связь выражается в различной степени развития каналов системы боковой линии на голове, характеризующей приспособление к ощущению движения. Следовательно, комплекс рассматриваемых морфологических особенностей заключает приспособление к движению — к различной протяженности миграций и к ощущению движения.
Несколько особняком стоит изменение пигментации, не имеющей приспособительного характера к движению. Однако параллельное изменение интенсивности ее в связи с изменением комплекса рассматриваемых признаков резко бросается в глаза, имеет не случайный характер и обусловлено не прямым отношением к движению, а косвенными причинами. Как известно, наиболее сильно изменяется окраска большинства рыб, в частности лососевых, в период наиболее интенсивной их жизнедеятельности, во время икрометания и, повидимому, стимулируется не только созреванием половых продуктов, но и усиленным обменом веществ в связи с совершаемыми ими в этот период отдаленными миграциями. Пигментация более развита, как указывалось, у тех сельдевых, которые совершают отдаленные миграции, и развитие пигментных клеток, вероятно, стимулировано усиленным обменом веществ этих сельдевых. Таким образом, судя по всему, и различие в развитии пигментации сельдевых стоит в связи с движением, с размерами совершаемых ими миграций.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
Современное распространение семейства сельдевых, как и распространение основной массы водной и наземной фауны, помимо причин исторических, обусловлено главным образом их экологическими особенностями, в частности отношением к температуре.
Преобладающее большинство представителей семейства по своему распространению является тропическим. Из 50 родов и около 190 видов сельдевых распространение почти 150 видов, относящихся к 37 родам, ограничено тропиками, причем некоторые виды ряда родов (Harengula zunazi, Sardinella aurita, Ilisha elongata и др.) заходят в южные части субтропических морей.
Умеренным широтам свойственна сравнительно небольшая часть сельдевых, в которую входят роды, распространенные в бореальной и в большей части субарктической областях Дерюгина (1915 : 717; 1924 : 96; 1928 : 352). Здесь встречается около 20 видов, относящихся к 7 родам, причем виды некоторых родов или формы некоторых видов свойственны более южным широтам, субтропикам (Sprattus sprattus phalericus, Alosa fallax nilotica и ряд пресноводных форм этого вида из бассейна Средиземного моря, Alosa alosa africana и некоторые пресноводные и проходные формы этого вида из бассейна Эгейского моря и юго-западной части Черного) и даже отчасти тропикам (Pomolobus chrysochloris, Brevoortia tyrannus patronus).
Незначительная часть сельдевых по своему распространению занимает промежуточное положение между тропическими и бореальными, являясь в основном субтропическими. Не считая некоторых упомянутых выше форм бореальных видов и некоторых заходящих сюда видов тропических родов, в субтропических морях северного полушария встречаются лишь 4 вида, относящиеся к 3 родам семейства (Etrumeus, Sardina, Sardinops), причем 2 из этих видов относятся к двум родам (Sardina и Sardinops). Формы Sardinops свойственны субтропическим морям и южного полушария, одна из форм Sardina (Sardina pilchardus pilchardus) встречается в умеренных морях. Из 4 видов рода Etrumeus один (Etrumeus sadina) свойствен субтропическим и прилегающим тропическим частям морей северного полушария (от Мексиканского зал. до м. Код), второй (Etrumeus jacksoniensis) — южного; третий вид этого рода (Etrumeus micropus) является отчасти тропическим (Гавайские о-ва), но в основном встречается также в субтропических морях северного (у берегов Японии, в зал. Петра Великого, у Калифорнии) и южного (у южных берегов Африки) полушарий. Скорее субтропическим следует считать также один из видов рода Dorosoma (Dorosoma cepedianum cepedianum — от м. Код до Техаса, Dorosoma cepedianum exile — Техас).
Арктических среди сельдевых нет ни одного рода и вида. В арктических морях встречаются лишь некоторые подвиды Clupea harengus (Clupea harengus pallasi n. maris-albi, Clupea harengus pallasi n. suworowi и отчасти Clupea harengus pallasi), являющиеся умеренноарктическими (Niederarktische по терминологии Hofsten, 1915 : 202). Высокоарктических (Hocharktische, по Hofsten, 1915) среди сельдевых нет.
В южном полушарии южнее тропиков встречается значительно меньшее количество сельдевых, чем в соответствующих частях северного. Умеренным и отчасти субтропическим морям южного полушария свойствен встречающийся и в северном полушарии род Sprattus, некоторые из 5 известных здесь видов которого, судя по всему, являются лишь подвидами. 2 вида этого рода (Sprattus arcuatus, Sprattus melanostoma) встречаются у берегов Южн. Америки, 3 — у южных берегов Австралии (Sprattus bassensis) и Новой Зеландии (Sprattus muelleri, Sprattus antipodum). В основном субтропическим в южном полушарии является единственный вид рода Potamalosa (Potamalosa richmondia, пресные воды Австралии), точно так же как и оба вида рода Hyperolophus, причем распространение одного из них (Hyperolophus translucidus, берега Нового Южн. Уэльса) ограничено самыми южными частями прилегающих тропических морей. Упомянутые выше для субтропических морей северного полушария роды Etrumeus и Sardinops в субтропических морях южного полушария встречаются у берегов южной Африки (Etrumeus micropus, Sardinops sagax ocellata), Южн. Америки (Sardinops sagax sagax), Австралии (Etrumeus jacksoniensis, Sardinops sagax neopilchardus) и Новой Зеландии (Sardinops sagax neopilchardus). Субтропическим морям южного полушария свойственно также несколько видов в основном тропического рода Spratelloides, встречающихся здесь у берегов южной Африки (Spratelloides aestuarius) и Австралии (Spratelloides robustus). В субтропиках южного полушария встречаются некоторые и тропические виды как упомянутых родов (например Spratelloides delicatulus), так и ряда других (например Ilisha natalensis, Hilsa durbanensis — у берегов южной Африки, Nematalosa richardsoni — южная Австралия, некоторые виды широкораспространенных родов Harengula, Sardinella). В антарктических частях южного полушария сельдевых нет.
Таким образом, основная масса сельдевых — тропические рыбы; бореальных и субтропических среди сельдевых значительно меньше, арктических почти нет; в высокоарктической подобласти сельдевые отсутствуют.
Второй экологической особенностью, обусловившей распространение сельдевых, следует считать их отношение к солености. По этой экологической особенности сельдевые представляют довольно смешанную группу. Смешанный характер по отношению к солености имеет не только семейство сельдевых в целом, но, как будет указано ниже, и отдельные роды и даже виды.
Основная масса сельдевых, около 25 родов, заключающих в общей сложности свыше 100 видов, является морскими рыбами тропических, умеренных и отчасти арктических морей. Сюда относятся роды с наибольшим количеством видов, как, например, Harengula, Sardinella. Некоторые виды в основном морских родов заходят в пресную воду, в устья рек и даже в реки и озера (например некоторые формы Clupea harengus, в особенности Clupea harengus pallasi, Ilisha indica и другие виды этого рода), некоторые роды (Clupeonella) или виды и подвиды морских родов живут в солоноватой воде (например некоторые подвиды Sprattus sprattus) или проводят всю жизнь в пресной воде.
Значительная часть сельдевых (около 15 родов, примерно с 30 видами) является пресноводными рыбами, причем распространение преобладающего большинства их ограничено тропиками. Большая часть их, 8 родов (Pellonula и близкие роды из подсем. Pellonulinae) с 14 видами, населяют пресные воды западной Африки. Ряд родов того же подсемейства с 6 видами встречается в пресных водах Индо-малайского архипелага (Clupeoides, Corica, Clupeichthys) и Австралии (Potamalosa). Несколько родов (Gudusia, Gonialosa, подсем. Alosinae) с 4 видами свойственны пресным подам Индии. Все виды рода Signalosa (подсем. Dorosomatinae) живут в пресных водах атлантического побережья центральной и тропической части Сев. Америки. Из 4 видов рода Dorosoma 3 связаны в основном с пресными водами; один вид этого рода (Dorosoma cepedianum), имеющий более северное распространение, живет в пресной и солоноватой воде, реже встречается в морской, будучи отчасти, повидимому, проходным. Пресноводными являются также некоторые виды ряда морских родов (например Rhinosardinia amazonica — р. Амазонка и реки Гвианы, Ilisha macrogaster — реки Борнео, Harengula tawilis — пресноводное озеро на Филиппинских о-вах и некоторые другие). Из пресноводных сельдевых более северных широт следует отметить Clupeonella abrau из пресноводного озера Абрау. Из остальных морских, точнее солоноватоводных, видов этого рода один (Clupeonella delicatula), будучи в равной мере морским и проходным и отчасти пресноводным, образует пресноводную морфу (Clupeonella delicatula caspia m. tscharchalensis). Ряд пресноводных форм образует виды проходных сельдей из родов Alosa и Pomolobus. Таковы из рода Alosa Alosa alabamae ohiensis, Alosa alosa macedonica, Alosa fallax lacustris, Alosa fallax gracilis и Alosa fallax killarnensis, — все, за исключением последней, умеренной формы, субтропические. Из рода Pomolobus местами в пресной воде живут постоянно проходные Pomolobus chrysochloris и Pomolobus pseudoharengus.
Проходных среди сельдевых, несколько меньшее количество, чем морских, и по количеству видов столько же, сколько и пресноводных. К числу их относится около 7 родов не более чем с 30 видами. Значительное количество проходных сельдей, в отличие от пресноводных, встречается в умеренных широтах. Таковыми являются роды Alosa (4 или 5 проходных и 4 или 5 морских, солоноватоводных) и Pomolobus (4 вида), причем некоторые проходные формы Alosa живут и размножаются в субтропиках (например Alosa fallax nilotica, Alosa alosa africana). Из каспийско-черноморских видов Alosa, помимо типично проходного вида Alosa kessleri, имеется Alosa caspia, одни формы которого размножаются в море, другие — в реках или устьях рек. Из сельдевых умеренных широт следует отметить упомянутый выше вид — Clupeonella delicatula, который размножается как в море, так и в реках. К сельдевым более южных широт относится род Brevoortia, относящийся, судя по всему, к солоноватоводным рыбам. Один вид этого рода (Brevoortia pectinata) встречается в умеренных водах южного полушария (от северной Патагонии до южной Бразилии), второй — в умеренных водах северного полушария (Brevoortia smithi — в эстуариях Сев. Каролины), третий — как в умеренных водах Сев. Америки (Brevoortia tyrannus tyrannus и Brevoortia tyrannus brevicauda), так почти и в тропических (Brevoortia tyrannus patronus в Мексиканском зал.). Из тропических сельдевых проходными являются некоторые виды рода Hilsa (Hilsa reevesii, Hilsa macrura) и, возможно, остальные 5 видов этого рода. Проходными также, вероятно, являются роды Nematalosa и Anodontostoma. Образ жизни ряда других тропических сельдевых не известен (например Ethmidium, Ethmalosa), но возможно, что некоторые из них тоже проходные. Имеющиеся для ряда сельдевых указания на их отношение к солености носят крайне неопределенный характер. В частности, это особенно относится ко многим видам рода Ilisha.
Таким образом, большинство сельдевых — морские рыбы, значительно меньшая часть их, будучи тесно связана с морем, — проходные; приблизительно такое же количество сельдевых по числу видов, как и проходных, но большее по числу родов — пресноводные. Во всех широтах, где встречаются сельдевые, преобладающая масса их является морскими. Таковы все из немногих сельдевых, встречающихся в арктических широтах. Сельдевые умеренных широт — морские или проходные; лишь некоторые из них, упомянутые выше, пресноводные, причем гораздо большее количество пресноводных форм образуют проходные умеренные сельдевые в субтропиках, чем в умеренных широтах. Пресноводными из умеренных и субтропических сельдевых в основном являются подвиды и лишь единичные виды. Ни одного рода, обитающего в пресных водах, здесь нет. Основная масса пресноводных сельдевых свойственна тропикам, где встречаются как ряд родов, все виды которых пресноводные, так и значительное количество пресноводных видов морских родов. В тропиках встречается значительное количество и проходных сельдей. Следовательно, при относительном все же преобладании морских количество пресноводных сельдевых в тропических областях наибольшее, что стоит в полном согласии с фактами, известными и для других групп животных.
Следует отметить, что морской или пресноводный образ жизни свойствен целым крупным подразделениям семейства — подсемействам. Морскими в основном, за редкими исключениями, являются подсемейства Dussumieriinae, Clupeinae и Pristigasterinae, в основном пресноводными — Pellonulinae. Представители подсемейства Alosinae и Dorosomatinae — проходные или пресноводные и отчасти солоноватоводные и морские.
Рассмотрение распределения сельдевых по зоогеографическим областям представляет некоторые трудности вследствие указанного выше неоднородного экологического характера их по отношению к солености. Однако основная масса сельдевых, как указывалось выше, является морскими. Если же принять во внимание и проходных, значительную часть своей жизни проводящих в море, то следует считать, что распространение преобладающего большинства представителей семейства теснейшим образом связано с морем. Поэтому в основу зоогеографического анализа сельдевых взяты морские и проходные. Пресноводные рассматриваются попутно соответственно зоогеографическим подразделениям, разработанным для пресноводных рыб Бергом (1909 : 242 и сл.).
Зooгеографические подразделения для морской фауны пелагической и для прибрежной (литоральной и неритической) существенно различаются. Сельдевые, как известно, являются рыбами пелагическими, однако по распространению и образу жизни относятся скорее к неритической фауне, чем к океанической. Действительно, в открытых частях океанов, на большом отдалении от берегов сельдевые обычно не встречаются. Распространение большинства представителей семейства связано с берегами. Многие из сельдевых вообще держатся в прибрежной зоне, в заливах и эстуариях. Даже наиболее пелагические и подвижные из них во время размножения подходят к берегам, в заливы и т. п. (Clupea harengus, Sardina pilchardus, Sardinella aurita, Sprattus sprattus и др.). Поэтому анализ распределения сельдевых по зоогеографическим областям произведен по зоогеографическим подразделениям для прибрежной фауны.
Из всех зоогеографических подразделений моря (Ortmann, 1895; Dahl, 1921 и др. и на основе их Гептнер, 1936) наиболее полно разработанное предложил Экман (Ekman, 1935). Границы основных подразделений Экмана, называемых им фаунами, наиболее близко совпадают с границами областей и подобластей Ортманна (Ortmann, 1895) для литоральной фауны. Однако подразделения Экмана, основанные на новейших исследованиях, значительно более детальны, в частности в отношении южного полушария южнее тропиков, как это видно из нижеследующего сопоставления:

Зоогеографические подразделения, относящиеся к тропическому и арктическому поясам, у обоих авторов одинаковы. Фауна умеренных частей Атлантического океана разделяется Экманом на европейскую бореальную и американскую североатлантическую, Тихого океана — на умеренные северо-западноамериканскую и северо-восточноазиатскую. Фауна южного полушария южнее тропиков, рассматриваемая Ортманном как единая антарктическая область, Экманом разбивается на ряд подразделений. Фауна сельдевых южного полушария южнее тропиков, как указывалось выше, значительно беднее фауны соответствующих частей северного полушария и в связи с этим детальное подразделение южного полушария, как увидим ниже, является для сельдевых излишним. Поэтому зоогеографический анализ сельдевых дается в основном по подразделениям Ортманна. Лишь значительное богатство сельдевых в умеренных частях Атлантического океана дает основание для разделения атлантической бореальной подобласти Ортманна на атлантическо-европейскую и атлантическо-американскуто бореальные подобласти. Бедная фауна сельдевых соответствующих частей Тихого океана оснований для подобного разделения не дает. Границы средиземноморской подобласти Ортманна в нижеследующем приняты несколько шире, по Экману, включая сюда фауну прилегающей части и Атлантического океана и понтическую фауну сельдевых Черного и Каспийского морей. В таком понимании эту подобласть правильнее называть средиземноморско-атлантической. Экман называет ее средиземноморско-атлантической и сарматской, что не совсем правильно, так как, точнее говоря, последняя фауна есть не сарматское, а понтическое наследие. Что касается границ рассматриваемых областей, то они у обоих авторов в основном совпадают. Вообще же говоря, как справедливо отмечает Экман (Ekman, 1935 : 144), это будут не границы, а скорее переходные районы областей и подобластей, так как морская фауна не имеет таких резких границ, как пресноводная.
Преобладающая масса сельдевых, как указывалось выше, сосредоточена в тропиках. При этом более двух третей общего числа тропических морских и проходных сельдевых, встречающихся в тропических областях, или почти половина общего количества всех морских и проходных видов свойственны индо-пацифической области (см. табл. 11. В таблицу не включены некоторые виды, систематическое положение которых неясно (например Clupea fuegensis, Clupea bentincki, Sardinops dakini, см. о них в сравнительных замечаниях родов).). Здесь, помимо большого количества видов, встречается наибольшее количество и родов, распространение которых ограничено пределами только этой области. Некоторые более широко распространенные роды (Spratelloides, Harengula, Sardinella, Nematalosa, Ilisha), встречающиеся и за пределами индо-пацифической области, преобладающим количеством видов представлены также здесь, причем необходимо отметить, что эти роды относятся к числу наиболее богатых видами. В реках, впадающих в моря индо-пацифической области и относящихся к сино-индийской области Берга, встречаются, кроме того, 14 видов пресноводных сельдевых, принадлежащих к 5 эндемичным для этой области родам (Clupeoides, Corica, Clupeichtys, Gudusia, Gonialosa). Таким образом, индо-пацифической морской области и относящейся к системе ее сино-индийской пресноводной области свойственно около 88 видов сельдевых, что составляет половину общего числа видов сельдевых, встречающихся в тропических областях. Эти виды относятся к 20 родам, из которых 10 эндемичные (помимо перечисленных пресноводных, Dussumieria, Kowala, Hilsa, Clupanodon, Raconda).
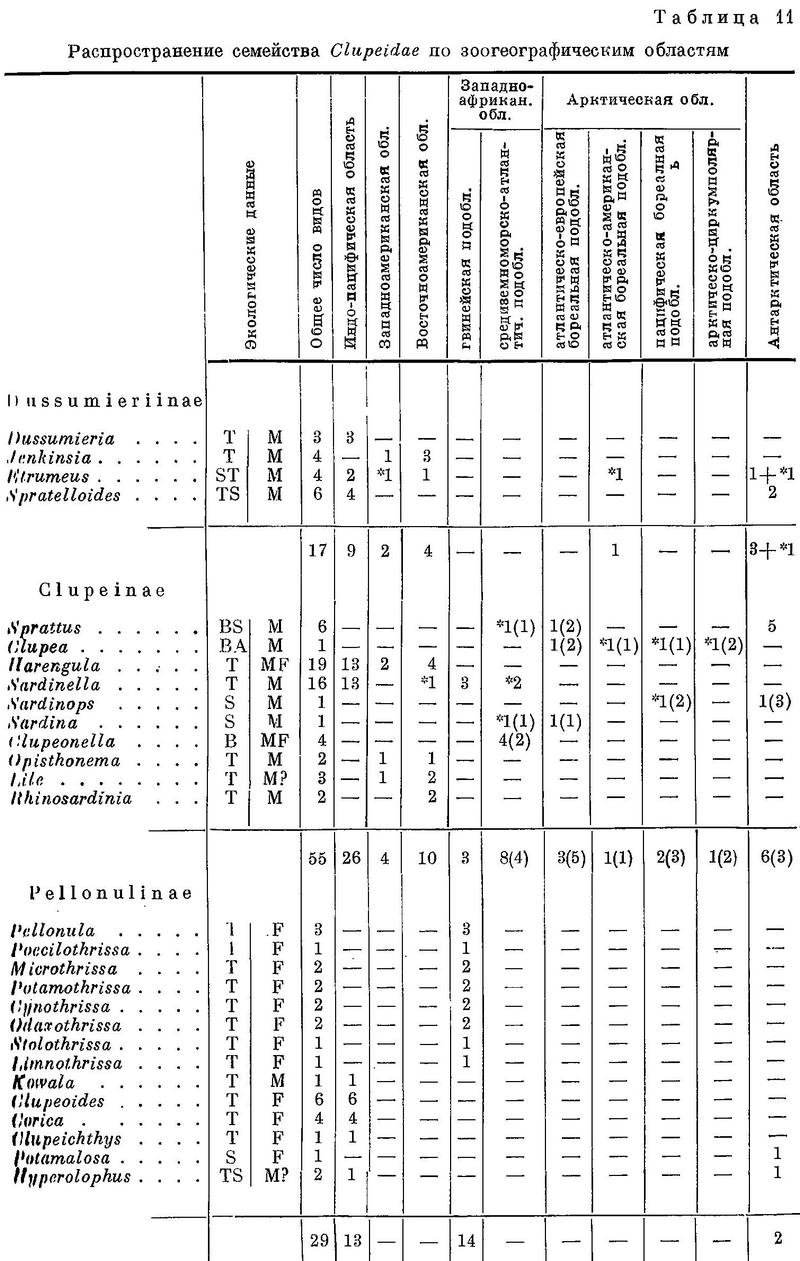
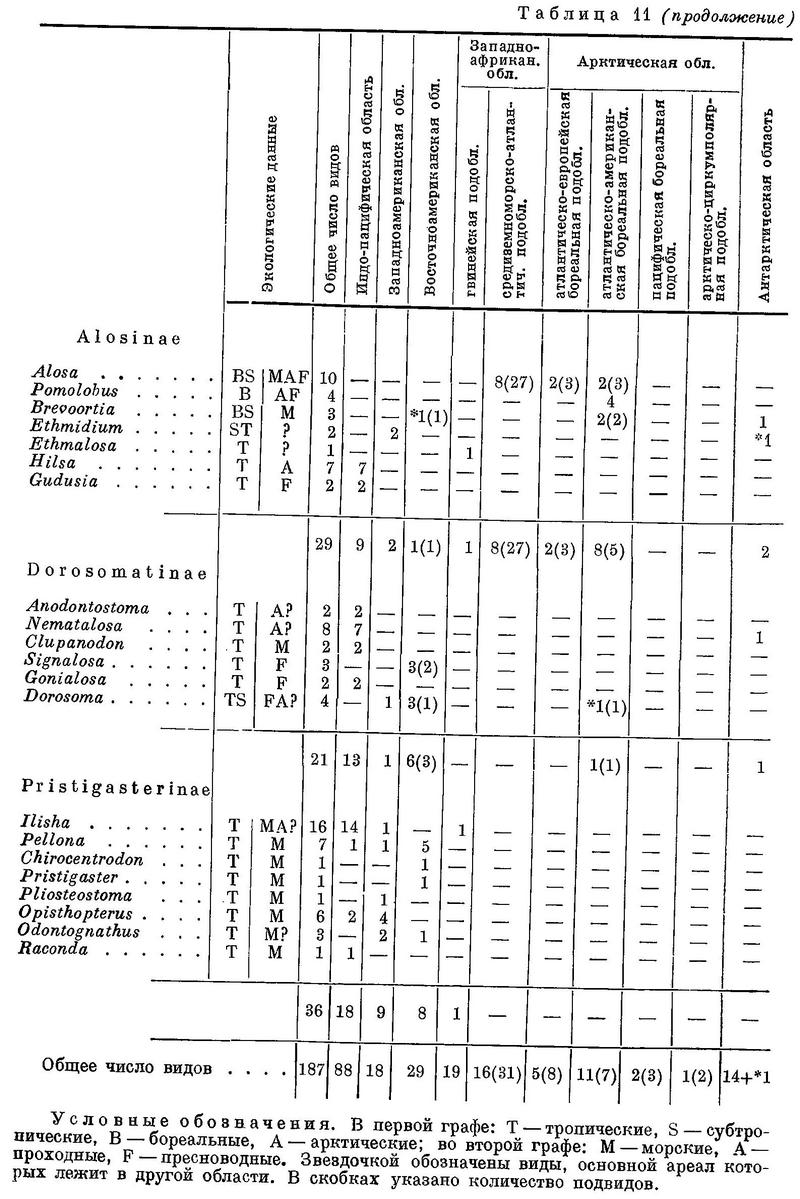
В остальных тропических областях сельдевых значительно меньше. Из этих областей в наибольшем количестве они представлены в восточноамериканской области, которой свойственно около 23 морских и проходных видов. Несколько родов (Rhinosardinia, Pristigaster, Chirocentrodon) эндемичны для этой области. Большая часть видов некоторых родов (Jenkinsia, Lile, Pellona), распространенных и в других тропических областях, встречается в восточноамериканской области, причем по количеству видов эти роды сравнительно малочисленны. Пресные воды бассейна восточноамериканской морской области относятся в основном к бразильской подобласти Л. С. Берга. Здесь встречаются 6 видов сельдевых, принадлежащих к 2 родам (Dorosoma, Signalosa), отдельные виды которых встречаются и далее к северу за пределы подобласти. Некоторые виды, упомянутые среди морских, заходят и, возможно, даже проводят всю жизнь в пресной воде (например Lile platana, Rhinosardinia amazonica). Общее число сельдевых, свойственных восточноамериканской морской области и бразильской пресноводной подобласти, достигает до 30 видов, относящихся к 15 родам, из которых 4 эндемичные (помимо перечисленных морских, Signalosa).
В еще меньшем количестве встречаются сельдевые в западноамериканской области, имеющей лишь 17 видов, относящихся к 11 родам. Единственным эндемичным для этой области, не встречающимся за пределами ее, является монотипический род Pliosteostoma. Из родов, общих западноамериканской и другим тропическим областям, нет ни одного, который был бы представлен в этой области преобладающим количеством видов. В пресных водах, относящихся к бассейну западноамериканской области, из пресноводных сельдевых известен лишь один, описанный недавно вид (Dorosoma smithi). До этого род Dorosoma являлся эндемичным для восточноамериканской области.
В западноафриканской области насчитывается до 18 видов морских и проходных сельдевых, но тропическими из них будут лишь 5 видов, встречающиеся в гвинейской подобласти. Эндемичный в этой подобласти лишь один монотипический род Ethmalosa, образ жизни и условия размножения которого точно не известны. Остальные 4 встречающихся в этой подобласти морских вида относятся к 2 родам (Sardinella и Ilisha) с наибольшим среди сельдевых количеством видов, преобладающее количество которых свойственно индо-пацифинеской области. Значительно больше сельдевых в пресных водах гвинейской подобласти, которые относятся к среднеафриканской подобласти принятых нами зоогеографических подразделений для пресноводных рыб. Этой подобласти свойственно 14 видов, принадлежащих 8 эндемичным родам (Pellonula, Poecilithrissa, Microthrissa, Potamothrissa, Cynothrissa, Odaxothrissa, Stolothrissa, Limnothrissa), обитающих главным образом в р. Конго и ее бассейне и оз. Танганайке. Упомянутый эндемичный для гвинейской подобласти род Ethmalosa, известный из устья р. Конго, возможно, есть также пресноводный или проходной. Следовательно, общее количество (сельдевых гвинейской подобласти и относящейся к ее бассейну среднеафриканской подобласти распространения пресноводных рыб исчисляется 19 видами, из которых 15 относятся к 9 эндемичным родам.
В средиземноморско-атлантической подобласти встречаются 16 видов морских, солоноватоводных и проходных сельдевых западноафриканской вблисти, причем тропических из них лишь 2 (Sardinella eba и Sardinella aurita); последний широко распространен почти во всех тропических областях, заходит и в субтропики. Остальные сельдевые этой подобласти относятся к субтропическим родам (Sardina) или являются субтропическими формами (Sprattus sprattus phalericus, средиземноморские формы Alosa) видов в основном бореальных родов, большая же часть видов (все виды рода Clupeonella и каспийско-черноморские виды рода Alosa) являются бореальными. Эндемичны для средиземноморско-атлантической подобласти лишь два последних рода. Морских или солоноватоводных среди сельдевых этой подобласти 11 видов, проходных или полупроходных 4. Из числа последних видов Alosa caspia является в равной мере и морским, солоноватоводным. Фауна пресноводных рыб бассейна средиземноморско-атлантической подобласти относится к средиземноморской, западнобалканской и понто-каспийской провинциям средиземноморской подобласти Берга (Berg, 1933 : 182—208). Чисто пресноводных сельдевых здесь имеется всего лишь один вид (Clupeonella abrau) и 3 подвида рода Alosa (Alosa fallax lacustris, Alosa fallax gracilis, Alosa alosa macedonica) в озерах средиземноморского бассейна. Таким образом, всего в рассматриваемой подобласти вместе с пресноводными встречается 16 видов с 31 подвидом сельдевых, относящихся к 6 родам, в преобладающей массе морских или солоноватоводных.
Обширной арктической области Ортманна, включающей бореальные, арктические и отчасти субтропические элементы морской фауны, свойственно всего лишь около 15 видов морских (4) или проходных (10) сельдевых. Эндемичными для этой области являются роды Clupea и отчасти Pomolobus, причем первый встречается во всех ее подобластях. Чисто пресноводных родов и даже видов в пресных водах бассейна арктической области нет. Пресноводные здесь лишь некоторые подвиды проходных видов.
В атлантическо-европейской бореальной подобласти встречаются 5 видов сельдевых, принадлежащих 4 родам; из этих видов 3 морские, 2 проходные. Эндемичных родов, встречающихся в пределах только этой подобласти, нет, но из 4 встречающихся здесь родов 2 (Sprattus и Sardina) в западной половине северной части Атлантического океана, у берегов Сев. Америки, отсутствуют. В пресных водах (в основном балтийская провинция средиземноморской подобласти и отчасти ледовитоморская циркумполярной подобласти Л. С. Берга), относящихся к бассейну атлантическо-европейской бореальной подобласти, имеется всего лишь один подвид (Alosa fallax killarnensis, оз. Килларней в Ирландии).
Наибольшее количество видов (11) сельдевых, относящихся к 6 родам, имеется в атлантическо-американской бореальной подобласти, причем большая часть их, 4 вида, относятся к эндемичному для этой подобласти роду Pomolobus, лишь один из видов которого (Pomolobus chrysochloris) встречается в прилегающих частях восточноамериканской области. Из числа встречающихся здесь видов чисто морской лишь один (Clupea harengus), все остальные проходные или полупроходные, или солоноватоводные. Соответственно несколько больше здесь по сравнению с пресными водами, относящимися к бассейну предыдущей подобласти, и пресноводных рыб, обитающих в миссисипской подобласти Л. С. Берга (Alosa alabamae ohiensis и формы, которые образует в пресных водах Dorosoma cepedianum). Кроме того, жилые формы в пресных водах имеют некоторые виды Pomolobus.
В пацифической бореальной подобласти сельдевые представлены лишь 2 подвидами, основные формы которых встречаются в других подобластях и областях. Основная форма одного из этих видов (Clupea harengus) свойственна атлантическо-европейской и атлантическо-американской бореальным подобластям, второго (Sardinops sagax) — антарктической области. Для пацифической бореальной подобласти характерно полное отсутствие проходных сельдевых как у американских, так и у азиатских берегов. В пресных водах, относящихся к бассейну этой подобласти (амурская переходная область, колорадская и отчасти циркумполярная подобласти Л. С. Берга), нет также и чисто пресноводных сельдевых.
Наиболее бедна сельдевыми арктическо-циркумполярная подобласть рассматриваемой области, где имеются всего лишь 2 подвида одного и того же вида (Clupea harengus), основная форма которого широко распространена в атлантическо-европейской и атлантическо-американской, а близкий подвид — в тихоокеанской бореальных подобластях. Проходных сельдевых в этой подобласти нет; нет и пресноводных в относящейся к бассейну ее циркумполярной подобласти Л. С. Берга.
Фауна сельдевых обширной антарктической области имеет в основном субтропический характер, в меньшей степени — тропический или бореальный. Антарктических среди сельдевых нет. Всего в этой области насчитывается около 14 видов, в основном морских, отчасти проходных (2 вида), относящихся к 9 родам, причем большая часть видов (5) относится к роду Sprattus и некоторые из них, судя по всему, являются лишь подвидами. Преобладающее число встречающихся в этой области видов (около 12) эндемичные. Эндемичных из числа морских и проходных родов здесь нет. В наибольшем количестве в антарктической области сельдевые имеются у южных берегов Австралии и Новой Зеландии (8 видов), южной оконечности Южн. Америки (5 видов), немногие (2 вида) у южных берегов Африки. Один из этих видов (Sardinops sagax) в качестве подвидов представлен во всех перечисленных местах. Пресноводных видов идесь лишь один, относящийся к эндемичному роду Potamalosa (реки Нового Южн. Уэльса и Виктории).
Тропические морские и проходные сельдевые наиболее многочисленны и эндемичны, как мы видели, в индо-пацифической области, которая наиболее богата представителями и других групп прибрежных водных животных (Ekman, 1934 : 321).
Из других тропических областей по числу видов, родов и эндемизму наиболее богата восточноамериканская область. Эта область, по Экману (Ekman, 1935 : 8), наиболее близка к индо-пацифической благодаря связям, которые имеются между обеими областями в ряде групп животных. Из 12 встречающихся в восточноамериканской области морских и проходных родов семейства общими с индо-пацифическими являются 4 рода: Sardinella (13 видов индо-пацифических, один из них встречается и в восточноамериканской области), который есть в гвинейской подобласти, но отсутствует в западноамериканской области, Harengula (13 видов индо-пацифических, 4 восточноамериканских), Etrumeus (2 вида индо-пацифических, один восточноамериканский), Pellona (один вид индо-пацифический, 5 восточноамериканских); последние 3 рода, наоборот, отсутствуют в гвинейской подобласти и есть в западноамериканской.
Такую же связь с индо-пацифической областью имеет и менее богатая сельдевыми западноамериканская область. Из 12 встречаемых здесь родов общими с индо-пацифической областью являются: Etrumeus (один вид, встречаемый и в индо-пацифической области), Harengula (2 вида), Opisthopterus (4 вида западноамериканских, 2 индо-пацифических), Pellona (один вид) и Ilisha (один вид западноамериканский, 14 индо-пацифических), который есть в гвинейской подобласти, но отсутствует в восточноамериканской области.
Наиболее бедная как сельдевыми, так и другими водными прибрежными животными гвинейская подобласть имеет много общего с сельдевыми индо-пацифической области. Из 3 встречающихся здесь морских родов сельдевых лишь один, эндемичный для этой области род Ethmalosa, не встречается в индо-пацифической области. Остальные роды (Sardinella — 3 вида и Ilisha — один вид) в большей мере представлены в индо-пацифической области. Первый род связывает гвинейских сельдевых также и с восточноамериканскими, последний (Ilisha) — с западноамериканскими.
Таким образом, пелагическая и абиссальная свободная от островов восточная часть Тихого океана между Полинезией и Америкой представляет преграду (ostpazifische Sperre; Ekman, 1935 : 106) к распространению из индо-пацифической области в западноамериканскую не только тропической литоральной фауны, но и сельдевых, которые хотя и являются пелагическими, но, как указывалось выше, теснейшим образом связаны с берегами. Однако благодаря пелагическому и более подвижному образу жизни сельдевых, для некоторых из них эта преграда оказалась менее непреодолимой, чем для ряда типично литоральных животных, так как и в индо-пацифической области, и в западноамериканской имеются не только виды, относящиеся к одним и тем же родам, но и виды идентичные (Etrumeus micropus).
Почти такую же преграду для распространения тропической литоральной фауны представляет и центральная часть Атлантического океана (zentralatlantische Sperre; Ekman, 1935 : 107), причем некоторые сельдевые расселяются и через эту преграду, так как по обе стороны тропической части Атлантического океана, помимо видов одних и тех же родов, встречается и общий вид (Sardinella aurita).
Значительно большую близость имеют сельдевые восточно- и западноамериканской областей. Общими для обеих областей являются следующие роды: с одной стороны, упомянутые выше, встречающиеся и в индо-пацифической области Pellona (5 видов в восточноамериканской области, один в западноамериканской), Harengula (4 вида в восточноамериканской области, 2 в западноамериканской), Etrumeus (по одному виду в восточно- и западноамериканской областях), с другой, — роды, свойственные только восточно- и западноамериканской областям — Odontognathus (один вид в первой, 2 во второй), Lile (2 вида в восточноамериканской области, один в западноамериканской), Opisthonema (по одному виду в каждой области) и Jenhinsia (3 вида в восточноамериканской области и один в западноамериканской) и в основном пресноводный род Dorosoma (3 вида в восточноамериканской области, один в западноамериканской). Следовательно, связь между сельдевыми восточно- и западноамериканской областями несколько большая, чем между этими областями и индо-пацифической областью. Помимо 3 эндемичных родов в восточноамериканской области и одного в западноамериканской здесь имеются 5 упомянутых родов, встречающихся только в этих областях. Характерно отсутствие общих видов, встречающихся и в восточно- и в западноамериканской областях, обусловленное отсутствием возможностей обмена фаун обеих областей с миоцена или по крайней мере с плиоцена (Берг, 1918 : 1838) вследствие поднятия Панамского перешейка. Наличие эндемичных родов в каждой из рассматриваемых областей и значительного числа родов, встречающихся только в этих областях, свидетельствует о том, что восточно- и западноамериканскую области следует рассматривать как географический центр сельдевых, правда, значительно меньший, чем индо-пацифическая область. Значительно большее в восточноамериканской области, чем в западноамериканской, общее количество видов, эндемичных родов и видов, принадлежащих общим только для обеих областей родам, следует рассматривать как свидетельство того, что географический центр этих сельдевых лежит в восточноамериканской области.
Бедность прибрежной тропической фауны западного побережья Африки (гвинейской подобласти), помимо влияния причин исторических, объясняют (Ekman, 1935 : 88) более низкими температурами воды, строением дна и пр. Для сельдевых, в частности, наибольшее значение помимо узости тропической области у западных берегов Африки, имеет, как нам кажется, слабое развитие береговой линии вследствие малой изрезанности берегов и отсутствия островов, что для сельдевых, хотя пелагических, но в основном прибрежных рыб, имеет существенное значение. Совершенно очевидно, что, помимо общеизвестных исторических причин (Ekman, 1935 : 97), богатство индо-пацифической области как сельдевыми, так и вообще литоральными животными в значительной степени обусловлено наиболее сильным развитием здесь береговой линии и большим количеством островов. Подобным же образом следует объяснять и меньшее богатство западноамериканских морских сельдевых по сравнению с восточноамериканскими, так как помимо большей узости западноамериканской области по сравнению с восточноамериканской береговая линия западных берегов Америки изрезана слабо и почти не имеется островов.
Пресноводные сельдевые наиболее многочисленны и эндемичны, как указывалось выше, в тропических областях, где известны пресноводные виды даже таких типично морских рыб, как акулы и скаты. В тропических областях условия для перехода от морского к пресноводному образу жизни наиболее благоприятны; в объяснение причин этого высказан ряд мнений [сводку этого вопроса см. у Пирса (Pearse, 1936)]. В частности, Пельзенер (Pelseneer, 1906 : 699) более активный переход морских животных в пресные воды в Индо-малайском архипелаге объясняет пониженными соленостью и плотностью воды окружающих морей. Однако наиболее многочисленны и эндемичны пресноводные сельдевые, как указывалось выше, в пресных, водах западной Африки, у берегов которой таких опреснений нет. Большое количество пресноводных сельдевых в среднеафриканской подобласти, нам кажется, обусловлено теми же причинами, что и бедность их в морских водах гвинейской подобласти — слабой изрезанностью береговой линии и почти полным отсутствием здесь островов и, кроме того, наличием обширных пресноводных бассейнов (р. Конго, оз. Танганайка). Не найдя здесь достаточно благоприятных условий в море, сельдевые перешли в пресные воды, где дали пышный расцвет, образовав большое количество эндемичных родов. Что касается пресноводных сельдевых сино-индийской области, то значительное количество их, как и исключительное богатство морских индо-пацифической области, обусловлено в первую очередь причинами историческими. Известное значение, конечно, имело и поверхностное опреснение морей Индо-малайского архипелага (Pelseneer, 1906), но несомненно большее — обширные речные системы, в которые даже при наличии исключительно благоприятных условий для прибрежных животных в море (развитие береговой линии, архипелаг островов с массой проливов и заливов) и перешел ряд сельдевых. Последнее обстоятельство, нам кажется, наиболее удовлетворительно объясняет и наличие сельдевых в пресных водах бассейна, восточноамериканской области, в которую впадает, ряд крупных речных систем, и отсутствие их в пресных водах бассейна западноамериканской области, не имеющей больших рек.
Во внетропических областях северного полушария количество видов семейства и их эндемизм последовательно уменьшаются в средиземноморско-атлантической, атлантическо-американской бореальной, атлантическо-европейской бореальной, пацифической бореальной и арктическо-циркумполярной подобластях.
Средиземноморско-атлантическая и атлантическо-европейская бореальные подобласти характерны некоторыми общими, эндемичными для обеих них родом (Sardina) и видами (Sprattus sprattus, Alosa alosa, Alosa fallax); первая подобласть отличается от последней отсутствием рода Clupea и наличием рода Clupeonella и одного из видов Sardinella (Sardinella aurita).
Общими для атлантическо-европейской и атлантическо-американской бореальных подобластей являются роды Alosa и Clupea, встречающиеся по обе стороны северной части Атлантического океана; последний есть в северной части Тихого океана и отчасти в арктических морях, являясь общим для всех подобластей арктической области (см. табл. 11). Некоторые роды встречаются лишь по одну сторону северной части Атлантического океана: роды Sprattus и Sardina — у берегов Европы в атлантическо-европейской бореальной и средиземноморско-атлантической подобластях, роды Pomolobus и Brevoortia — у берегов Сев. Америки, причем роды Sprattus и Brevoortia имеются и в южном полушарии (Brevoortia у берегов Южн. Америки, Sprattus, кроме того, у берегов Австралии и Новой Зеландии).
Для атлантическо-европейской бореальной подобласти характерно преобладание морских сельдевых (3 вида) над проходными (2 вида), для атлантическо-американской бореальной подобласти наоборот — преобладание проходных (8 видов) над морскими и солоноватоводными (3 вида). Наряду с преобладанием проходных сельдевых в последней подобласти естественно значительно больше и пресноводных форм (4), образуемых проходными, чем в первой подобласти (1).
Сельдевые пацифической бореальной подобласти, как указывалось выше, связаны, с одной стороны, с сельдевыми обеих атлантических бореальных подобластей и арктическо-циркумполярной (Clupea harengus), с другой — с антарктической областью (Sardinops sagax). Сельдевые арктическо-циркумполярной подобласти теснейшим образом связаны с обеими атлантическими бореальнымй подобластями и в еще большей степени с пацифической. О причинах отсутствия в пацифической бореальной и арктическо-циркумполярной подобластях проходных и пресноводных сельдевых сказано ниже.
Сельдевые антарктической области имеют связи с сельдевыми почти всех областей и подобластей, за исключением гвинейской и арктическо-циркумполярной подобластей; наиболее теплолюбивые из них, имеющие в основном субтропический характер, связаны с индо-пацифической областью (Spratelloides — 4 вида индо-пацифические, 2 антарктические; Hyperolophus — один вид пацифический, один антарктический, Nematalosa — 7 видов индо-пацифических, один антарктический) и отчасти с другими тропическими областями (Etrumeus — 2 вида индо-пацифических, встречающихся, кроме того, в западноамериканской и антарктической областях, один восточноамериканский, один антарктический). Ряд родов является общим с умеренными и субтропическими подобластями северного полушария (Sprattus — один вид в средиземноморско-атлантической и атлантическо-европейской бореальной подобластях, 5 в антарктической области у берегов Южн. Америки, Австралии и Новой Зеландии; Sardinops — один вид в пацифической бореальной подобласти у берегов Сев. Америки и юго-западной Азии и в антарктической области у берегов Австралии, Южн. Америки и южной Африки; Brevoortia — 2 вида в атлантическо-американской бореальной подобласти, один из которых представлен подвидом в восточноамериканской области и один в антарктической области). Наибольшие связи, следовательно, наблюдаются с индо-пацифической областью, в меньшей степени — с умеренными подобластями северного полушария и с восточно- и западноамериканской областями.
Таким образом, наибольшие связи естественно обнаруживают сельдевые морские и проходные; распространение пресноводных как правило ограничено какой-либо одной областью или подобластью.
Большинство морских рыб (например сем. Gadidae, Pleuronectidae, Scombridae и др.), как известно, имеют пелагическую икру. Однако у ряда морских рыб (например у сем. Blenniidae, Gobiidae и др.) икра приклеивающаяся. По этой особенности икры, как и по отношению к солености, сельдевые сильно разнятся между собой. У преобладающего большинства сельдевых икра пелагическая, у многих — демерсальная, некоторые имеют приклеивающуюся икру. При этом приклеивающаяся икра свойственна как морским (Clupea), так и проходным и пресноводным (Pomolobus). Точно также и пелагическую икру имеют как почти все морские сельдевые, так и проходные и пресноводные. Следует отметить, что икра проходных сельдевых, например Alosa kessleri volgensis, может развиваться при солености до 25—30‰ (Олафан, 1939 : 470), а наилучшие условия для развития икры Alosa sapidissima, по экспериментальным исследованиям (Leim, 1924 : 459), — солоноватая вода с содержанием солей около 7.5‰. Икра некоторых сельдевых (например Clupeonella delicatula, Alosa caspia) может развиваться как в пресной, так и в солоноватой воде. Ряд сельдевых, наоборот, развивается при повышенном содержании солей (например Sardinella aurita). Следовательно, судя по этой экологической особенности, сельдевых по происхождению следует считать морскими или солоноватоводными рыбами. Приклеивающаяся икра Clupea harengus ни в коей мере не свидетельствует в пользу пресноводного происхождения сельдевых и более поздней миграции этой сельди из пресных вод в море (Есипов, 1939 : 164). Если у сельдевых, как полагает В. К. Есипов, при переходе от пресноводного образа жизни к морскому икра из приклеивающейся становится пелагической, то почему же она пелагическая или демерсальная у пресноводных сельдевых? Проходные или пресноводные сельдевые, повидимому, произошли от солоноватоводных. Морское или солоноватоводное происхождение сельдевых подтверждается и нахождением их ископаемых остатков (в пресноводных отложениях сельдевых не найдено), все они известны из морских отложений.
Таким образом, сельдевых по происхождению следует считать морскими и солоноватоводными, а по отношению к температуре — тропическими и в меньшей степени умеренно тепловодными. Богатство как сельдевых, так, по взглядам Экмана (1934 : 361; 1935 : 110), и других тропических прибрежных животных в индо-пацифической области не свидетельствует о том, что эту область следует рассматривать в качестве центра их происхождения. Прибрежная индо-пацифическая фауна, как и фауна других тропических областей, есть наследие фауны мезозойского моря Тетис, которое простиралось через среднюю часть современного Атлантического океана и Средиземное море к Индокитаю и Индо-малайскому архипелагу и объединяло восточную часть Тихого океана, среднюю часть Атлантического, Средиземное море, Индийский океан и западную часть Тихого в единое море. Фауна Тетиса была также единой и одинаково богатой во всех частях его. В миоцене Тетис значительно сократился, потерял связь с Индийским океаном и западной частью Тихого; приблизительно одновременно или несколько позднее в связи с поднятием Панамского перешейка утратилось соединение с восточной частью Тихого океана. Вследствие одновременного похолодания климата меловая и нижнетретичная фауна Тетиса в разобщенных частях его сохранилась не везде в одинаковой мере. Все это и обусловило неодинаковое богатство и различные степени связи современных сельдевых и других прибрежных животных разных тропических областей.
В ископаемом состоянии сельдевые и близкие к ним семейства Elopidae и Мegalopidae известны с нижнего мела (Берг, 1940 : 222, 223), сем. Leptolepidae — от верхнего триаса до середины мела (Берг, 1940 : 221; Woodward, 1895 : 500—530; Rayner, 1937 : 46—74). Из области Тетиса сельдевые известны как из неогеновых, так и из палеогеновых отложений (Woodward, 1901 : 146—159; Arambourg, 1925 : 10—15; 1927 : 18—32, и др.). Фауна рыб Тетиса в палеоцене, эоцене и олигоцене была тропической (Leriche, 1902, 1905, 1906, 1910), но с похолоданием климата, который к концу миоцена стал почти современным, тропическая фауна Тетиса исчезла. Уже в среднем миоцене климат в области Чокракского бассейна был близок к субтропическому — приблизительно такой, как современный климат Мраморного моря (Жижченко, 1940 : 153). Ископаемые сельдевые из области Тихого океана известны значительно позднее, из калифорнийских отложений в миоцене (Jordan a. Gilbert, 1919 : 25—34; Jordan, 1921 : 241—248; 1925 : 8—12; David, 1943 : 3—9, 54—55, 89—95, 120—122). В южном полушарии в ископаемом состоянии сельдевые известны из верхнеэоценовых и четвертичных отложений Бразилии (Schaeffer, 1947 : 17—24).
Бореальные и субтропические сельдевые, как указывалось выше, наиболее бедны в пацифической бореальной подобласти, между тем как в соответствующих ей обеих атлантических подобластях их значительно больше. В то же время известно, что фауна как рыб, так и других водных животных северных частей Тихого океана, имея большое сходство с фауной соответствующих частей Атлантического океана, отличается значительно большим числом и видов, и родов, и даже семейств (Günther, 1880 : 178; Шмидт, 1904 : 394; Берг, 1918 : 1835; Broch, 1936 : 42, и др.). По общепринятым в настоящее время взглядам, современная фауна северных частей Атлантического океана, а также отчасти и арктических морей в основной своей массе есть обедненная фауна Тихого океана и ведет начало от последней. Среди рыб исключение в этом отношении представляют тресковые, которые, наоборот, наиболее эндемичны и наибольшим количеством видов и родов представлены в северной части Атлантического океана. Этот общеизвестный и отмеченный уже давно (Günther, 1880) факт был подробно рассмотрен автором (Световидов, 1940 : 50; 1948 : 41). Подобным же исключением, как указывалось выше и как уже ранее отметил автор (Световидов, 1944 : 152), оказываются и сельдевые.
В северной части Атлантического океана, в атлантическо-европейской и атлантическо-американской бореальных подобластях встречаются 14 видов сельдевых (см. табл. 11), относящихся к родам Clupea, Alosa, Pomolobus, Sprattus, Brevoortia, Sardina, Etrumeus и Dorosoma, виды трех первых родов — бореальные, Sprattus и Brevoortia — несколько более теплолюбивы, умеренно-субтропические, Sardina — субтропический, виды трех последних родов наиболее теплолюбивы. Если принять во внимание 12 видов (роды Alosa, Clupeonella и Sardinella), встречающихся, кроме того, в средиземноморско-атлантической подобласти, то во внетропических областях северной части Атлантического океана всего насчитывается до 26 видов. Из числа последних 12 видов 2 (Sardinella aurita, Sardinella eba) в основном тропические, встречающиеся и в субтропических морях; остальные — бореальные.
В северной части Тихого океана, в пацифической бореальной подобласти, встречается всего лишь 2 вида, из которых один (Clupea harengus) свойствен умеренным широтам, второй (Sardinops sagax) является субтропическим. Кроме того, сюда заходят более теплолюбивые субтропические и тропические (Etrumeus micropus, Clupanodon punctatus, Harengula zunasi, Ilisha elongata).
Северной части Атлантического океана, как видим, свойственно значительно большее количество родов и преобладающее количество видов и подвидов (см. табл. 11) сельдевых по сравнению с северной частью Тихого океана. Кроме того, что самое существенное, в северной части Атлантического океана сосредоточены все бореальные роды сельдевых северного полушария. Не менее важно также то, что все бореальные проходные роды и виды семейства (все виды Pomolobus и Alosa), многие из которых образуют пресноводные формы, свойственны также северной части Атлантического океана и прилегающим морям.
Численное преобладание в северной части Атлантического океана бореальных и субтропических сельдевых, наличие здесь ряда эндемичных и среди них в особенности проходных родов и видов, отсутствие самобытности фауны сельдевых северной части Тихого океана и явная зависимость единственного бореального представителя ее (Clupea harengus pallasi) от североатлантических — все это с несомненностью указывает на его атлантическое происхождение. Так как в умеренных частях современного Атлантического океана в нижнетретичное время климат и фауна были тропическими (Leriche, 1902, 1905, 1910), то бореальных сельдевых, как и других рыб, в частности тресковых (Световидов, 1940 : 54; 1948 : 43), в них не было. Очевидно, бореальные сельдевые, подобно тресковым, в нижнетретичное время держались в более холодноводном бассейне — в полярном и прилегающем к нему скандском, впадины которых, как показали исследования последних лет (Панов, 1946 : 29), к началу третичного времени уже имели вполне морской характер и достаточные глубины.
В расселении и формировании современного распространения сельдевых те изменения, которые претерпело в верхнетретичное время море Тетис, а позднее, северная часть Атлантического океана и в области Берингова пролива северная часть Тихого океана, и сопутствующее им похолодание климата играли такую же существенную роль, как и для тресковых (Световидов, 1940 : 57; 1948 : 44). При верхнетретичном похолодании климата бореальные представители сельдевых вместе с тресковым могли спуститься из полярного бассейна в северную часть Атлантического океана, а не Тихого, так как в области Берингова пролива в это время, как известно, простиралась суша. В северную часть Тихого океана бореальные сельдевые могли попасть не ранее плиоцена и затем в послеледниковое время, когда в области Берингова пролива происходили трансгрессии, а климат был более теплым (Берг, 1918 : 1839; 1934 : 73; Smith, 1919 : 167).
Таким образом, неоднократно высказываемый взгляд (Дерюгин, 1928 : 444; Берг, 1934 : 77), согласно которому Clupea harengus проникла в Белое море из Тихого океана, в силу изложенных выше фактов должен быть изменен на обратный.
Современное распространение Clupea harengus может служить, как и у тресковых (Световидов, 1940 : 58; 1948 : 47), указанием на пути проникновения его в Тихий океан. Наиболее северное распространение в Атлантическом океане Clupea harengus имеет не у берегов Сев. Америки, как одна из самых холодолюбивых форм Gadus morhua (Gadus morhua ogac — западные берега Гренландии до 72.5—73.5° с. ш., Гудсонов прол. и зал. Джемса), а у берегов северной Азии (Clupea harengus pallasi n. suworowi — от Чёшской губы на восток до Обской губы, Clupea harengus pallasi — на запад до бухты Тикси близ дельты р. Лены). Это различие в распространении совокупно с большей евригалинностью Clupea harengus по сравнению с Gadus morhua является характерным примером различных путей миграции еври- и стеногалинной фауны из одного океана в другой.
Как известно, многочисленные и многоводные реки Сибири оказывают значительно более опресняющее влияние на моря, омывающие ее берега, чем немногочисленные и сравнительно маловодные реки Сев. Америки на моря ее северного побережья. Благодаря этому все наиболее стеногалинные элементы морской фауны мигрировали из одного океана в другой вокруг Сев. Америки. К числу их относятся, например, иглокожие, наиболее стеногалинные из морских животных. Миграции иглокожих из Тихого океана в Атлантический происходили вокруг Сев. Америки (Дьяконов, 1927 : 275; 1936 : 752; 1945 : 141). Этим же путем, но в обратном направлении — из Атлантического океана в Тихий — прошла и Gadus morhua, о чем свидетельствует современное распространение наиболее холодолюбивых широкоголовых подвидов этого вида и морфологическая и экологическая близость тихоокеанской трески (Gadus morhua macrocephalus) и гренландской (Gadus morhua ogac) (Световидов, 1940 : 58; 1944 : 154; 1948 : 48). Наоборот, Clupea harengus, и в особенности малопозвонковый подвид его (Clupea harengus pallasi и его формы, см. сравнительные замечания этого вида в «Систематической части» и Световидов, 19496 : 59), более евригалинный, чем Gadus morhua, совместно с другими евригалинными морскими организмами проник из Атлантического океана в Тихий вокруг северной Азии, о чем свидетельствует распространение малопозвонковых подвидов его и морфологическая и экологическая близость Clupea harengus pallasi и Clupea harengus pallasi n. suworowi. Этим же путем из тресковых в Тихий океан из Атлантического прошла, в частности, навага (Eleginus navaga — на восток до Обской губы, Eleginus gracilis — на запад до бухты Питлекай), рыба значительно более евригалинная, чем треска (Световидов, 1940 : 58, 1948 : 48, фиг. 33). Clupea harengus и Gadus morhua, как показал автор (1944 ; 146), имеют большое сходство в систематическом отношении и экологических особенностях отдельных подвидов, обладая в то же время рядом диаметрально противоположных эколого-биологических отличий. Эти черты сходства и различия обоих видов и их подвидов, несомненно, обусловлены их совместным существованием на протяжении по крайней мере верхнетретичного времени.
В настоящее время затруднительно сказать, когда Clupea harengus проник в Тихий океан — в плиоценовую или, как предполагал Дерюгин (1928 : 444), позднее, в послеледниковую трансгрессию. Все соображения, высказанные автором (Световидов, 1940 : 57; 1948 : 47) на этот счет относительно Gadus morhua целиком приложимы и к Clupea harengus, т. е. наиболее вероятно, что и этот вид прошел сюда в послеледниковую трансгрессию. В связи с этим следует остановиться на рассмотрении вопроса о возможности проникновения из одного океана в другой этим же путем субтропических элементов фауны.
По мнению Берга (1934 : 76), в верхнеплиоценовом Сибирском море должны были встречаться, а следовательно, и иметь возможность проникнуть этим путем из одного океана в другой некоторые и субтропические, называемые им средиземноморско-бореальные виды животных, встречаемые в настоящее время лишь у берегов Калифорнии и Японии и в форме тождественных или близких видов в Средиземном море. К числу таковых он относит угрей Echelus ( = Myrus) uropterus (берега южной Японии) и Echelus myrus (Средиземное море и Бискайский зал.) и Cepola schlegeli (Токио и южнее) и Cepola taenia (Средиземное море и берега Англии до Йоркшира). Оба эти рода, заключающие по 2 упомянутых вида, в тропиках отсутствуют. Из моллюсков сходное распространение имеют средиземноморские Gibbula adriatica и Cymatium carrigatum, представленные близкими формами у берегов Калифорнии. Казалось, этим же путем могли бы проникнуть в Тихий океан и некоторые субтропические, сельдевые и тресковые, однако в действительности этого нет. Ряд фактов заставляет признать, что субтропические организмы не имели возможности мигрировать вокруг Сев. Америки и северной Азии из одного океана в другой. Более того, даже некоторые из бореальных тресковых, лишь несколько более теплолюбивые, чем Gadus morhua, пройти из Атлантического океана в Тихий этим путем не могли. Так, например, Melanogrammus aeglefinus и Pollachius virens и ряд других лишь слегка более теплолюбивых тресковых, чем Gadus morhua, и не заходящих так далеко на север, как эта рыба, в Тихий океан не проникли несомненно потому, что температурные условия в Сибирском море ни в послеледниковую, ни даже в плиоценовую трансгрессии для них не были подходящими. Это станет особенно ясным, если принять во внимание, что из подвидов Gadus morhua в Тихий океан проникла не узкоголовая (Gadus morhua morhua), а более холодолюбивая широкоголовая форма (Gadus morhua macrocephalus, близкая к Gadus morhua ogac). Равным образом из подвидов Clupea harengus в Тихий океан проникла не многопозвонковая (Clupea harengus harengus), а более холодолюбивая малопозвонковая форма (Clupea harengus pallasi, близкая к Clupea harengus pallasi n. suworowi). Температурные условия в области Берингова пролива во время обоих трансгрессий, повидимому, вообще не обладали широкой амплитудой. Верхний предел их в послеледниковую трансгрессию, повидимому, был таков, что смогли пройти лишь наиболее холодолюбивые подвиды Gadus morhua и Clupea harengus — широкоголовая (Gadus morhua macrocephalus) и малопозвонковая (Clupea harengus pallasi) формы, а более теплолюбивые — узкоголовая (Gadus morhua morhua) и многопозвонковая (Clupea harengus harengus) и некоторые другие, в частности уже упомянутые тресковые, пройти не смогли. Возможно, в плиоценовую трансгрессию температурные условия в области Берингова пролива были несколько выше, чем в послеледниковую, так как иначе в Тихий океан не смогли бы проникнуть некоторые из тресковых (например Theragra chalcogramma), но не настолько высоки, чтобы могли пройти субтропические организмы; в этом случае в Тихий океан имел бы возможность попасть ряд других, более теплолюбивых тресковых и сельдевых. Что касается нижнего температурного предела, то он, судя по всему, ни в одну из трансгрессий не был ниже современных. Это видно хотя бы из того, что Boreogadus saida проникла лишь в самую северную часть Берингова моря и вследствие отсутствия вдоль восточных берегов Камчатки южнее мыса Лопатки и у Алеутских о-вов льдов не может пройти далее — в Охотское море, где эта рыба нашла бы для себя подходящие условия. Очевидно, этот вид проник сюда лишь в самое последнее время, по окончании более теплого послеледникового времени. Таким образом, во время обоих трансгрессий было лишь сравнительно небольшое потепление, давшее возможность пройти, через Берингов пролив, из одного океана в другой лишь части умеренных элементов фауны обоих океанов. Об этом же свидетельствуют и многочисленные примеры ископаемых моллюсков, приводимые Бергом (1934 : 74). Этим и обусловлено отсутствие в северной части Тихого океана субтропических и даже более теплолюбивых из умеренных тресковых и сельдевых, близких к североатлантическим.
Субтропический представитель сельдевых северной части Тихого океана (Sardinops sagax), судя по всему, проник сюда из южного полушария. Близкие между собой субтропические роды Sardinops и Sardina наиболее близки в свою очередь к тропическому роду Sardinella и, повидимому, имеют общее происхождение. Род Sardina обособился в субтропических морях северного полушария, Sardinops, судя по его распространению (Sardinops sagax sagax — тихоокеанские берега Южн. Америки, Sardinops sagax neopilchardus — у южных берегов Австралии и вокруг Новой Зеландии, Sardinops sagax ocellata — берега южной Африки, Sardinops sagax melanosticta — берега юго-восточной Азии, Sardinops sagax caerulea — тихоокеанские берега Сев. Америки) — в субтропических морях южного полушария. Современное биполярное распространение Sardinops sagax сложилось в ледниковое время, когда вследствие похолодания климата имел место обмен субтропических элементов фауны обоих полушарий (Берг, 1920 : 273; 1947 : 128; Berg, 1933 : 449). Род Sardina в южное полушарие распространиться не мог, так как наиболее близкое место, куда бы он мог в первую очередь попасть (берега южной Африки), оказалось занятым совершенно сходным биологически родом Sardinops (Sardinops sagax ocellata). В свою очередь те же причины препятствовали распространению и Sardinops sagax ocellata в северную часть Атлантического океана к берегам Европы. Род Sardinops имел возможность беспрепятственно расселиться лишь в северную часть Тихого океана, куда он и проник как вдоль американских (Sardinops sagax caerulea), так и азиатских берегов (Sardinops sagax melanosticta), дав таким образом биполярное и амфипацифическое распространение (Андрияшев, 1939). Южное полушарие было, повидимому, центром происхождения не только рода Sardinops, но и рода Sprattus, большинство видов которого распространено также здесь. Биполярный род Sprattus имеет ту особенность, что он в отличие от преобладающего большинства других рыб, имеющих биполярное распространение, отсутствует у берегов южной Африки. Такое распространение, встречаясь в Атлантическом океане у берегов Европы, с одной стороны, и у берегов Южн. Америки, с другой, из биполярных рыб имеют немногие (например род Micromesistius) и обусловлено оно несоответствием температурных условий вод, омывающих южную Африку, температурным условиям нереста этих рыб: поверхностная температура воды у берегов южной Африки не бывает ниже 15° даже в холодное время года, икрометание же Sprattus sprattus в северной части Атлантического океана и в Средиземном море происходит в среднем при 8—10° (Световидов, 1949). Южному полушарию помимо двух упомянутых родов свойственны и другие эндемичные роды сельдевых (Ethmidium, Potamalosa, отчасти Hyperolophus), что дает основание считать здесь третий, менее богатый центр сельдевых.
Бедность морских и отсутствие проходных и пресноводных сельдевых в арктическо-циркумполярной подобласти обусловлена теплолюбивым характером семейства в целом. Так, например, более холодолюбивое семейство тресковых, представленное наибольшим количеством родов и видов и умеренных областях и отсутствующее в тропиках, в арктическо-циркумполярной подобласти имеет значительно большее количество видов, чем сельдевые. Среди тресковых имеются не только арктические, но и высоко-арктические, в том числе несколько эндемичных высоко-арктических родов (Boreogadus, Arctogadus). Из значительно более теплолюбивых сельдевых один из самых холодолюбивых видов (Clupea harengus) проник лишь в умеренные части арктическо-циркумполярной подобласти. Холодолюбивых проходных, могущих проникнуть так далеко на север, как Clupea harengus, среди сельдевых не оказалось. Это обусловило отсутствие проходных сельдевых и в северной части Тихого океана, потому что бореальные вокруг северной Азии и Сев. Америки пройти не могли, а для проходных субтропических здесь нет подходящих температурных условий. Отсутствие проходных сельдевых в северной части Тихого океана является одной из характерных ее особенностей (Световидов, 1949в).
При сопоставлении состава фауны сельдевых Каспийского, Черного и Азовского морей обращают на себя внимание несколько особенностей. Одна из этих особенностей заключается в том, что фауна рыб Черного моря, как известно, значительно более богатая и разнообразная по своему составу, чем Каспийского, более богата и разнообразна и в отношении сельдевых, но не видами и более низкими формами (подвидами), а родами. Здесь, помимо общих с каспийскими родом Clupeonella и подродом Caspialosa, имеются еще 3 рода из этого семейства — Sprattus, Sardina, Sardinella и подрод Alosa s. str., широко распространенных, кроме Черного моря, в Средиземном море и в Атлантическом океане, а некоторые из них, кроме того, в Индийском и Тихом океанах. Эта особенность черноморской фауны сельдевых, свойственная, как отмечалось выше, вообще фауне Черного моря, обусловлена, как известно, инвазией в Черное море средиземноморской фауны после прорыва Босфора.
Еще больший интерес представляет другая особенность черноморских и каспийских сельдевых, не отмеченная до сего времени и заключающаяся в том, что общие обоим морям род Clupeonella и подрод Caspialosa в Каспийском море как в видовом отношении, так и по количеству более низких форм (подвидов) представлены значительно более богато, чем в Черном и Азовском морях. Эта особенность была отмечена автором ранее в предварительном сообщении (Световидов, 1941 : 807, предст. 13 VI 1940). Эта статья должна была быть напечатана одновременно со статьей А. А. Михайловской (1941 : 559, предст. 24 VI 1940), которая была знакома с выводами автора и, говоря о богатстве каспийских сельдей, сделала ссылку на нее. Вследствие того, что статья автора была опубликована позднее: статьи А. А. Михайловской, эта ссылка в ее статье помещена не была.
Состав фауны сельдевых этих родов в Каспийском и Черном и Азовском морях таков (В скобках указано количество подвидов.):

1 Четвертый вид этого рода Clupeonella abrau встречается в оз. Абрау и Абулионд; о нем сказано особо ниже.
Род Clupeonella в Каспийском море представлен 2 эндемичными видами и лишь одним — формой того же вида, что и в Каспийском, — в Черном и Азовском морях. В Каспийском море из рода Alosa имеются 6 видов, из которых 3 свойственны только этому морю; в Черном же и Азовском морях из этого рода встречается только 3 вида, представленные здесь формами видов, общих с каспийскими. Таким образом, в Черном и Азовском морях нет ни одного вида, который не был бы представлен в Каспийском и притом значительно большим количеством форм, чем в Черном и Азовском морях, в то время как в Каспийском море имеются 2 вида из рода Clupeonella и 3 вида из рода Alosa, отсутствующие в Черном и Азовском морях.
Богатство сельдевых обоих этих родов в Каспийском море обусловлено преобладанием главным образом морских, непроходных видов и подвидов. Таковыми из числа видов являются Clupeonella grimmi и Clupeonella engrauliformis и Alosa saposhnikovi, Alosa sphaerocephala и Alosa curensis, в то время как проходная (Alosa kessleri s. l.) и полупроходная (Alosa caspia s. l.) сельди и тюлька (Clupeonella delicatula s. l.) представлены в обоих морях. Из числа морских, непроходных сельдей общими для обоих морей являются лишь Alosa brashnikovi s. l. Подвидами в Каспийском море особенно богат морской, непроходной вид Alosa brashnikovi s. l., представленный здесь 8 подвидами, против одного черноморско-азовского (Alosa brashnikovi maeotica). Полупроходные и проходные виды имеют в Каспийском море значительно меньшее количество подвидов. Так, например, Alosa caspia, одни подвиды которого являются морскими, непроходными (Alosa caspia salina), другие полупроходными или отчасти даже проходными (Alosa caspia caspia и др.), представлен в Каспийском море 4 подвидами и, повидимому, еще
2 более низкими формами (nationes), а в Черном и Азовском морях — 3 подвидами. Типично проходной вид Alosa kessleri s. l. имеет в Каспийском море 3 подвида, а в Черном и Азовском лишь один, распадающийся на 2 более низких формы (nationes).
Необходимо также отметить некоторые биологические особенности каспийских и черноморско-азовских форм Alosa brashnikovi s. l. и Alosa caspia s. l., а именно то, что каспийские формы этих видов являются более морскими рыбами, чем черноморско-азовские. Все каспийские формы Alosa brashnikovi s. l. избегают пресной воды и размножаются только в море, между тем как черноморско-азовская форма этого вида (Alosa brashnikovi maeotica) заходит в совершенно опресненные участки Таганрогского зал., и в нижнюю часть дельты Дона. Имеются указания и на нерест Alosa brashnikovi maeotica почти в пресной воде. Среди каспийских форм Alosa caspia s. l. есть такие, которые мечут икру в солоноватой воде в море (Alosa caspia salina) или в предустьевых пространствах (Alosa caspia caspia), в то время как все черноморско-азовские формы этого вида мечут икру только в пресной воде, заходя в реки, озера и лиманы. То же самое следует сказать и о Clupeonella delicatula s. l.: Clupeonella delicatula caspia мечет икру как в пресной воде, так почти и по всему Каспийскому морю, между тем как Clupeonella delicatula delicatula в Азовском море нерестует в самых опресненных частях его — в Таганрогском заливе, в дельте Дона, в кубанских опресненных лиманах и лишь отчасти в море.
Заслуживает внимания и еще одна особенность каспийских форм обоих рассматриваемых родов, отмеченная ранее Киселевичем (1918 : 21), — их значительно большие размеры, по сравнению с черноморско-азовскими. Clupeonella delicatula caspia достигает в длину до 14 см, средние размеры ее около 8—10 см, между тем как размеры Clupeonella delicatula delicatula в Азовском море не превышает 9 см, имея в среднем около 7 см. Каспийские формы Alosa caspia s. l. точно так же значительно крупнее черноморско-азовских. Alosa caspia caspia иногда достигает в длину до 28 см, средние размеры его около 18—21 см; таковы же размеры и остальных каспийских форм этого вида, в то время как черноморско-азовские формы не превышают в длину 20 см, средние размеры их около 15 см; каспийские формы Alosa brashnikovi s. l. достигают в длину почти до 50 см, средние размеры их около 30 см; наибольшие размеры черноморско-азовскои формы этого вида (Alosa brashnikovi maeotica) не превышают 31 см, средние размеры крупной расы этой формы — от 16 до 20 см, мелкой расы — около 12—13 см. Наиболее крупная из каспийских форм Alosa kessleri s. l., Alosa kessleri kessleri, достигает в длину до 52 см, в половозрелом состоянии во время хода в реку основная масса ее имеет длину от 36 до 44 см. Более мелкая форма этого вида, Alosa kessleri volgensis, достигает в длину до 40 см, средние размеры ее во время хода в реку около 28 см, в то время как наибольшие размеры черноморско-азовской формы (Alosa kessleri pontica) в Дону не превышают 30, а в Днепре 39 см.
Наиболее важная особенность сельдевых обоих рассматриваемых родов заключается, однако, в том, что каспийские формы их различаются значительно больше, чем черноморско-азовские формы тех же видов. Различия между каспийскими формами Alosa caspia s. l. значительны как в количестве жаберных тычинок, так в некоторых случаях и внешне (например Alosa caspia knipowitschi). Черноморско-азовские формы Alosa caspia s. l., не различающиеся между собой внешне, значительно меньше, чем каспийские формы этого вида, отличаются друг от друга и количеством жаберных тычинок. Разница в средних значениях количества жаберных тычинок у наиболее различающихся по этому признаку черноморско-азовских форм Alosa caspia s. l., Alosa caspia tanaica и Alosa caspia nordmanni, равна только 4, а у менее различающихся Alosa caspia tanaica и Alosa caspia palaeostomi — всего лишь 0.1, между тем как у наиболее различающихся в этом отношении каспийских форм этого вида Alosa caspia salina и Alosa caspia knipowitschi разница в средних значениях количеств жаберных тычинок равна 55.5. Даже у наименее отличающихся по количеству жаберных тычинок каспийских форм этого вида Alosa caspia salina и Alosa caspia persica разница в средних значениях равна 7.8. Такова же степень различия между каспийскими формами Alosa caspia s. l. с одной стороны, и черноморско-азовскими, с другой, и в остальных признаках. Черноморско-азовская проходная сельдь (Alosa kessleri pontica), как показали исследования последних лет, распадается на 2 формы, существенно различающиеся биологически, но морфологически ни количеством и строением жаберных тычинок, ни какими-либо другими особенностями, кроме размеров, не отличающиеся и являющиеся более низкими таксономически формами (nationes). Обе каспийские проходные сельди (Alosa kessleri kessleri и Alosa kessleri volgensis) существенно отличаются друг от друга количеством и строением жаберных тычинок, некоторыми другими особенностями и отчасти даже внешне. Еще более различаются между собой каспийские формы Alosa brashnikovi s. l., представленного в Черном и Азовском морях одной формой, которая, подобно Alosa kessleri pontica, повидимому, распадается на 2 более низкие, чем подвиды, формы (nationes), различающиеся лишь размерами и биологическими особенностями.
Менее различаются между собой в Черном море и виды Alosa brashnikovi s. l. и Alosa kessleri s. l. Наиболее близкая к Alosa brashnikovi s. l. по количеству жаберных тычинок форма Alosa kessleri s. l., а именно Alosa kessleri kessleri, имеет не менее 59 тычинок, в то время как у Alosa brashnikovi kisselevitshi, имеющей наибольшее количество жаберных тычинок из всех каспийских форм этого вида, количество их не превышает 49. Что касается черноморско-азовских форм Alosa brashnikovi s. l. и Alosa kessleri s. l., то у Alosa brashnikovi maeotica наибольшее количество жаберных тычинок 46, а наименьшее количество их у Alosa kessleri pontica 47.
Богатство каспийских сельдевых родов Clupeonella и Alosa как в видовом отношении, так и количеством форм общих видов, их гигантизм по сравнению с черноморско-азовскими формами, отмеченные выше биологические особенности и большая степень отличий обусловлены различиями в условиях развития и формообразования сельдевых как в Каспийском и Черном и Азовском морях, так и в тех морях, которые предшествовали им. Поэтому для рассмотрения условий развития и формообразования каспийских и черноморско-азовских сельдевых обоих этих родов и выяснения причин такого резкого различия в составе их, размерах, биологических особенностях и степени отличий необходимо обратиться к истории развития этих морей, так как история развития фауны, по вполне справедливому замечанию Колесникова (1940 : 6), не должна быть оторвана от истории развития бассейна, в котором она обитает.
Понто-каспийский морской бассейн, как известно, претерпел сложную геологическую историю. К концу миоцена, при сокращении Тетиса, одновременно с похолоданием климата, северо-восточная часть его отделилась, образовался сарматский и ряд других пришедших ему на смену бассейнов, фауна которых в дальнейшем развивалась в значительной степени изолированно, находясь, в отличие от фауны образовавшегося к этому времени Атлантического океана и стоявшего в связи с ним Средиземного моря, под сильным воздействием периодических опреснений и осолонений. В меньшей степени опреснения и бассейны с затрудненным сообщением с нормально соленым морским бассейном или даже замкнутые бассейны в северо-восточной части Тетиса были и раньше, в нижнем и среднем миоцене (Жижченко, 1940 : 42, 148).
Таким образом, условия развития сельдевых в Атлантическом океане, и замкнутых в сарматском море и других подобных ему бассейнах сущестненно различались. В Атлантическом океане и в связанном с ним, смежном с Черным, Азовским и Каспийским морями, Средиземном море условия существования и эволюции фауны были, повидимому, довольно однообразны — здесь не было значительных и неоднократных изменений солености, при ледниковом похолодании климата теплолюбивые элементы фауны могли отступить южнее. В связи с этим состав фауны Атлантического океана и Средиземного моря с конца неогена и начала четвертичного периода больших изменений не претерпевал.
Совершенно в иных условиях протекало развитие сельдевых в сарматском и других, сменивших его впоследствии бассейнах. При первом же еще небольшом опреснении, в сарматском море, часть стеногалинных, чисто морских сельдевых, наряду с частью чисто морской фауны, должны были погибнуть, что отчасти могло иметь место и раньше в нижнемиоценовом и чокракском бассейнах (Жижченко, 1940 : 42, 151). Оставшиеся сельдевые под влиянием дальнейшего последовательного осолонения и мэотическом бассейне и опреснения в понтическом озере-море и последовательных изменений солености в ряде других более мелких морей, сменивших разделившееся на части понтическое озеро-море, должны были частично вымереть, а оставшиеся, повидимому, наиболее евригалинные, претерпеть ряд изменений, неоднократно приспособляясь к изменявшейся солености и сопутствовавшим этим изменениям сменам фауны. Каждый раз при осолонении часть сельдевых должна была вымирать или отступать в опресненные участки, при опреснении же, повидимому, давали пышный расцвет. При этом необходимо отметить, что при таких изменениях, повидимому, внутри вида могли появляться вновь отдельные признаки, свойственные предшествовавшим формам как это имело место при осолонении и опреснении в области Балтийского моря у форм четырехрогого бычка Myoxocephalus quadricornis (Берг, 1916 : 1343) и ракообразного Limnocalanus grimaldii (Ekman, 1914 : 336), т. е. солоноватоводные формы, приспособившись к морской воде и фауне и претерпев соответствующие изменения, при повторном опреснении и сопутствовавшем ему изменении фауны приспособлялись к новым условиям, могли давать формы, отчасти сходные с формами предыдущих солоноватоводных бассейнов. На возможность подобных обратных изменений у черноморских моллюсков указывает Садовский (1934 : 61). Пример подобных изменений в размерах и форме раковин являет также моллюск Planorbis multiformis (Plate, 1920 : 217). Многочисленные случаи возврата (реверсии) прежних признаков приводит Огнев (1946 : 3), давший сводку по этому вопросу.
В ископаемом состоянии из опресненных бассейнов понто-каспийской области известна сравнительно бедная в родовом отношении (главным образом роды Clupeonella и Alosa) фауна сельдевых из нижне-, средне- и верхнемиоценовых отложений Апшеронского п-ова, б. Ставропольской губ., окрестностей Дзауджикау и Бинагадов, Кировского района (Богачев, 1913 : 219; 1933 : 30—37; 1938 : 42—52; 1942 : 89—92; Жижченко, 1940 : 19, 20, 34; Колесников, 1940 : 270, 448; Геккер и Мерклин, 1946 : 646, и др.). Из плиоценовых отложений Болгарии и понтических Румынии ископаемые Alosa nordmanni и Clupea delicatula и Clupea sulanae описаны в двух недоступных автору статьях (Pauca, Bull. Acad. Roumaine, XIV, 1931 : 106—107, первый вид; Beregov, Geol. Balkan, Sofia, III, 1938 : 18—22, все три вида). Некоторые из ископаемых сельдей достигали довольно крупных размеров (22 и 44 см, один образец имеет в длину 58 см без головы) (Богачев, 1933 : 35).
Следовательно, из богатой, чисто морской фауны Тетиса и пришедшей на смену ей под влиянием, верхнетретичного похолодания климата более холодолюбивой фауны, вследствие ряда последовательных опреснений и осолонений в изолированных морях северо-восточной части его, оказалась отобранной небольшая обедненная фауна, дававшая временами в некоторых бассейнах пышный расцвет, сменявшийся угнетением и отчасти вымиранием.
Этот расцвет обычно обусловливается богатством фауны не родами, а видами, что, например, констатировано для фауны моллюсков сарматского моря (Колесников, 1940 : 313). Под влиянием последовательных изменений солености выработалась своеобразная еврибионтная, главным образом евригалинная фауна, представители которой обладают, как будет указано ниже, меньшей жизненностью по сравнению с фауной Средиземного моря и Атлантического океана.
Изоляция в периодически опресняемых бассейнах оказала влияние не только на развитие и формообразование каспийско-черноморских сельдевых, но и на состав их паразитофауны, причем все они освободились от некоторых паразитов, широко распространенных у многих сельдевых Средиземного моря, Атлантического океана, Белого моря и Тихого океана. Так, например, в семенниках большинства сельдевых северного полушария встречается кокцидия Eimeria. Eimeria sardinae или близкие к ней формы известны у сельдевых Средиземного моря и Алтантического океана (Sardina pilchardus sardina, Clupea harengus harengus), Белого моря (Clupea harengus pallasi n. maris-albi) и Тихого океана (Clupea harengus pallasi, Sardinops sagax melanosticta, Etrumeus micropus). Этот паразит встречается также и у Engraulis encrasicholus и его черноморских форм, но у всех черноморско-азовских и каспийских сельдевых из родов Clupeonella и Alosa отсутствует. Повидимому, Eimeria ранее была свойственна и каспийско-черноморским сельдевым, но затем под влиянием опреснений они от нее освободились. В пользу этого предположения говорит тот факт, что у Clupea harengus harengus n. membras в опресненном Финском заливе этого паразита нет (Догель, 1938 : 903; 1941 : 33; 1947 : 298). Вместе с тем некоторые паразиты, свойственные другим сельдям, у каспийско-черноморских сельдей сохранились несомненно с древних времен. Таковой является миксоспоридия Mitraspora caspialosae, обнаруженная у каспийских и черноморско-азовских сельдей. Близкий или идентичный вид этой миксоспоридии известен у Alosa fallax fallax (Догель, 1938 : 903). Ряд паразитов, свойственных черноморско-азовским сельдям из рода Alosa (Alosa kessleri pontica и Alosa caspia nordmanni) и у каспийских сельдей этого рода отсутствующих, повидимому, получен черноморско-азовскими сельдями от других рыб, средиземноморских иммигрантов, уже после соединения Черного моря со Средиземным. К числу таких паразитов относятся черви сосальщики Hemiurus appendiculatus и Lecithaster confusus и паразитические ракообразные Clacellisa marginata и Cymathoa punctata. Помимо широкого распространения в Черном и Азовском морях все эти паразиты встречаются и в Средиземном море, а некоторые — и у европейских берегов Атлантического океана. Отсутствие этих паразитов у каспийских сельдей трудно объяснить иным образом, в частности различиями условий существования их в Каспийском море и в Черном и Азовском морях, так как, например, Hemiurus appendiculatus найден в сельдях, зашедших в Днепровский лиман, a Clacellisa marginata — у сельдей в Азовском море и в низовьях впадающих в него рек (Османов, 1940 : 254).
Биологические взаимоотношения видов Clupeonella и Alosa с другими рыбами как в настоящее время в Каспийском море и Черном и Азовском морях, так несомненно и в морях, предшествовавших им в предыдущие эпохи, различны. Пелагическая фауна открытых частей Каспийского моря состоит исключительно из сельдевых, которые (все виды Clupeonella и формы Alosa caspia s. l.) являются здесь основными потребителями планктона (см. об этом Бенинг, 1938 : 33). Других пелагических рыб, кроме сельдевых, в галистатической области Каспийского моря в сущности нет. Население пелагиали более открытых частей Черного моря составляют в основном средиземноморские иммигранты как из сельдевых, но других родов (Sprattus sprattus phalericus, Sardina pilchardus sardina, Sardinella aurita), так и из других семейств — Engraulis encrasicholus, Trachurus trachurus, некоторые виды Syngnathus (повидимому Syngnathus schmidti и Syngnathus typhle argentatus), личинки и молодь Mugil sp., молодь Mullus barbatus и Odontogadus merlangus euxinus, Scomber scomber, Scomber japonicus, Sarda sarda и некоторые другие рыбы (см. об этом: Ильин, 1933 : 63; Малятский, 1938 : 96; 1940 : 237; Водяницкий, 1939 : 69; 1940 : 29). Последняя из этих рыб (Борисенко, 1937 : 99) и скумбрия (Макаров, 1938 : 33) являются хищниками, питающимися в основном рыбой (скумбрией, хамсой, шпротом, атеринкой), т. е. замещая в этом отношении здесь хищных сельдей Каспийского моря, в частности Alosa brashnikovi s. l. Почти все остальные из этих рыб, за исключением Trachurus trachurus, питающейся крупными ракообразными (Leander, Crangon, Gammaridae) и рыбой (Макаров, 1939 : 1021), и все перечисленные выше сельдевые являются основными потребителями планктона Черного моря. К пелагическим рыбам как Черного (Ильин, 1933 : 63), так и некоторых других морей (Гурвич, 1938 : 140) следует отнести и колюшку (Gasterosteus aculeatus), размножающуюся, однако, у самых берегов. Все эти рыбы населяют те более открытые части Черного моря, в которых виды Clupeonella и Alosa не встречаются, временами подходят в прибрежную область, некоторые из них заходят в Азовское море, многие (Engraulis encrasicholus, Sprattus sprattus phalericus и др.) встречаются в массовых количествах. Что касается сельдевых из родов Clupeonella и Alosa, то они вместе с остальной автохтонной реликтовой фауной понтического озера-моря встречаются в опресненных частях Черного моря и в Азовском море. Тюлька (Clupeonella delicatula delicatula) встречается здесь только в Азовском море, в лиманах и в опресненных участках у берегов северо-западной части Черного моря, Румынии, Болгарии, в Феодосийском зал. и в оз. Палиастоми. Сельди в зимнее время держатся вдоль берегов Кавказа, у берегов Крыма, летом заходят в опресненные части Черного моря и в Азовское море и в реки.
В галистатической части Черного моря ни один из представителей этих двух родов не найден.
Таким образом, виды рода Clupeonella, живущие только в море и притом в открытых его частях (Clupeonella grimmi и Clupeonella engrauliformis), и морские, непроходные виды рода Alosa (Alosa saposhnikovi, Alosa sphaerocephala, Alosa brashnikovi s. l.), мечущие икру в море, в Черном море не могли образоваться или получить развитие потому, что пелагиаль открытых частей его после прорыва Босфора была заселена более сильными конкурентами, иммигрантами из Средиземного моря. Наоборот, можно полагать, что сельдевые из этих двух родов в настоящую фазу существования Черного моря испытывают в нем сильную депрессию, будучи оттеснены в наиболее опресненные части его и в Азовское море более жизненными средиземноморскими иммигрантами. Успешная акклиматизация в Каспийском море кефалей (Mugil auratus и Mugil saliens), креветки Leander adspersus, червя Nereis succinea и моллюска Mytilaster lineatus, причем последний в значительной степени вытесняет автохтонного моллюска Dreissena (повидимому Dreissena caspica), a Leander adspersus каспийских мизид (Жадин 1941: 10; Броцкая и Неценгович, 1941 : 79), показывает, насколько более жизненны средиземноморские иммигранты по сравнению с фауной автохтонной (см. об этом: Зенкевич и Бирштейн, 1934 : 38; 1937 : 443).
Совершенно иные условия оказались в Каспийском море, которое в средней и, в особенности, в южной части в настоящую фазу его существования представляет солоноватоводный, в южной части почти такой же морской бассейн, как Черное море. В этом бассейне пелагических рыб кроме сельдевых из родов Clupeonella и Alosa не оказалось, поэтому фауна пелагиали его, в особенности в южной и средней частях, могла формироваться только из этих рыб, и поэтому же они, в особенности морские непроходные, при отсутствии конкуренции с другими видами и получили здесь пышный расцвет; расцвет этот сопровождается и гигантизмом; аналогичное явление наблюдалось и в понтической реликтовой фауне, например в киммерийском бассейне (Колесников, 1940 : 930). Расцвет в Каспийском море одного из наиболее богатого здесь формами видов, Alosa brashnikovi s. l., в особенности в южной части моря, где локализовано большинство форм его, обусловлен не только отсутствием здесь других пелагических рыб, кроме сельдевых, но также приспособлением некоторых форм его и к придонному образу жизни и питанию. При этом некоторые формы Alosa brashnikovi s. l., помимо существенных различий в образе жизни, характере питания и в связанных с этим некоторых морфологических особенностях, о чем подробно указывалось выше, отличаются большим разнообразием и во времени нереста, имея в этом отношении некоторое сходство с атлантической сельдью (Clupea harengus harengus). Среди форм этого вида имеются как наиболее холодолюбивые (Alosa brashnikovi brashnikovi), так и самые теплолюбивые (Alosa brashnikovi kisselevitshi) из всех каспийских сельдей, причем по окончании нереста одной формы (например Alosa brashnikovi grimmi, в июне и даже в июле) на тех же местах позднее начинают метать икру другие (Alosa brashnikovi kisselevitshi).
Сопоставив условия развития сельдевых в Каспийском, в Черном и Азовском морях, нельзя не коснуться условий развития пелагических рыб в третьем солоноватоводном бассейне — в Аральском море, по солености близком к Каспийскому (S 10.19‰; Бадер, 1934 : 5) и в историческом развитии отчасти связанном с последним. Отсутствие по берегам современного Аральского моря каких-либо древних (нижнечетвертичных и плиоценовых) морских или озерных отложений и распространение древнеаллювиальных отложений в центральной части Турана заставляет предполагать, что Аральское море образовалось лишь во второй половине четвертичного периода (Архангельский, 1931 : 171 и сл.; Герасимов, 1937 : 58). Таким образом, геологически Аральское море значительно моложе понто-каспийского бассейна. Тем не менее условия развития и формирования пелагической фауны Аральского моря имеют некоторое сходство и вместе с тем различия с таковыми в Каспийском. Вопрос о связи Аральского моря с Каспийским в настоящее время не может считаться решенным. В частности остается нерешенным вопрос о времени, когда была трансгрессия с Cardium edule. Кесь (1939 : 1), подвергнувшая этот вопрос детальному анализу, считает возможный утверждать лишь то, что в период максимальной трансгрессии моря с Cardium edule Узбой был еще оводнен. Отсутствие в Аральском море некоторых рыб средиземноморских (Atherina, Syngnathus) и понтических реликтов (бычки из сем. Gobiidae) Берг (1908 : 451) объясняет тем, что это море прошло в своем развитии пресноводную фазу. Никольский (1940 : 192) не считает, однако, это достаточным для объяснения причин отсутствия в Аральском море упомянутых рыб и высказывает мысль, что отсутствие здесь этих рыб обусловлено тем, что ихтиофауна Аральского моря сформировалась из ихтиофауны Аму- и его древнего притока (в период древнего плювиаля; Герасимов, 1937 : 57) Сыр-дарьи, а современные обитатели его представляют собой лимнофильные элементы плейстоценовой ихтиофауны этих рек, и, возможно, что первоначальная фауна Аральского моря была несколько богаче современной. При последовавшем затем осолонении Аральского моря некоторые виды принуждены были уйти в дельты и озера Аму- и Сыр-дарьи, а часть приспособилась к изменившимся условиям и осталась в самом море.
Таким образом, пелагическая фауна рыб в Аральском море могла формироваться только из сравнительно стенобионтных, пресноводных рыб, так как здесь не оказалось в наличии ни средиземноморских рыб, как в Черном море, ни пелагических из понтических реликтов, как в Каспийском море. Пелагических в числе рыб, из которых формировалась ихтиофауна Аральского моря, не оказалось, не оказалось и таких, которые могли бы приспособиться к условиям жизни в пелагиали осолонившегося бассейна. Вследствие этого рыб, постоянно живущих в обширных пространствах открытых частей Аральского моря, подобных видам Clupeonella и Alosa в Каспийском море и Engraulis, Sprattus и др. в Черном, в этом море нет; зоопланктон его в качестве основного объекта питания используется в сущности лишь колюшкой (Pungitius platygaster aralensis), важнейшие же промысловые рыбы (лещ, вобла, судак, белоглазка, чехонь, шемая) хотя и откармливаются в открытой части моря с глубинами от 15 до 30 м, но среди них нет ни одной кормящейся только в пелагиали, так как все они питаются за счет и донной фауны (ракообразные, моллюски), и пелагиаль этого моря остается в сущности незаполненной (Никольский, 1940 : 197). Весьма характерно также то, что ни одна рыба в открытых частях Аральского моря не размножается (Никольский, 1940 : 194).
В Каспийском море, благодаря, как указывалось выше, особым условиям развития и состава его ихтиофауны, сельдевые являются более морскими рыбами, чем формы тех же видов в Черном и Азовском морях. Вследствие тех же причин ряд рыб, которые в черноморско-азовском и каспийском бассейнах являются полупроходными или пресноводными, в Аральском море живут в открытых, осолоненных частях его. Так, например, аральская вобла (Rutilus rutilus aralensis) встречается по всему Аральскому морю, даже в осолоненных заливах его восточного берега (Никольский, 1940 : 74), в то время как каспийская вобла (Rutilus rutilus caspicus) встречается только в опресненной северной части моря и в опресненных заливах южной части его — заливы Гасанкулинский, Астрабадский (Rutilus rutilus caspicus n. knipowitschi), Кизилагачский и, вероятно, Энзелийский (Rutilus rutilus caspicus n. kurensis). Близкая к аральской и каспийской вобле черноморско-азовская, называемая здесь таранью, (Rutilus rutilus heckeli), встречается также только в опресненных частях Черного и Азовского морей. Каспийские формы воблы и тарань мечут икру только в пресной воде (Берг, 1949 : 502, 520, 527, 528), между тем как аральская вобла выметывает икру главным образом в море по всем берегам его как в пресной, так и в солоноватой воде с содержанием солей до 11‰ (Никольский, 1940 : 77). Чехонь (Pelecus cultratus) в Аральском море встречается по всем берегам и при любой солености, в то время как в Каспийском, Черном и Азовском морях распространение ее ограничено предустьевыми, опресненными пространствами. В Каспийском, Черном и Азовском морях чехонь размножается только в пресной воде, аральская же чехонь, повидимому, мечет икру и в солоноватой воде (Никольский, 1940 : 148). То же самое можно сказать и о ряде других рыб Аральского моря. Даже чисто пресноводная рыба — красноперка (Scardinius erythrophthalmus) в Аральском море держится по всем берегам и размножается, по указанию Никольского (1940 : 191), при солености свыше 10‰. Аральская шемая (Chalcalburnus chalcoides aralensis) широко распространена по всему морю, где мечет икру как в опресненных предустьевых пространствах, так, главным образом, по берегам его при содержании солей свыше 10‰ (Никольский, 1940 : 111, 191). Каспийская шемая (Chalcalburnus chalcoides typ.) встречается главным образом в южной части моря, преимущественно в не слишком осолоненных районах ее, размножается только в пресной воде, в реках (Кура, Терек и отчасти другие; Берг, 1949 : 736). Черноморская шемая (Chalcalburnus chalcoidess danubicus и Chalcalburnus chalcoides schischkovi) встречается только в опресненных пространствах Азовского и северо-западной части Черного моря, размножается только в реках, причем в Азовском море ведет себя как настоящая проходная рыба, в Кубань заходит с осени и поднимается очень высоко, до Усть-Лабинской и выше, заходит в притоки (Пшиш, Белая, Лаба; Берг, 1932 : 478). В реках же Крыма и западного Закавказья шемая, отсутствующая у берегов этих мест в море, представлена особыми пресноводными формами (Chalcalburnus chalcoides mentoides, Chalcalburnus chalcoides derjugini; Берг, 1949 : 740, 741).
Таким образом, состав и распределение ихтиофауны в каждом из трех рассматриваемых солоноватоводных морей различны. Вследствие того, что ихтиофауна Аральского моря сформировалась из пресноводных рыб, среди которых настоящих пелагических не оказалось, пелагиаль его остается незаполненной, а открытые части населяют, размножаясь в самом море в прибрежной области, такие рыбы, которые в Каспийском, Азовском и Черном морях держатся преимущественно в опресненных предустьевых пространствах, а размножаются в пресной воде, в реках. Открытые части Каспийского моря населены понтическими реликтами из сельдевых, настоящими пелагическими рыбами, давшими здесь пышный расцвет и особенно богато представленными морскими непроходными видами и подвидами. Часть этих рыб в открытых частях Каспийского моря держится постоянно, здесь же и размножается, многие виды мечут икру в море в прибрежной области, некоторые для икрометания заходят в опресненные предустьевые пространства и в реки. Эти же понтические реликтовые сельдевые, представленные в Черном море значительно меньшим количеством близких подвидов, общих с некоторыми из каспийских видов, в открытых частях этого моря не встречаются, будучи вытесненными отсюда рядом средиземноморских пелагических рыб. Все эти сельдевые в Черном море держатся лишь вблизи берегов и преимущественно в зимнее время, в северо-западной части его и в Азовском море, преобладающее большинство их размножается в пресной воде и лишь один-два представителя — в солоноватой.
Пелагиаль открытых частей Средиземного моря и Атлантического океана, благодаря постоянству состава их ихтиофауны с конца миоцена, была всегда заселена, помимо упомянутых для Черного моря, рядом других пелагических рыб. В частности, в пелагиали бореальной области Атлантического океана наиболее существенную роль играет атлантическая сельдь (Clupea harengus harengus). Поэтому средиземноморско-атлантические сельди из Alosa, не имея подходящих условий для расцвета в прошлом, не имеют и, вероятно, не имели и ранее ни одного вида, который размножался бы в море или в солоноватой воде; все они являются проходными видами. Более того, многие из них образовали чисто пресноводные, речные или озерные формы, которых каспийско-черноморские сельди не имеют. Таковыми являются: Alosa fallax lacustris (озера Маджоре, Комо, Лугано), Alosa fallax gracilis (оз. Гарда), Alosa fallax killarnensis (оз. Килларней в юго-западной части Ирландии), Alosa alosa macedonica (оз. Бешик-гёль, бассейн Эгейского моря), Alosa alabamae ohiensis (р. Огайо, бассейн Миссисипи).
Средиземноморско-атлантические сельди Alosa alosa и Alosa fallax морфологически и биологически соответствуют каспийским проходным сельдям Alosa kessleri kessleri и Alosa kessleri volgensis. Alosa alosa alosa и Alosa kessleri kessleri — крупные и жирные сельди, заходящие в реки с еще не вполне зрелыми половыми продуктами и подымающиеся для нереста высоко вверх по течению. Alosa fallax fallax и Alosa kessleri volgensis — менее крупные и жирные сельди, в реки заходят с вполне зрелыми половыми продуктами и высоко вверх не подымаются (Alosa fallax fallax в Рейне мечет икру в пределах Голландии, Alosa kessleri volgensis — в Волге в главной массе ниже Сталинграда, в то время как обе первые сельди нерестятся значительно выше). Черноморско-азовская проходная сельдь Alosa kessleri pontica, как указывалось выше, распадается на 2 формы (nationes) — крупную и мелкую. Крупная форма в реки входит раньше и с менее зрелыми половыми продуктами, чем мелкая, она более жирна. В настоящее время вследствие возведения плотин на Днепре и на Дону трудно сказать, подымается ли крупная форма Alosa kessleri pontica вверх по течению выше, чем мелкая, но в прежние годы в Днепре на порогах ловилась только крупная сельдь.
Как нетрудно видеть, крупная форма Alosa kessleri pontica биологически соответствует Alosa kessleri kessleri и Alosa alosa alosa, а мелкая — Alosa kessleri volgensis и Alosa fallax fallax, и все эти проходные сельди в трех рассматриваемых морских бассейнах представляют 3 формы разной степени обособления. Благодаря тому что в Атлантическом океане и в Средиземном море с конца миоцена не было таких изменений солености и сопутствующих им изменений в составе ихтиофауны, как в бассейнах, давших начало Черному и Каспийскому морям, в конечном итоге, в силу изменчивости в одном направлении, при которой не появлялись формы, имевшие признаки, отчасти свойственные предшествовавшим формам, отличия между Alosa alosa и Alosa fallax оказались более значительными и они обособились здесь до степени видов, имеющих и качественные различия. Каспийско-черноморские проходные сельди, замкнутые в сарматском и других сменивших его бассейнах, вследствие ряда последовательных опреснений и осолонений в этих морях, вызывавших, как указывалось выше, появление форм, имевших некоторые признаки, отчасти свойственные предыдущим формам, обособились менее. Каспийские сельди, являющиеся в настоящую фазу существования Каспийского моря единственными пелагическими рыбами, обособились до степени подвидов, имеющих лишь количественные отличия, в то время как черноморско-азовские проходные сельди, оттесненные средиземноморскими иммигрантами в опресненные части Черного моря и в Азовское море и находящиеся в менее благоприятных условиях, обособились менее, до степени менее обособленных форм (nationes). Большей степенью различий, чем черноморско-азовские, отличаются, как указывалось выше, и другие каспийские формы общих с черноморско-азовскими формами видов (Alosa caspia s. l., Alosa brashnikovi s. l.). Разная степень различия между видами подрода Alosa s. str. и формами каспийских и черноморско-азовских проходных сельдей подрода Caspialosa наблюдаются, между прочим, и в количестве лучей в анальном плавнике. Количество лучей в анальном плавнике у всех, даже у видов Alosa, трансгрессивно, но разница в средних у видов Alosa значительно больше, чем у подвидов и более низких форм (nationes) Alosa, что видно из нижеследующего: Alosa alosa alosa — 23.1, Alosa fallax fallax — 22.0, Alosa kessleri kessleri — 21.1, Alosa kessleri volgensis — 20.8, Alosa kessleri pontica крупная форма — 20.7, Alosa kessleri pontica мелкая форма — 20.5.
Подводя итоги всему сказанному об условиях развития и формообразования сельдевых из рода Clupeonella и Alosa, следует подчеркнуть, что богатство их в Каспийском море обусловлено расцветом их в этом море главным образом в настоящую фазу его существования, со времени последнего соединения его с морями, предшествовавшими Черному и Азовскому, которое могло быть в послехвалынское и в бакинское, а ранее в апшеронское и акчагыльское время (см. об этом: Колесников, 1940 : 928), а не в одном из морей, предшествовавших Каспийскому, как полагал Киселевич (1918 : 24). При этом расцвет их обусловили не «значительное протяжение и разнообразие климатических условий в различных частях бассейна» (Киселевич, 1918 : 24), а наличие в Каспийском море незаселенной другими рыбами пелагиали, которую могли заполнить только сельдевые рассматриваемых двух родов. Что касается приводимого К. А. Киселевичем мнения Сарса, что каспийские сельди «находятся в стадии незакончивщегося видообразования» (1918 : 24), то в стадии видообразования находится весь органический мир, даже наиболее древние его представители.
Среди других видов рода Clupeonella по своему распространению особый интерес представляет Clupeonella abrau. Этот вид встречается в небольшом пресноводном озере Абрау и, кроме того, в качестве особого вида (Clupeonella muhlisi), идентичного Clupeonella abrau или самое большее являющегося формой этого вида, описан из пресноводного озера Абулионд (басс. Мраморного моря).
О реликтовом характере оз. Абрау, расположенного на значительной (70 м) высоте над современным уровнем Черного моря, высказался первый исследователь его Кричагин (1877 : 14). Его точка зрения была поддержана и подкреплена новыми фактами рядом других исследователей (Чернявский, 1879 : XI; Совинский, 1903 : 36; Державин, 1924 : 113; 1925 : 10; Фадеев, 1925 : 13; Малятский, 1930 : 70; Водяницкий, 1930 : 132), изучавших фауну оз. Абрау. Правда, высказывались и иные взгляды на происхождение этого озера (Скориков, 1904 : XIX; Арнольди, 1922 : 47), однако против них были сделаны серьезные возражения, и реликтовый характер фауны оз. Абрау в настоящее время более не оспаривается. Наличие в оз. Абрау некоторых форм с определенно выраженным реликтовым характером и среди них в особенности Clupeonella abrau приводит к необходимости признать фауну этого озера за реликтовую.
Относительно времени, когда оз. Абрау потеряло связь с морским солоноватоводным бассейном, высказывались предположения, что это могло произойти в начале плиоцена (связь с понтическим озером-морем; Милятский, 1930 : 73) или в постплиоценовое, а может быть, и в более позднее время (Державин, 1924). Геологическое строение Абрауского п-ова не дает указаний на непосредственную связь оз. Абрау с древним солоноватоводным морем (Ивченко, 1923), однако Державин (1924) на основании нахождения к юго-востоку от Новороссийска (между Геленджиком и Джубгою) и далее постплиоценовых террас с остатками современных каспийских моллюсков и средиземноморских Balanus считает, что реликтовый характер озера не стоит в противоречии с геологическими фактами.
Большинство форм оз. Абрау, которые по происхождению своему связаны с морем (Heterope caspia, Ectinosoma abrau, Gammarus robustoides, Corophium curvispinum, Jaera nordmanni, Mesomysis kovalevskyi, Theodoxus fluviatilis abrauensis, Clupeonella abrau), будучи в основной своей массе автохтонными реликтами понтического озера-моря, имеют довольно широкое распространение помимо оз. Абрау. Эндемичным считается лишь Theodoxus fluviatilis abrauensis, причем Водяницкий (1930 : 133), со слов В. И. Жадина, указывает, что он чрезвычайно близок к встречающемуся в солоноватых частях Аральского, Каспийского, Азовского и Черного морей Theodoxus pallasi ( = Neritina liturata Eichw.), которого Жадин (1933 : 184) считает родоначальной формой Theodoxus danubialis. Theodoxus fluviatilis abrauensis является вообще формой мало изученной, обладающей рядом промежуточных признаков между Theodoxus fluviatilis и Theodoxus danubialis, вследствие чего Жадин (1933 : 184) относит его частично к последнему виду. Что касается Clupeonella abrau, то он не может считаться эндемичным потому, что, как указывалось выше, найден и в оз. Абулионд. По ряду морфологических особенностей, положенных в основу системы семейства и отдельных видов, Clupeonella abrau относится к числу наиболее примитивных видов рода и наиболее близок не к встречающемуся также и в Черном и Азовском морях виду Clupeonella delicatula, а к видам, встречающимся только в Каспийском море.
Геологических указаний на былую связь оз. Абрау с морским бассейном, как уже указывалось, нет, точно так же как не известно и время, когда в него проник Clupeonella abrau. Однако все изложенное выше все же дает возможность относительно этого высказать некоторые соображения. Из числа нескольких предположений, которые можно высказать на этот счет, наиболее вероятно то, что форма, давшая Clupeonella abrau, заселила озера Абрау и Абулионд на самых ранних этапах видообразования, до образования современных видов рода, так как в противном случае оз. Абрау, несомненно, оказалось бы заселенным формой, наиболее близкой к прибрежному и евригалинному виду — Clupeonella delicatula, который при наличии подходящих условий образует пресноводные формы (m. tsharchalensis). Далее, можно предполагать, что проникновение формы, давшей начало Clupeonella abrau в оз. Абрау, произошло, повидимому, во время одного из соединений Черного и Каспийского морей, так как при повышении современного уровня Черного моря до уровня оз. Абрау долина р. Маныча должна была бы быть затопленной, вследствие чего эти моря соединились бы друг с другом. Такие условия после понтического озера-моря позднее были, как указывалось выше, в акчагыльское, ашперонское, а в четвертичный период в бакинское и послехвалынскоег время (Колесников, 1940 : 928; Архангельский и Страхов, 1938 : 165—192). Один из примитивных видов одного из этих времен и дал начало Clupeonella abrau, который в условиях изоляции сохранил ряд особенностей, свойственных наиболее примитивным видам рода, причем у него выработались своеобразные и биологические особенности.
БИОЛОГИЯ
Биологическими особенностями сельдевых, как указывалось в двух предыдущих разделах, в значительной мере обусловлено их географическое распространение и пути расселения, точно так же как в известной степени с биологическими же особенностями связаны и систематические отношения сельдевых. Биологические особенности обусловливают и характер хозяйственного использования сельдевых — места, время и способы лова. Не менее важное значение для промысла имеет ряд других рассматриваемых ниже биологических особенностей.
Почти каждую из рассматриваемых ниже биологических особенностей во взаимоотношениях со средой, как то отношение к температуре, солености, глубине и др., имеет известный размах колебаний между крайними пределами, к которым приспособлен какой-либо вид. У одних видов этот размах колебаний более, у других менее широк; наименее узок он в период размножения, и в сущности характером отношения к основным факторам среды в этот период и определяется в значительной мере как биологический облик вида, так и ряд других его особенностей, в частности ареал (Световидов, 1949) и характер промысла.
Отношение сельдевых к температуре и солености охарактеризовано отчасти выше (табл. 11).
По отношению к температуре семейство довольно разнородно, однако в основном, по преобладающему большинству представителей, его следует считать достаточно теплолюбивым, так как подавляющее количество сельдевых, как указывалось, является тропическими формами.
По общему характеру отношения к температуре сельдевых следует отнести к наиболее евритермным семействам костистых рыб. Действительно среди морских рыб распространение большинства тропических семейств ограничено тропиками, и лишь немногие из них встречаются в морях субтропических и более высоких широт. Сельдевые благодаря широкой евритермности семейства в целом являются единственным семейством среди костистых рыб, распространение которого охватывает моря почти всех широт, исключая высокоарктические.
Сельдевые встречаются даже при отрицательных температурах воды. При таких температурах держатся в зимнее время, и отмечено даже начало нерестовых подходов малопозвонковых форм Clupea harengus (например у Clupea harengus pallasi n. maris-albi, Clupea harengus pallasi). Это один из самых холодолюбивых видов семейства, ряд форм которого зимнее время проводит подо льдом. Указать верхний предел температуры, при которой встречаются сельдевые, затруднительно, так как наиболее теплолюбивые, тропические, виды в этом отношении почти не изучались. У Clupea harengus по сравнению с другими сельдевыми и икрометание происходит при наиболее низкой температуре воды: у малопозвонковых форм, в частности у Clupea harengus pallasi, нерест начинается при 2°, а у некоторых форм (Clupea harengus pallasi n. maris-albi) — начиная от 0°, а все развитие икры протекает при температуре от 0 до 5°. Многопозвонковые формы Clupea harengus размножаются при несколько более высокой температуре, причем у Clupea harengus harengus нерест осенних сельдей происходит при более высокой температуре воды, чем весенних. Икрометание осенних сельдей происходит при температуре до 14°. Нерест сельдевых, размножающихся в пресных водах, проходных и пресноводных, происходит при температуре воды, значительно более высокой, чем морских. В качестве примера приведем роды Sprattus и Alosa, занимающие в море приблизительно одинаковый в меридиональном направлении ареал. Sprattus sprattus phalericus размножается в Черном и Адриатическом морях при температуре 6—10°, Alosa fallax nilotica — в реках Италии при 22—25°. Ареал Sardinops sagax в северной части Тихого океана расположен несколько южнее, чем ареал Alosa sapidissima у тихоокеанских берегов Сев. Америки, однако нерест первой происходит при более низкой температуре, чем нерест второй. Калифорнийская сардина (Sardinops sagax caerulea), как и дальневосточная (Sardinops sagax melanosticta), нерестует в основном при 13—15°, Alosa sapidissima — при 14—18°. При еще более высокой температуре протекает размножение и развитие икры Hilsa reeversii, встречаемой в южной части ареала (от Хайнана до Манчжурии) Sardinops sagax melanosticta. Развитие икры Hilsa reeversii происходит при 28°, между тем как икра ряда других сельдевых погибает при значительно более низкой температуре. Это различие в температурах икрометания и развития икры морских, с одной стороны, и пресноводных и проходных сельдевых, с другой, обусловлено более высокими температурами в пресных водах по сравнению с температурами воды в морях тех же широт и является важным биологическим приспособлением. Пелагическая икра сельдевых в большей степени подвержена гибельным воздействиям в реке, чем в море. Развитие же икры при высокой температуре, как известно, происходит быстрее, чем при низкой. Таким образом, более быстрое развитие икры сельдевых в пресных водах является крайне важным приспособлением, способствующим уменьшению гибели ее при развитии, а следовательно, и обеспечению численности вида. Для икры, откладываемой на дно или другой субстрат, наоборот, условия развития в море неблагоприятны вследствие обилия здесь врагов ее. Условия для развития такой икры более благоприятны в реках, чем и обусловлены анадромные нерестовые миграции (Васнецов, Зоолог. журн., XXIII, 5, 1944 : 198—211; см. также: Крыжановский, Тр. Инст. морфол. жив., I, 1948).
Всякое изменение температуры воды оказывает весьма существенное влияние на сельдевых. В частности Clupea harengus harengus крайне чутко реагирует даже на ничтожное изменение температуры воды до 0.2° (Shelford a. Powers, 1915). Особенно резко влияет на сельдевых быстрое понижение температуры. Так, Sardinops sagax melanosticta при быстром понижении температуры воды погибает при 6°, при медленном же выживает и при 4° и даже при 2°. Сезонные изменения температуры, под влиянием которых рыбы перемещаются в другие районы моря или в другие слои по вертикали, обусловливают в основном миграции сельдевых, их вертикальное распределение, места и время размножения, многочисленные примеры чего приведены в описании биологии каждого вида и формы сельдевых в «Систематической части». Изменения температурного режима моря в отдельные годы или под влиянием изменения режима океанических течений более длительные, на протяжении ряда лет, вызывают существенные изменения в распространении, а также и в состоянии численности отдельных видов и форм сельдевых. Изменение температурного режима моря в отдельные годы оказывает влияние на урожайность поколений этих лет. Так, поколения Clupea harengus pallasi n. maris-albi в годы с низкой температурой воды Белого моря отличаются большой численностью и рядом других особенностей, поколения годов с более высокой температурой — малой численностью. В более теплые годы в Белое море происходит в большем количестве заход Clupea harengus harengus, подвида более теплолюбивого, чем предыдущий. Особенно существенное влияние оказывают длительные изменения температурного режима моря. Так, в 1941—1942 гг. под влиянием похолодания центральной части Японского моря катастрофически снизились в наших водах уловы дальневосточной сардины (Sardinops sagax melanosticta), являвшейся до этого важнейшей промысловой рыбой, а в 1943—1944 гг. она встречалась лишь единично. Исчезновение сардины под влиянием похолодания положило конец неясности в вопросе о том, была ли она в наших водах в промысловых количествах ранее 1925 г., когда начался ее промысел, или она пришла сюда вследствие наблюдавшегося в эти годы потепления. Эти изменения температурного режима моря оказали существенное влияние на распространение и численность и тихоокеанской сельди (Clupea harengus pallasi), в частности ловимой у берегов южной части Сахалина и у Хоккайдо (южносахалинско-хоккайдская сельдь). Потепление, начавшееся в 1922—4923 гг., вызвало перемещение южносахалинско-хоккайдской сельди из южных частей ареала в восточные (зал. Анива) и отчасти северные (западное побережье южной части Сахалина). Численность и общие уловы ее при этом не изменились, так как уменьшению уловов в одних районах сопутствовало увеличение их в других. Однако уловы к концу этой эпохи потепления резко снизились во всех промысловых районах. Наступление следующей эпохи похолодания обусловило некоторое увеличение численности южносахалинско-хоккайдской сельди, но сравнительно небольшое, так как меньшее количество нерестовой сельди вследствие небольшой численности ее не могло дать таких поколений, которые давала эта сельдь в годы большой численности.
В теснейшей зависимости от температуры воды на ранних стадиях развития личинок находится и количество позвонков у сельдевых, как и у всех других рыб. Как известно, средние числа позвонков у поколений разных годов одной и той же сельди различаются, хотя и в небольшой степени. Установлено (Rounsefell a. Dahlgren, 1932; Tester, 1938), что среднее число позвонков находится в зависимости от температуры воды, при которой происходит развитие личинок: у поколений сельди в годы с низкой температурой воды в период развития личинок среднее число познонков больше, у поколений годов с более высокой температурой — меньше. Этой зависимостью количества позвонков от температурных условий в период развития личинок объясняется и кажущееся на первый взгляд противоречие общему правилу распространение у Clupea harengus harengus многопозвонкового и малопозвонкового подвидов. Известно, что как среди морских рыб северные роды и виды имеют большее количество позвонков, чем южные, так и внутри вида у северных и океанических форм количество позвонков обычно также больше, чем у более южных и прибрежных (сводку этого вопроса см. у Расса, 1941). У Clupea harengus наблюдается обратное — малопозвонковые формы, в частности Clupea harengus pallasi n. maris-albi и Clupea harengus pallasi n. suworowi, имеют более северное распространение, чем многопозвонковые. Хотя икрометание многопозвонковых сельдей и происходит при более высокой температуре воды, чем малопозвонковых, однако вследствие того, что икра их откладывается на значительной глубине и в отдалении от берегов, где температура воды на протяжении длительного периода развития икры меняется мало, развитие личинок происходит при температуре, мало отличающейся от температуры нереста. У малопозвонковых сельдей, несмотря на то, что они нерестуют при более низкой температуре, развитие личинок в узко прибрежной области, где они откладывают икру, в условиях быстрого прогревания и нарастания здесь температуры воды происходит при более высокой температуре, чем у многопозвонковых. Более высокие температурные условия, при которых происходит развитие личинок и формирование позвоночника малопозвонковых сельдей, и обусловили на протяжении длительного периода существования наличие у них меньшего числа позвонков, несмотря на более северное распространение, чем многопозвонковых. У Clupea harengus harengus икрометание осенних сельдей, как указывалось выше, происходит при более высокой температуре воды, чем весенних, однако позвонков у них меньше, чем у последних. Обусловлено это тем, что, несмотря на более высокую температуру нереста, развитие личинок осенних сельдей происходит зимой или поздней осенью уже в сильно охлажденной воде, развитие же весенних — в условиях слабо нарастающей или неизменной температуры воды.
Наиболее различно отношение сельдевых к солености. Несмотря на значительное преобладание морских видов, количество которых превышает половину общего числа их, среди сельдевых имеется также большое количество как проходных, так и пресноводных (табл. 11). В этом отношении сельдевые почти не имеют себе подобных среди других семейств костистых рыб. Так, среди морских рыб преобладающее большинство семейств является исключительно морскими или отчасти и солоноватоводными, размножение преобладающего большинства их, если даже представители некоторых из них и заходят в пресные воды, происходит в морской воде. Точно так же и среди пресноводных рыб большинство семейств является чисто пресноводными, ряд видов и даже родов некоторых семейств является проходными, и лишь немногие виды некоторых семейств живут и размножаются в солоноватой воде. Из числа семейств, обитающих в основном в море, но представленных также и в пресных водах, можно назвать сем. Cottidae, Gobiidae и Osmeridae. Первое семейство имеет пресноводные виды и роды, второе — только виды, последнее — формы проходных видов. Проходные виды имеет лишь последнее семейство, у двух первых их нет. Сем. Clupeidae, представленное в пресных водах большим количеством видов и родов, чем первые два семейства, имеет значительно большее количество и проходных видов и родов, чем последнее. Из других евригалинных семейств укажем сем. Atherinidae и Mugilidae.
Особенно характерен для сельдевых рассмотренный в предыдущей главе разнородный и смешанный характер но отношению к солености не только семейства в целом, но и отдельных родов и даже видов, причем некоторые виды и формы и сами по себе являются в значительной степени евригалинными. Из числа таких видов укажем Alosa caspia, одни формы которого (Alosa caspia salina) живут и размножаются в море, встречаясь местами (в зал. Мертвый Култук) при содержании солей до 34.1— 35.8‰, другие (Alosa caspia caspia и др.) — как в пресной, так и в солоноватой воде. Clupeonella delicatula caspia также живет и размножается как в солоноватой воде в море, так и в пресной. Не имея возможности выйти из волжских затонов и ильменей в море, Clupeonella delicatula caspia остается в них на зиму, причем пресноводная форма (m. tscharchalensis) живет постоянно в оз. Чархал.
При переходе проходных сельдей из морской воды в пресную осмотическое давление крови у них падает. Благодаря этому разница между осмотическим давлением крови и воды в море и в реке остается приблизительно одинаковой, у Alosa caspia caspia равная 0.666—0.671. Эта разница несколько увеличивается, достигая своего максимума (до 0.747), на границе между морской водой и пресной, и стимулирует регуляторный механизм, позволяющий рыбе существовать как в соленой, так и в пресной воде. Регуляторный механизм начинает функционировать не сразу, а спустя некоторое время (Бетешева, 1938), и вследствие этого происходит задержинание и скопление проходных сельдей на границе морской воды и пресной, что имеет важное значение для промысла.
Сельдевые являются пелагическими рыбами поверхностных слоев воды. Однако почти все представители семейства, более или менее изученные в этом отношении, в определенные периоды жизни и при некоторых условиях опускаются на глубины, иногда довольно значительные. В частности, Clupea harengus harengus опускается на глубины до 300 м, Sardinops sagax melanosticta находили в желудках рыб (Scombrops chilodipteroides, Pterothrissus gisu и др.), выловленных на значительных глубинах. Нахождение сельдевых на различных глубинах в разные сезоны и в разные периоды их жизни обусловлено в основном температурой воды. Так, тихоокеанская сельдь (Clupea harengus pallasi) весной, по мере прогревания поверхностных слоев воды и по мере созревания половых продуктов, подымается в поверхностные слои воды из более глубоких, где она находилась зимой. Более теплолюбивая дальневосточная сардина (Sardinops sagax melanosticta), наоборот, в начале лета держится в узко поверхностном и более прогретом слое воды. В дальнейшем, с прогреванием глубоких слоев воды она распространяется и глубже, занимая больший по вертикали горизонт воды. Равным образом ряд сельдевых при повышении температуры поверхностных слоев воды выше известного для каждого вида предела опускается в нижние горизонты с более низкой температурой. Нахождение сельдевых на разных глубинах на протяжении суток обусловливается вертикальными миграциями пищи — планктона, вызванными, в свою очередь, влиянием света. Для ряда планктоноядных сельдевых установлено, что в темное время суток они держатся в поверхностных слоях воды, опускаясь днем в более глубокие горизонты.
Влияние света обусловливает не только вертикальные суточные миграции, но в значительной степени и половое созревание сельдевых. Так, например, интенсивное развитие половых желез каспийских сельдей из рода Alosa начинается с марта месяца, когда температура воды по сравнению с зимними месяцами не изменяется, а интенсивность солнечного сияния резко возрастает (Смирнов, 1946). В связи с влиянием солнечного освещения на сельдевых следует отметить воздействие на них света, испускаемого искусственными источниками освещения. Свойство сельдевых положительно реагировать на свет применяется для лова некоторых из них. В последнее время этот вопрос, имеющий перспективы использования его промыслом, успешно разрабатывается на ряде сельдевых (виды рода Clupeonella, Sprattus sprattus и др.) и уже дал положительные результаты (ссылки на литературу см. при описании указанных видов).
Сельдевые, подобно ряду других рыб, обладают хорошо развитым электротаксисом. Так, Clupea harengus harengus в условиях опыта в огороженных участках моря или в заливах весьма чутко реагирует на электрический ток, поворачиваясь головой в сторону анода и перемещаясь в электрическом поле постоянного тока только в этом направлении, причем стаи можно удерживать на месте или передвигать в определенном направлении. Сельдь реагирует на электрический ток при широком колебании градиентов потенциала электрического поля от ничтожных до высоких, наиболее сильно при последних. В недавнее время обнаружено, что в морях существует природный электрический ток, при этом установлено, что электрический ток в море течет непрерывно, сохраняя некоторое постоянство преимущественного направления и имея суточную периодичность колебания в усиленном развитии быстрых пульсаций в часы, близкие к полудню, и в некотором повышении их вечером. Электрические токи в море имеют большую плотность, вполне достаточную для воздействия на сельдь. Этот факт (Миронов, 1948), повидимому, поможет объяснить некоторые еще недостаточно ясные стороны биологии сельдевых и других морских рыб, в частности причины заходов летней и жирной сельди в губы. Установлено, что разные виды рыб неодинаково чувствительны к электрическому току, причем более чувствительны виды, обладающие более развитой сетью нервных окончаний в коже (Бодрова и Краюхин, 1948; Краюхин и Литвинова, 1950).
Преобладающее большинство сельдевых планктоноядные, сравнительно немногие из них хищники. К числу планктоноядных относятся виды, имеющие наибольшее хозяйственное значение: тихоокеанская сардина (Sardinops sagax), океаническая сельдь (Clupea harengus), из каспийских сельдей каспийский пузанок (Alosa caspia) и др. Некоторые из планктоноядных сельдей, в частности волжская сельдь (Alosa kessleri volgensis), питаются отчасти и мелкой рыбой. К числу хищных сельдей, питающихся рыбой, относятся преимущественно крупные виды (Alosa kessleri kessleri, Alosa brashnikovi), хотя все же самые большие по размерам сельди являются планктоноядными (например Alosa sapidissima). Характер питания, как известно, теснейшим образом связан со строением жаберного аппарата сельдевых — у всех планктоноядных видов количество жаберных тычинок значительно больше, а зубы отсутствуют или развиты менее, чем у хищных или у питающихся крупными ракообразными (например Alosa saposhnikovi). Связь с характером питания имеется и в строении пищеварительного тракта. Так, например, количество пилорических придатков у каспийских сельдей из рода Alosa находится в обратной занисимости от состава пищи, чем количество жаберных тычинок: у планктоноядных видов пилорических придатков меньше, чем у питающихся более крупной пищей хищных (Световидов, 1932). У детритоядных представителей семейства, о которых говорится ниже, как и у сходных по характеру пищи кефалей, желудок имеет мускулистую часть, подобную зобу птиц, кишечник длинный, расположенный в несколько петель.
Среди планктоноядных многотычинковых сельдевых имеются и такие, которые в значительной степени питаются, кроме мелких планктонных организмов, как взвешенным в воде, так, судя по строению рта (как у леща) и захватываемым со дна детритом, заглатывая вместе с ним и ил. Жаберные тычинки, которые отфильтровывают частицы детрита и ила, у этих сельдевых (род Dorosoma и, повидимому, другие представители посдсем. Dorosomatinae) особенно многочисленные и тонкие, как и у кефалей, имеющих сходное питание; зубы отсутствуют. Многочисленные жаберные тычинки не только отфильтровывают пищу, но, судя по всему, играют также роль фильтра, предохраняющего жабры от засорения взвешенными в воде детритными и иловыми частицами. Повидимому, этим следует объяснить распределение в Каспийском море более многотычинковых форм Alosa caspia у западных, а малотычинковых — у восточных берегов (рис. 9). В воды у восточных берегов Каспийского моря почти никаких взвешенных частиц с суши не поступает, так как притоков здесь нет. К западным же берегам моря большое количество детритных и иловых частиц выносится из Урала и Волги и сносится течением вдоль кавказского побережья к югу. В южной части моря сюда поступают еще более мутные воды Куры. Повидимому, вследствие этого жаберных тычинок особенно много у Alosa caspia knipowitschi, локализованной в самой юго-западной части Каспийского моря, меньше у Alosa caspia caspia от западных берегов и менее всего у Alosa caspia persica и Alosa caspia salina от восточных берегов. Сходное распределение имеют и черноморско-азовские формы Alosa caspia (рис. 9): более многотычинковая форма (Alosa caspia nordmanni) живет в северо-западной части Черного моря, куда поступают выносы из многочисленных впадающих сюда рек (подробнее см.: Световидов, 1950 : 421).
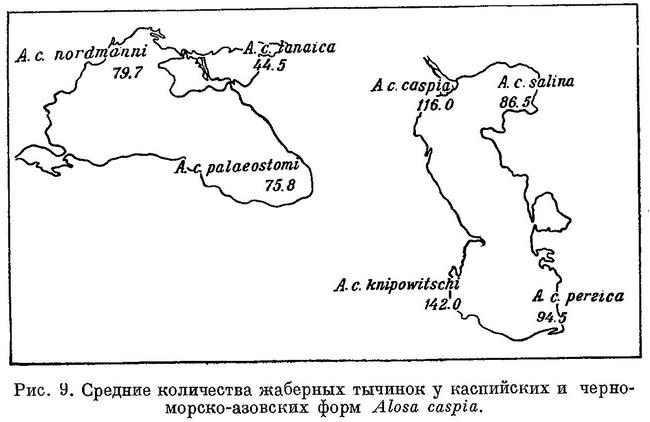
Интенсивность питания сельдевых на протяжении года неодинакова. Так, Clupea harengus в период нереста не питается, питается весной перед нерестом и особенно интенсивно летом после икрометания. В период нереста не питается и ряд других сельдевых, в частности почти все виды каспийских сельдей из рода Alosa.
Икринки преобладающего большинства сельдевых пелагические, плавающие в поверхностных или в придонных слоях воды, у немногих (роды Clupea, Pomolobus) икра клейкая, приклеивающаяся к подводному субстрату. В придонных слоях воды происходит развитие икры большинства сельдевых, размножающихся в пресных водах, в частности проходных сельдей из рода Alosa. Так, например, икринки Alosa kessleri volgensis, имеющие удельный вес немного меньше единицы, в спокойной воде лежат на дне, легко поднимаясь вверх при перемешивании ее. В реке икринки этих сельдей находятся во взвешенном состоянии в придонных слоях воды, сносясь вниз течением. Развитие икры некоторых пресноводных сельдевых (Clupeonella abrau) происходит в поверхностных слоях воды. Плодовитость сельдевых в зависимости от размеров колеблется в широких пределах. Наименьшей плодовитостью отличаются мелкие по размерам виды, наибольшей — крупные. Наименее всего икринок (от 525 до 2416, в среднем 1.4 тыс.) имеет Sprattus sprattus phalericus, обычные размеры которого 60—80 мм, наибольшее (от 135 до 312, в среднем 218 тыс.) — одна из самых крупных сельдей, Alosa kessleri kessleri. Виды, размножающиеся в пресных водах, при приблизительно равной величине имеют большую плодовитость, чем виды, икрометание которых происходит в море. В частности, среди каспийских сельдей у Alosa brashnikovi brashnikovi (размеры наичаще 20—35, максимально до 49 см), размножающейся в море, плодовитость равна в среднем около 66 тыс. икринок; у проходных же сельдей количество икринок таково: у Alosa kessleri volgensis (размеры наичаще около 27—29, наибольшие до 39 см) в среднем около 179 тыс., а у Alosa kessleri kessleri (размеры наичаще около 36—44), как указывалось, значительно больше. Однако южнокаспийские формы Alosa brashnikovi созревают раньше, в подавляющей массе в двухгодовалом возрасте, и поэтому их абсолютная плодовитость выше, чем у проходных сельдей, созревающих позднее (см.: Замахаев, 1949 : 550). Это различие в плодовитости сельдевых, размножающихся в море и в реках, имеет такое же важное приспособительное значение для обеспечения численности вида, как и отмеченные выше различия в температурах икрометания и длительности развития их икры: для сельдевых, размножающихся в реках, в условиях большей возможности гибели развивающейся икры, большая плодовитость имеет крайне важное значение. Такое же значение имеет и порционное икрометание, свойственное многим, если не преобладающему большинству, или всем сельдевым, размножающимся как в пресных водах, так и в море. Последовательное, а не одновременное созревание и выметывание икры дает возможность вместить в полости тела большее ее количество, чем при одновременном. Плодовитость видов, имеющих приклеивающуюся икру, больше, чем видов с пелагической икрой. Так, у Clupea harengus средние количества икринок в зависимости от размеров подвидов колеблются от 6.9 до 72.2 тыс. У Sardinops sagax melanosticta, несмотря на ее почти такие же размеры (15—24 см), как у самых мелких форм Clupea harengus pallasi n. maris-albi (12—20 см), среднее количество икринок больше (45 тыс.) по сравнению с некоторыми даже крупными формами Clupea harengus.
Икрометание сельдевых происходит вблизи берегов, иногда у самого берега или в заливах и бухтах. В открытых частях моря на большом отдалении от берегов сельдевые как правило не размножаются. Места и время икрометания сельдевых определяются по 1) нахождению текучих или близких к зрелости особей, 2) наичаще по уловам оплодотворенной икры, в зависимости от особенностей ее и условий нерестилищ, в понерхностных или придонных слоях воды или у видов с клейкой икрой по нахождению ее на дне, 3) по нахождению личинок на первых стадиях развития, 4) у Clupea harengus, икра которой поедается пикшей, но нахождению в желудках этой рыбы икры сельдевых. Места икрометания сельдевых, размножающихся в море над небольшими глубинами или с приклеивающейся икрой, расположены, за немногими исключениями, на жестких или полужестких грунтах. В частности, среди каспийских сельдей из рода Alosa, размножающихся на небольших глубинах, на местах с мягкими и илистыми грунтами размножается лишь Alosa brashnikovi agrachanica. Точно так же и Clupea harengus откладывает икру на илистые и песчано-илистые грунты в тех случаях (Clupea harengus harengus), когда дно покрыто растительностью. Места икрометания сельдевых размножающихся в пресных водах, в частности проходных из рода Alosa, расположены как в реках, так и в их придаточных системах — в ильменях, на полоях и пр. В этом отношении виды этого рода делятся на две группы — реофильных и стагнофильных, соответствующих группам сельдей и пузанков. Сельди размножаются в основном в реках. Так, например, исключительно по руслу Дона происходит икрометание Alosa kessleri pontica. Alosa kessleri volgensis в низовьях Волги размножается в реках дельты, а не в ильменях. Нерест Alosa kessleri kessleri происходит на значительном протяжении Волги как в местах с замедленным течением, так и на стрежне и, кроме того, отчасти в затонах. В ильменях, на полоях, в лиманах и пр. размножаются пузанки. Нерестилища Alosa caspia caspia расположены в мелководной северной части моря, в основном в предустьевом пространстве Волги, отчасти в ильменях дельты Волги. Alosa caspia tanaica мечет икру на разливах притоков Дона, в лиманах дельты Кубани, отчасти перед устьями Дона. Таким образом, деление видов рода Alosa на пузанков и сельдей соответствует и их биологическим различиям. Следует отметить, что в реках с быстрым течением и, кроме того, с большой мутностью воды за счет неорганических взвесей размножения сельдевых не происходит (мутность воды, вызываемая органическими взвесями, большой роли не играет), поэтому проходные сельди из рода Alosa не идут для нереста в такие быстрые и мутные реки, как, например, Кура, Кубань; небольшое их количество заходит в Урал, а в основном они размножаются в Волге, а также в Дону, Днепре и других менее быстрых и мутных реках бассейна Черного моря.
Отношение сельдевых в период икрометания к температуре охарактеризовано отчасти выше. Здесь следует отметить, что развитие икринок в естественных условиях происходит при оптимальных условиях, крайние пределы которых в условиях опыта значительно шире. В частности, крайние пределы температуры нахождения икринок Alosa kessleri volgensis в естественных условиях колеблются от 12.7 до 25—27°, в условиях же опыта от 10.5—12 до 31—32°. При развитии икринок при низкой температуре воды размеры выклюнувшихся личинок больше и морфологически они более развиты, чем личинки, вышедшие из икринок, развивающихся при более высокой температуре. Например, в районе Исландии только выклюнувшиеся личинки Clupea harengus harengus имеют длину около 6 мм, в более южных районах — около 5 мм. Длительность периода развития икринок у представителей семейства, как указывалось, неодинакова. Наиболее длителен он у самого холодолюбивого вида — Clupea harengus. При 0.1° икринки Clupea harengus harengus проходят развитие в течение 47—50 суток, при наиболее высокой температуре — 14—19° — в течение 6—8 суток. Продолжительность развития икринок Clupeonella delicatala как в Каспийском, так и в Азовском морях при наиболее обычных и естественных условиях температуры воды в период разгара нереста и развития икры (10—22°) колеблется от 98 до 25 час. У размножающейся в море Alosa brashnikovi brashnikovi развитие икринок при температуре разгара нереста (около 17°) длится от 67 до 77 час., у проходной Alosa kessleri kessleri при наиболее обычной температуре в период развития ее икры в реке (около 19°) — в течение 49 час.
Отношение сельдевых к солености наиболее характерно проявляется в период икрометания и развития икры. Ряд видов размножается в воде с нормальной океанической соленостью, причем у некоторых икрометание происходит даже при повышенном содержании солей. Так, например, у Sardinella aurita нерест и развитие икры происходит в среднем при солености 38‰, у Sardina pilchardus pilchardus — в среднем при 35.27—36.14‰, у Sardina pilchardus sardina в Средиземном море при 36.55—38.24‰, но последний размножается и в Черном море при значительно более низкой концентрации солей. Некоторые виды размножаются при еще более широком колебании солености. Так, икрометание одних форм Clupea harengus harengus происходит при нормальной океанической солености, других — при пониженной до 8‰. При еще более низкой концентрации солей размножается Clupea harengus harengus n. membras. Развитие икринок Clupea harengus pallasi n. maris-albi на нерестилищах протекает при солености от 21.3 до 26.0‰; развитие может происходить и при значительно более высокой солености — до 43—46‰, но выживает при этом лишь небольшая часть икринок. При понижении солености до 5‰ икра Clupea harengus harengus развивается вполне нормально, причем и в естественных условиях вода на нерестилищах может быть почти пресной. Нормально идет развитие икры и пересаженной из морской воды в пресную, но личинки в пресной воде погибают в течение суток. К числу наиболее евригалинных видов относятся виды рода Alosa. У Alosa caspia salina икринки в естественных условиях развиваются при содержании солей до 22.0—32.2‰, у Alosa caspia caspia — как в пресной, так и в солоноватой воде с концентрацией солей до 4—6 и даже 8.4‰. В условиях эксперимента икра этой сельди развивается при 12.5‰, и, вероятно, верхним пределом, при котором икринки гибнут, следует считать соленость 15‰. Таким же евригалинным видом является Clupeonella delicatula, икринки каспийской формы которой развиваются как в пресной, так и в солоноватой воде до 20‰, при которой они гибнут. Хотя проходные виды сельдей и размножаются в пресной воде, однако некоторые (например Alosa kessleri volgensis) иногда выметывают икру и в солоноватой воде в море, где развитие их протекает при солености 1‰, а личинок — при 9.5‰. В условиях опыта икра этой сельди может развиваться при 20—30‰. Как показали эксперимзнтальные исследования, развитие икринок другого проходного вида этого рода (Alosa sapidissima) наилучшим образом происходит при содержании солей около 7.5‰.
Икра некоторых сельдевых имеет свойство развиваться партеногенетически. Так, например, неоплодотворенная икра Alosa kessleri volgensis, помещенная в воду, проходит начальные стадии дробления, но при этом наблюдаются некоторые аномалии в дроблении, и процесс этот идет медленнее, чем у оплодотворенной икры. Неоплодотворенная икра развивается при 17—18° в течение 29 час. до стадии гаструляции, после чего до стадии глазных бокалов идет ее разрушение (Соин, 1941). Икра Alosa fallax lariana в слабых гипотонических растворах некоторых солей и под действием в течение короткого времени (20—30 сек.) слабого тока (2—3 вольта) развивается вполне нормально, возможно даже более быстро, чем оплодотворенная, и из нее выходят личинки, не отличимые от личинок, вышедших из оплодотворенной икры (Lestage, 1934).
На развивающуюся икру оказывают существенное влияние механические воздействия — удар, падение и встряхивание. Особенно гибельно действуют на икру в условиях опыта и в рыбоводных аппаратах удары и падение. Встряхивание, имеющее место и в естественных условиях, оказывает меньшее влияние. Наиболее чувствительна ко всем этим воздействиям у проходной сельди Alosa kessleri volgensis икра, развивающаяся при температуре около 19°, близкой к температуре развития ее в естественных условиях, в первые часы после оплодотворения на стадии 16—64 бластомеров, особенно на 2—3-м часу, и в дальнейшем на время перед выклевыванием личинок, на 35—50-м часу (Строганов, 1939). В связи с этим следует отметить, что первый чувствительный период развивающейся икры у проходных сельдей приходится на наиболее тихие ночные часы, так как икрометание у этих сельдей происходит в вечернее время при закате солнца и непосредственно перед и после него.
Многие из сельдевых совершают миграции, протяженность которых у некоторых значительна. Ряд сельдевых ведет более или менее локальный образ жизни в пределах своего, иногда ограниченного ареала. Размеры миграций бывают различны иногда даже у форм одного и того же вида и подвида. Так, одни подвиды Alosa caspia (Alosa caspia knipowitschi, Alosa caspia persica) почти не выходят за пределы южной части Каспийского моря, другие (Alosa caspia caspia, Alosa caspia salina) совершают миграции значительной протяженности с мест зимовок в южной части моря в северную для икрометания. В южной части Каспийского моря, почти не выходя за пределы его, обитает и большинство подвидов Alosa brashnikovi (Alosa brashnikovi orientalis, Alosa brashnikovi kisselevitshi и др., всего 6 подвидов), другие (Alosa brashnikovi brashnikovi, Alosa brashnikovi agrachanica) идут для размножения весной из Южной части моря в северную. Точно так же одни подвиды Clupea harengus (Clupea harengus pallasi, Clupea harengus pallasi n. suworowi, Clupea harengus pallasi n. maris-albi) ведут более, другие (Clupea harengus harengus) менее локальный образ жизни. Clupea harengus harengus может служить примером разной степени миграций и у разных форм подвида. Наибольшие по протяженности миграции из всех форм Clupea harengus harengus совершают атлантическо-скандинавские сельди, в частности норвежская и мурманская сельди; более локальны сельди, например, датских проливов.
Большая протяженность миграций свойственна, как указывалось (см. «Система семейства и принципы классификации»), видам и формам, наиболее прогрессивным по комплексу морфологических особенностей, положенных в основу системы.
Миграции сельдевых, как и других рыб, в основном связаны с размножением или питанием. Примеры миграций первого рода приведены выше. В качестве миграций второго рода следует указать на миграции дальневосточной сардины (Sardinops sagax melanosticta), которая после зимовки и икрометания в южной части Японского моря и у южных островов Японии с тихоокеанской стороны в начале лета идет на север до самой северной части Татарского прол. и в Тихом океане в годы потепления до восточного побережья Камчатки. Такие же обширные миграции совершает и калифорнийская сардина (Sardinops sagax caerulea). Эти миграции относятся к числу самых больших по своей протяженности.
Миграции, связанные как с размножением, так и с питанием, во всех приведенных примерах, а также и у проходных каспийских сельдей иа рода Alosa имеют меридиональное направление, с юга на север. Эти миграции имеют одну, общую почти всем им особенность: продвигаясь на север вдоль берегов на некотором расстоянии от них, обычно в области морского течения, эти сельди при наличии положительного градиента температуры воды подходят в прибрежную область, где и ловятся закидными и ставными неводами и другими орудиями лова.
Существуют миграции и более длительные по времени, продолжающиеся в течение нескольких лет; они начинаются уже в личиночной, стадии, и совершают их некоторые формы Clupea harengus harengus. Личинки мурманской сельди по выходе из икры, откладываемой в некотором отдалении от берегов, попадают в сферу действия Североатлантического течения и уносятся вдоль берегов северной Норвегии и Финмаркена на север и восток. В дальнейшем мальки и подрастающая сельдь уже активно расселяются в центральных и восточных частях Баренцова моря, заходят частично в Белое море. После первого икрометания мурманская сельдь отходит к северу, к Шпицбергену. Приблизительно таков же характер миграций и у норвежской сельди (подробнее см. в систематической части). Повидимому, подобные миграции совершают и некоторые формы Clupea harengus pallasi, но меньшей протяженности, и охватывают лишь часть ее жизненного цикла, так как личинки этих сельдей по выходе из икры, откладываемой на небольшой глубине у самого берега, здесь, же на местах икрометания и остаются до превращения в мальков. Лишь после этого, отходя от берегов и попадая в область морского течения, мальки расселяются в сфере действия этого течения, но, по понятным причинам, на меньшие расстояния от мест икрометания. Такие миграции, судя по всему, совершает южносахалинско-хоккайдская сельдь. Большинство остальных форм Clupea harengus pallasi проводит жизнь вблизи мест икрометания, занимая часто очень ограниченный ареал (подробнее см.: Световидов, 1949 б). Следует отметить, что миграции сельдевые, как и другие рыбы, совершают в области морских течений, проходящих вблизи мест нереста; в сфере влияния этих течений и совершаются миграции. При этом миграции совершаются как против морского течения, так и по течению. Так, например, Alosa caspia caspia и Alosa kessleri kessleri идут в северную, часть Каспийского моря в основном вдоль западных берегов против течения, Alosa brashnikovi brashnikovi и Alosa kessleri volgensis — как вдоль западных, так и вдоль восточных, против течения.
Миграции сельдевых, а также в первую очередь характер питания обусловливают весьма важное в хозяйственном отношении различие в численности отдельных видов и форм сельдевых. Этот вопрос рассмотрен более подробно в статье автора (О некоторых факторах, обусловливающих численность сельдевых), публикуемой в Трудах конференции по вопросам рыбного хозяйства. Как известно, одни из видов и форм сельдевых отличаются большей численностью и являются важными в промысловом отношении, численность других не велика, и эти виды и формы не имеют промыслового значения или оно незначительно. К числу видов, отличающихся большой численностью и высокими уловами, относятся тихоокеанская сардина (Sardinops sagax), океаническая сельдь (Clupea harengus), сельдь, известная под названием менхэден (Brevoortia tyrannus), атлантическая сардина (Sardina pilchardus) и некоторые другие (см. «Хозяйственное значение»). Все эти виды по характеру питания являются планктоноядными. Ни одна из хищных сельдей не является по численности и уловам рыбой первостепенного промыслового значения. Равным образом и среди, например, каспийских сельдей наибольшей численностью, а в связи с этим и наибольшими уловами, отличается планктоноядный каспийский пузанок (Alosa caspia). Хищные, питающиеся в основном рыбой, бражниковские сельди (Alosa brashnikovi), несмотря на большое количество форм, имеют значительно меньший промысловое значение. Точно так же из форм Alosa kessleri в значительно большем количестве ловится в основном планктоноядная волжская сельдь (Alosa kessleri volgensis), чем черноспинка (Alosa kessleri kessleri), питающаяся главным образом рыбой. Таким образом, численность и промысловое значение того или иного вида и формы сельдевых обусловливается в первую очередь характером питания, планктоноядным или хищным. Хищные сельди питаются в основном рыбой, которая, в свою очередь, питается планктоном. Планктоноядные же рыбы минуют это промежуточное звено в пищевой цепи.
Далее, среди планктоноядных сельдей обращает на себя внимание то, что одни виды отличаются большей численностью, другие меньшей. Это различие в численности планктоноядных сельдей обусловлено отчасти размерами ареала и нерестилищ, а в основном протяженностью миграций. Наибольшую численность среди сельдевых имеют те виды, ареал которых наиболее широк, что вполне понятно. На первом месте по уловам среди сельдевых и вообще в уловах мирового промысла стоит тихоокеанская сардина (Sardinops sagax), занимающая обширный ареал по обе стороны северной части Тихого океана и в южном полушарии у берегов Южн. Америки и Австралии и у берегов Африки в Индийском океане. Однако в южной части Тихого океана промысел тихоокеанской сардины не развит, а основная масса ее добывается в северной его части, т. е. на ареале, значительно меньшем ареала океанической сельди (Clupea harengus), широко распространенной в северных частях Атлантического и Тихого океанов, но вылавливаемой в меньшем количестве. Таким образом, численность обусловливается не одними размерами ареала, а также в значительной мере, как указывалось, и протяженностью миграций. Sardinops sagax совершает наибольшие по своему размаху миграции. Находясь в зимнее время в южной части ареала, Sardinops sagax melanosticta, повидимому, питается и в это время, устремляясь затем весной на север, где также усиленно откармливается после икрометания. Таким образом, Sardinops sagax melanosticta наиболее полно использует свой ареал для нагула. Из форм Clupea harengus harengus, отличающихся большой численностью, следует отметить упомянутых выше норвежскую и мурманскую сельдь, которые, занимая значительный ареал, совершают в пределах его, как указывалось, миграции большой протяженности. Наибольшей численностью из всех форм Clupea harengus pallasi отличается южносахалинско-хоккайдская сельдь, которая, судя по всему, также, как уже отмечалось, на протяжении жизненного цикла совершает миграции. Большинство форм Clupea harengus pallasi, если не все остальные, ведет локальный образ жизни в пределах ограниченного ареала, и численность их невелика. Из каспийских подвидов Alosa caspia наибольшей численностью отличается Alosa caspia caspia, совершающий, как указывалось, миграции в северную часть моря, где после икрометания остается в течение некоторого времени и для нагула. Не совершающие таких миграций южно-каспийские подвиды (Alosa caspia knipowitschi, Alosa caspia persica) по численности значительно уступают Alosa caspia caspia. Равным образом среди подвидов Alosa brashnikovi наибольшей численностью отличается Alosa brashnikovi brashnikovi, совершающий, подобно Alosa caspia caspia, миграции в северную часть моря; уловы южнокаспийских подвидов Alosa brashnikovi значительно ниже уловов Alosa brashnikovi brashnikovi.
Наибольшая численность свойственна, как указывалось (см. «Система семейства и принципы классификации»), видам и формам, наиболее прогрессивным по комплексу морфологических особенностей, положенных в основу системы.
ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ
Цифры уловов как в этой главе, так и в «Систематической части» взяты из книги «Промысловые рыбы СССР» и у Расса (1948).
Сельдевые имеют наиболее важное значение в промысле, занимая по величине улова первое место среди рыб. В 1936—1937 гг. общий улов сельдевых составлял 63.4 млн ц, или 37.3% мирового улова рыбы.
Наибольшее значение в уловах сельдевых, а вместе с тем и в общих мировых уловах имела тихоокеанская сардина (Sardinops sagax), занимавшая по величине улова первое место среди промысловых рыб. Уловы тихоокеанской сардины, вернее двух ее подвидов северной части Тихого океана, в 1936—1939 гг. колебались от 27.3 до 34.8 млн ц, составляя около 43—55% мировой добычи сельдевых. Из этого количества у берегов юго-восточной Азии (дальневосточная сардина — Sardinops sagax melanosticta) добывалось 21.7—27.9 млн ц, у берегов Сев. Америки (калифорнийская сардина — Sardinops sagax caerulea) — 5.5—7.3 млн ц.
Второе место по величине улова среди сельдевых и вообще среди промысловых рыб занимает атлантическая сельдь (Clupea harengus harengus), которой в 1936 г. было добыто 16 млн ц, что составляет около 25 % общего улова сельдевых. У берегов Европы из этого улова поймано 15.2 млн ц, у берегов Сев. Америки — 0.8 млн ц.
Третье место по уловам среди сельдевых и четвертое в уловах мирового промысла занимает тихоокеанская сельдь (Clupea harengus pallasi). Уловы тихоокеанской сельди в 1936—1939 гг. колебались от 4 до 6.4 млн ц, из которых 1.4—1.8 млн ц поймано у берегов Сев. Америки. В среднем уловы тихоокеанской сельди составляют около 12% общего улова сельдевых.
Существенное значение в уловах сельдевых имеет сельдь, ловящаяся у атлантических берегов Сев. Америки и известная под названием менхэден (Brevoortia tyrannus). Уловы этой сельди в 1936 г. достигали 2.65 млн ц, или около 4.2% улова всех сельдей.
Почти такие же уловы дает и атлантическая сардина (Sardina pilchardus), которой в 1936 г. добыто свыше 2.5 млн ц, т. е. около 3.9% общего улова всех сельдей. Из этого количества в Атлантическом океане (Sardina pilchardus pilchardus) вылавливалось около 2.26 млн ц (1936 г.), в Средиземном море (Sardina pilchardus sardina) — 0.32—0.35 млн ц.
Весьма существенное значение в уловах мирового промысла сельдевых имеет салака (Clupea harengus harengus n. membras), которой добывалось около 0.79 млн ц, что составляет около 1.2% улова всех сельдей.
Из сельдевых, имеющих меньшее значение в уловах мирового промысла, следует отметить шпрота, или кильку (Sprattus sprattus), уловы которого не превышают 0.51 млн ц, т. е. 0.9% общего улова сельдей.
Уловы преобладающего большинства сельдевых, вылавливаемых как мировым, так и нашим промыслом, кроме упомянутых выше, менее значительны и существенной роли в мировых уловах не имеют. Однако уловы некоторых из сельдей, добываемых только в наших водах, имеют довольно существенное значение и в мировых уловах.
Основная масса общего улова сельдевых, около 65% его, добывалась в Тихом океане и в прилегающих к нему морях, значительно меньшая часть (немного больше 30%) — в Атлантическом океане со смежными морями. Сравнительно небольшая часть сельдевых вылавливается в солоноватых и пресных водах.
Общий улов сельдевых в СССР в годы, предшествовавшие Великой Отечественной войне, достигал в среднем свыше 3.5 млн ц, что составляет около 6% мирового улова сельдевых и около 24% общего улова рыбы СССР.
В уловах сельдевых СССР, так же как и в мировых, на первом месте стояла дальневосточная сардина (Sardinops sagax melanosticta), которая, однако, в общих наших уловах занимала пятое место. Уловы тихоокеанской сардины в 1931—1940 гг. колебались от 959 до 1397 тыс. ц. С 1943 г. эта рыба, как известно, ловиться перестала.
Большое значение в уловах СССР имеет тюлька (Clupeonella delicatula), которая в наших общих уловах сельдей занимает второе место. Уловы азовской тюльки (Clupeonella delicatula delicatula) в 1936—1938 гг. достигли 749—832 тыс. ц; каспийской (Clupeonella delicatula caspia) вылавливалось значительно меньше, в 1931 г. 169 тыс. ц.
Весьма существенное значение в уловах сельдевых СССР имеет океаническая сельдь (Clupea harengus), общий улов которой в годы, предшествовавшие Великой Отечественной войне, занимал в наших уловах одно из первых мест, уступая первенство дальневосточой сардине лишь в некоторые годы. В наибольшем количестве из подвидов этой сельди вылавливается тихоокеанская сельдь (Clupea harengus pallasi), уловы которой в 1936—1940 гг. составляли от 425 до 523 тыс. ц, занимая по величине улова третье место. Уловы остальных подвидов океанической сельди несколько менее значительны. Мурманской сельди (Clupea harengus harengus) в год наибольшего захода (1935) добыто 975 тыс. ц, в 1939 г. — 162 тыс. ц, в другие годы меньше. Балтийской сельди, или салаки (Clupea harengus harengus n. membras), в 1937 г. в пределах теперешних государственных границ поймано 230 тыс. ц. Уловы беломорской сельди (Clupea harengus pallasi n. maris-albi) в 1933—1946 гг. колебались от 13 до 36 тыс. ц. Чёшеко-печорской сельди (Clupea harengus pallasi n. suworowi) в 1937—1939 гг. вылавливалось 1.5—5.6 тыс. ц.
Каспийско-черноморские сельди, составлявшие когда-то основную массу добываемых в наших водах сельдей, в недавнее время по величине общего улова стояли на третьем месте после океанической сельди и тихоокеанской сардины.
На первом месте среди каспийско-черноморских сельдей по величине улова стоит каспийский пузанок (Alosa caspia caspia), который занимает видное (третье-четвертое место) и среди других форм сельдей. В 1936—1939 гг. уловы каспийского пузанка колебались от 56 (1936 г.) до 750 (1939 г.) тыс. ц. Другие формы каспийского пузанка ловятся в значительно меньшем количестве и обычно учитываются вместе с основной формой. Уловы черноморско-азовских форм этого вида незначительны.
Приблизительно в таком же количестве вылавливаются проходные сельди. За период времени с 1936—1939 гг. волжской сельди (Alosa kessleri volgensis) вылавливалось от 75 (1936 г.) до 691.7 (1939 г.) тыс. ц, черноспинки (Alosa kessleri kessleri) от 13.7 до 57.5 тыс. ц, азовско-черноморской сельди (Alosa kessleri pontica) от 52.3 до 76.9 тыс. ц.
Бражниковские сельди (Alosa brashnikovi) играют в уловах меньшую роль. Основная масса уловов этих сельдей приходится на долгинскую сельдь (Alosa brashnikovi brashnikovi), которой в 1936—1939 гг. вылавливалось от 73.9 до 126.9 тыс. ц. В небольшом количестве добывается аграханская сельдь (Alosa brashnikovi agrachanica); уловы ее в 1935—1940 гг. составляли от 3.2 до 16.1 тыс. ц. Гасанкулинская сельдь (Alosa brashnikovi kisselevitshi) за период времени с 1934 по 1938 гг. вылавливалась в количестве от 3 до 29.5 тыс. ц. Остальные формы этого вида вылавливаются еще в меньшем количестве.
Большеглазый пузанок (Alosa saposhnikovi) имеет еще меньшее значение, чем бражниковские сельди; в 1936—1939 гг. уловы его колебались от 38 до 88 тыс. ц.
Шпрот, или килька (Sprattus sprattus), ловится в наших водах в настоящее время в незначительном количестве, так как вылавливается лишь часть имеющихся запасов, в частности в Черном море; в Балтийском море в границах современных вод СССР в 1936 г. добыто (Sprattus sprattus balticus) около 22 тыс. ц, в Черном море в 1937—1938 гг. (Sprattus sprattus phalericus) 13.2—34.7 тыс. ц.
Наибольшее значение в уловах сельдевых в СССР имеют дальневосточные моря, где в годы, предшествовавшие Великой Отечественной войне, вылавливалось до 45% всего количества добываемой нашим промыслом сельди, или около 12—13% общего улова рыбы в Беринговом и Охотском морях и свыше 80% улова в Японском. В связи с падением промысла сардины уловы сельди в настоящее время имеют здесь меньший удельный вес, однако они могут быть значительно увеличены, так как запасы тихоокеанской сельди недоиспользуются. На втором месте по уловам сельдей стоит Каспийское море, дававшее в те же годы почти 30% нашего улова сельдевых, или около 25% общего улова рыбы этого бассейна. Уловы сельдей в Каспийском море могут быть значительно увеличены за счет лова тюльки (так называемой каспийской кильки). Третье место по уловам сельди занимают Черное и Азовское моря, где добывается свыше 20% нашего общего улова сельдевых, или почти 30% общего улова рыбы в Азовском море и около 15% в Черном. Уловы здесь могут быть также увеличены за счет организации промысла шпрота. Остальные морские промысловые районы имеют меньшее значение в наших уловах сельдевых. Баренцово море, имеющее значительные запасы сельди, в настоящее время по уловам ее не играет существенной роли в общих уловах сельдевых. В годы, предшествовавшие Великой Отечественной войне, уловы сельди в Баренцовом море и Чёшской губе составляли всего лишь около 2% общего улова сельдевых или менее 3% общего улова здесь рыбы. В отдельные годы, как, например, в 1935 г., уловы сельди в Баренцовом море имели больший удельный вес в общих уловах сельдевых СССР. Уловы сельдевых в Балтийском море в общих уловах их составляют несколько более 1%, но могут быть значительно увеличены. В Белом море вылавливается несколько менее 1% общего улова сельдевых СССР, однако в общих уловах этого моря сельдь составляет свыше 20%.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Сем. CLUPEIDAE — СЕЛЬДЕВЫЕ
— Clupeidae Cuvier, Regne animal, ed. II, II, 1829 : 316 (частью).
— Clupeoidei Müller, Arch. Naturgesch., 9 Jahrg., 1 Bd., 1843 : 324 (частью).
— Clupeidae Müller, Abh. Akad. Wiss. Berlin (1844), 1846 : 136 (частью), Günther, Cat. fish., VII, 1868 : 381 (частью).
— Clupeidae + Dorosomidae Jordan a. Evermann, Pish. N. a. M. America, I, 1896 : 415, 417.
— Clupeidae + Dussumieriidae + Dorosomidae Jordan, Classif. fish., 1923 : 120, 121, 122.
— Clupeidae Берг, Система рыб, 1940 : 223.
— Dorosomidae + Stolephoridae + Clupeidae Fowler, U. S. Nat. Mus., Bull. 100, v. 13, 1941 : 547, 561, 578.
Боковая линия на теле отсутствует, развита лишь в самой передней части в виде короткого канала с порами на первых 2 (у Sardina) — 5 (у Clupea) чешуях сразу за головой. Каналы системы боковой линии на голове на черепных костях заключены в костных каналах, пронизанных местами наружу отверстиями, или (на подглазничных и отчасти на нижнечелюстных) расположены под гребнями костей; от этих каналов ответвляются многочисленные более мелкие каналы, расположенные в кожных покровах на многих костях черепа, жаберной крышки, челюстей (исключая interoperculum, praemaxillare и maxillare) и на чешуях сзади головы сверху и у некоторых (Alosa, Sardina и др.) достигающие сильного развития; такой короткий канал, не заключающий нервных органов, имеется в кожных покровах на верхней части operculum. Отверстопузырные. Плавательный пузырь посредством выростов, заключенных в prooticum и pteroticum (у Sprattus и Clupeonella pteroticum выростов плавательного пузыря не заключает) соединяется с ушными капсулами; задний конец плавательного пузыря продолжен в канал, открывающийся наружу слева ostium genitale между анальным и мочевым отверстиями или заканчивающийся слепо. Рот окаймлен посредством praemaxillaria и maxillaria или только посредством praemaxillaria (у большинства Dorosomatinae). В верхней челюсти 1 или 2 supramaxillaria. У некоторых (Pomolobus, Sardinops) есть praeethmoidalia. Соединительнотканная связка между задним концом praemaxillare и серединой maxillare у некоторых (Pellona, Pliosteostoma) превращена в кость, несущую зубы. Миодом открыт и сзади черепа. Есть верхние (epipleuralia) и нижние (epicentralia) межмышечные косточки и нижние и верхние ребра; дистальная часть последних хрящевая, проксимальная — соединительнотканного происхождения. Postcleithrum 3, прикрепляются с наружной стороны cleithrum. Мезокоракоид, базисфеноид и орбитосфеноид есть. Спинной плавник по середине тела, иногда (у Pristigasterinae) над анальным, у Raconda отсутствует. Брюшные плавники на брюхе, иногда отсутствуют (у некоторых Pristigasterinae). Брюхо закругленное или сжатое с боков, брюшные килевые чешуи есть или отсутствуют (у Dussumieriinae), чешуя циклоидная или у некоторых (Brevoortia, Ethmalosa и Ethmidium) на заднем крае гребенчатая. Как и у других современных семейств подотряда Clupeoidei, жирового плавника нет, парапофизы не приращены к телам позвонков, грудные radialia расположены в один ряд, praedentale отсутствует, есть нормальные яйцеводы. Из признаков, свойственных другим Clupeiformes, следует отметить гомоцеркальный хвостовой плавник, отсутствие фулькр и веберова аппарата, непарные сошник и мезэтмоид, есть hypuralia и эндохондральное supraoccipitale.
Из характерных особенностей эмбрионального развития необходимо указать на положение спинного плавника у личинок далеко позади тела, над анальным; этот плавник по мере роста личинок перемещается над миотомами и ко времени превращения личинки во вполне сформировавшегося малька занимает положение, свойственное взрослым. Икринки имеют малое или большое кругожелтковое (перивителлиновое) пространство, без жировой капли или с одной крупной или многочисленными мелкими, быстро исчезающими каплями.
Около 50 родов и до 190 видов в тропических, субтропических, умеренных и единично в арктических морях и в соответствующих им широтах в пресных водах обоих полушарий. В ископаемом состоянии с нижнего мела.
По некоторым особенностям внутри семейства можно провизорно выделить следующие подсемейства.
I. Dussumieriinae. Брюшных килевых чешуй нет, брюхо закругленное. Верхняя челюсть без медиальной вырезки. Supraorbital 1 (у Dussumieria, Etrumeus, Spratelloides). На хвостовом плавнике удлиненных чешуй (ala) нет. Жировые веки есть или отсутствуют (у Spratelloides), у Etrumeus и, повидимому, у Dussumieria целиком закрывают глаза. Внутренняя стенка fossa praeepiotica перепончатая, не отделена костной перегородкой, от fossa temporalis posterior (у Dussumieria, Etrumeus, в меньшей степени у Spratelloides).
Роды Dussumieria, Etrumeus, Jenkinsia (включая Halecula Jordan, 1925), Spratelloides (= Stolephorus auct., включая Gilchristella Fowler, 1935, и Sauvagella Bertin, 1940 = Spratellomorpha Angel, Bertin et Gruibe, 1946). Морские рыбы тропических и субтропических морей; Spratelloides malabaricus — в эстуариях и реках.
Обзор индо-пацифических родов см. у Фаулера (Fowler, 1941 : 561—578), рода Jenkinsia у Джордэна и Эвермана (Jordan a. Evermann, 1896 : 418—421), подсемейства у Бертена (Bertin, 1943), описание Dussumieria productissima у Шабано (Chabanaud, 1933 : 4).
II. Clupeinae. Брюшные килевые чешуи, как и у всех следующих, обычно хорошо развитые и явственно заметные или реже менее развитые и заметные. Брюхо сжатое с боков или реже закругленное. Верхняя челюсть без заметной медиальной вырезки. Сочленение нижней челюсти с черепом и задний конец верхнечелюстной кости впереди или под вертикалью середины глаза, у Sardinops немного позади него. Жировое веко есть, у некоторых (Clupeonella) зачаточное или почти не развитое. Удлиненных чешуй при основании лопастей хвостового плавника нет, реже (у Sardinella, Sardinops, Sardina) имеются. У Sardinella, Sardinops, Sardina и Clupeonella два последних луча анального плавника удлиненные. У Opisthonema последний луч спинного плавника вытянут в длинную нить.
Роды Clupea, Sprattus, Harengula, Clupeonella, Sardina, Sardinops, Sardinella, Opisthonema, Lile, Rhinosardinia (= Heringia). Heringia есть nomen praeoccupatum (Diptera). См.: Myers, 1929 : 1. Морские и отчасти солоноватоводные и пресноводные рыбы тропических, субтропических, умеренных и частично арктических морей. Harengula tawilis и Clupeonella abrau — в пресноводных озерах, Clupeonella delicatula — отчасти проходной и пресноводный, Rhinosardinia amazonica — в Амазонке и реках Гвианы.
Обзор рода Clupea, включая виды рода Sprattus, см. у Ригэна (Regan, 1916 : 1—6; 1917а : 226—229) и Нормэна (Norman, 1936 : 491—492; 1937 : 37—39), Harengula, Sardinella, Opisthonema, Lile, Rhinosardinia — у Ригэна (Regan, 1917c : 377—395), индо-пацифических видов Harengula и Sardinella — у Фаулера (Fowler, 1941 : 584—620), ост-индских Harengula у Сторей (Storey, 1938 : 1—56), Sardina, включая Sardinops — у Ригэна (Regan, 1916 : 11—19).
III. Pellonulinae. Брюхо сжатое с боков, реже (у Stolothrissa и Odaxothrissa) впереди брюшных плавников закругленное, со слабым килем. Верхняя челюсть без медиальной вырезки. Supramaxillare только 1. Сочленение нижней челюсти с черепом под вертикалью середины глаза или впереди него, задний конец верхнечелюстной кости слегка простирается за вертикаль переднего края глаза или не достигает ее. Жирового века нет или у некоторых едва развито. У Corica и Clupeichthys позади анального плавника 2 плавничка. У Potamalosa и Hyperolophus между затылком и спинным плавником ряд килевидных щитков.
Роды Pellonula, Poecilothrissa, Microthrissa, Potamothrissa, Cynothrissa, Odaxothrissa, Stolothrissa, Limnothrissa, Kowala, Clupeoides, Corica, Clupeichtys, Potamalosa, Hyperlophus. Пресноводные тропические или реже субтропические рыбы Африки (первые 8 родов), Индо-малайского архипелага и отчасти Индии и Австралии. Kowala thoracata — в море вдоль берегов западной Индии, Явы и других островов; оба вида Hyperolophus, повидимому, также морские.
Обзор родов см. у Ригэна (Regan, 1917 : 198—207; 1922 : 587—590), последних шести родов — также у Фаулера (Fowler, 1941 : 635—648), который род Clupeoides считает идентичным роду Kowala, так же как род Clupeichthys роду Corica.
IV. Alosinae. Верхняя челюсть с заметной медиальной вырезкой. Сочленение нижней челюсти с черепом позади или реже (у Pomolobus) впереди вертикали заднего края глаза, задний конец верхнечелюстной кости простирается за вертикаль середины глаза или заднего края его. Есть жировое веко. При основании хвостового плавника на каждой лопасти по удлиненной чешуе (ala), отсутствующей у Hilsa и Gudusia. У Brevoortia, Ethmalosa и Ethmidium чешуя на заднем крае гребенчатая и между затылком и основанием спинного плавника имеется по 2 ряда увеличенных чешуй (у Brevoortia и Ethmalosa) или щитков (у Ethmidium).
Роды Alosa, Pomolobus, Brevoortia, Ethmidium, Ethmalosa, Hilsa (= Macrura), Gudusia. Проходные, некоторые пресноводные (Gudusia, озерные и речные формы видов Alosa s. str. и некоторых видов Pomolobus) или отчасти солоноватоводные (Brevoortia, некоторые каспийские виды и формы Alosa; биология Ethmidium и Ethmalosa не известна) рыбы умеренных, тропических и менее субтропических широт.
Обзор родов см. у Ригэна (Regan, 1917b : 297—308), родов Hilsa и Gudusia также у Фаулера (Fowler, 1941 : 626—635). Биологию видов. Pomolobus и Brevoortia см. у Бигелоу и Уелша (Bigelow a. Welsh, 1925 : 105—124), описание Brevoortia smithi у Гильдебрандта (Hildebrandt, 1941 : 224).
V. Dorosomatinae. Верхняя челюсть с медиальной вырезкой. Supramaxillare 1, реже 2 (у Signalosa). Рот без зубов, нижний или конечный, окаймлен посредством только praemaxillaria или посредством как praemaxillaria, так и maxillaria (у Clupanodon). Последний луч спинного плавника у всех, исключая Gonialosa и Anodontostoma, вытянут в длинную нить. Есть жировое веко. По внешнему, виду большинство, исключая Clupanodon, высоким и сжатым с боков телом, небольшим нижним ртом и длинным анальным плавником (у Dorosoma имеющем до 38 лучей) напоминают леща.
Роды Dorosoma, Clupanodon, Signalosa, Nematalosa, Gonialosa, Anodontostoma. Проходные или пресноводные и отчасти морские рыбы заливов, и эстуариев тропических или отчасти субтропических широт.
Обзор родов см. у Ригэна (Regan, 1917b : 308—316), четырех последних родов также у Фаулера (Fowler, 1941 : 547—561), Signalosa — у Уида (Weed, 1925 : 137—146), описание Dorosoma smithi — у Хэббса и Миллера (Hubbs a. Miller, 1941 : 232—238).
VI. Pristigasterinae. Анальный плавник длинный, более чем с 30 (33— 92) лучами. Верхняя челюсть без медиальной вырезки. Брюшные килевые чешуи развиты и на горле. Брюшные плавники малы или отсутствуют (у Pristigaster, Opisthopterus, Odontognathus, Raconda). Спинной плавник расположен за брюшными, впереди анального, над или позади начала его, у Raconda отсутствует. Соединительнотканная связка между задним концом praemaxillare и серединой maxillare у некоторых (Pellona, Pliosteostoma) превращена в кость, несущую зубы.
Роды Ilisha, Pellona (= Neosteus), Chirocentrodon (Включая Pseudochirocentrodon Miranda-Ribeiro (1920), описание которого дано в недоступном автору издании. Отличается положением спинного плавника.) (= Мedipellona), Pristigaster, Pliosteostoma, Opisthopterus, Odontognathus, Raconda. Морские рыбы тропических морей, немногие виды некоторых родов встречаются в пресных водах. Ilisha indica — входит в реки, Ilisha macrogaster — в реках, Ilisha xanthoptera — в солоноватой воде и в реках, Ilisha elongata, Ilisha africana, Ilisha pristigasteroides, Ilisha megaloptera — заходят в устья рек, Opisthopterus macrognathus — в море и эстуариях, Pellona castelnaeana — в море и в реках, Pellona altamazonica — в реках.
Обзор родов см. у Нормэна (Norman, 1923 : 1—22), индо-пацифических видов Pellona, Ilisha, Opisthopterus, Raconda — у Фаулера (Fowler, 1941 : 648—666), рода Chirocentrodon — у Бредера (Breder, 1942 : 133—138).
Некоторые из этих групп по характеру признаков, положенных в основу их разграничения, следует рассматривать скорее как трибы, чем как подсемейства. При более детальном изучении морфологии, в частности остеологии, всех представителей семейства, несомненно, будут обнаружены и другие, более существенные особенности, на основании которых окажется возможным внести изменения и дополнения и дать более обоснованное разделение на подсемейства. Наиболее примитивным по комплексу признаков, положенных в основу системы семейства (см. «(Система семейства»), в частности по положению спинного и анального плавников и размерам последнего, следует считать подсем. Pristigasterinae, наиболее высокоорганизованным — Dussumieriinae.
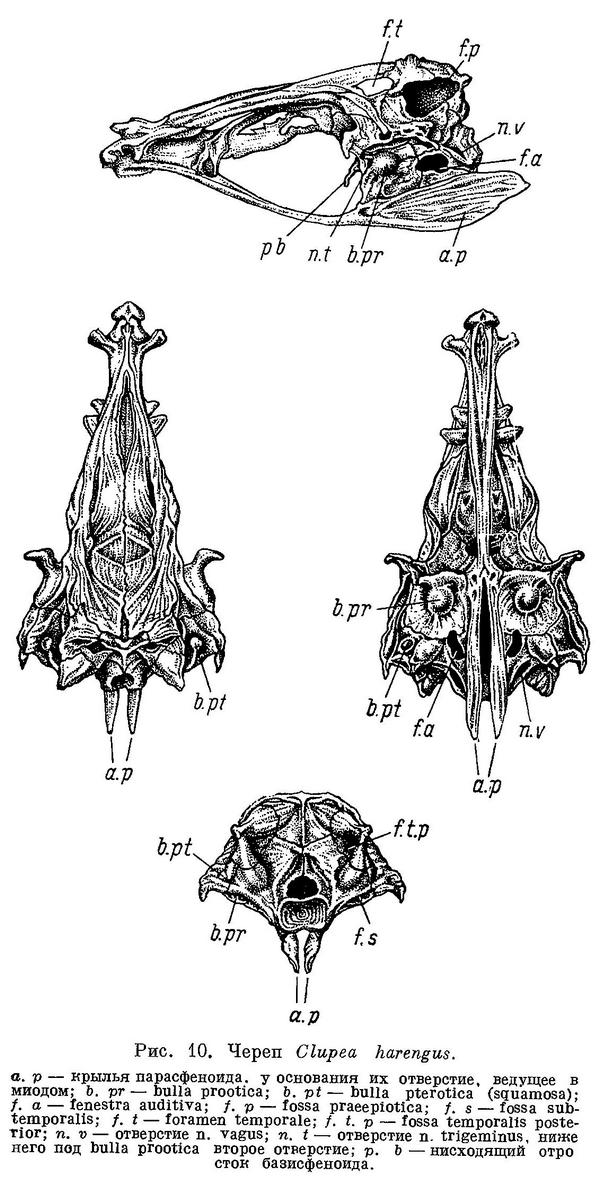
Следует отметить некоторые особенности строения черепа сельдевых (рис. 10). Fenestra auditiva (слуховое отверстие) располагается с каждой стороны сбоку черепа; спереди она ограничена посредством prooticum, снизу — basioccipitale, сверху — occipitale laterale и сзади — обоими последними костями; она закрыта перепонкой. Bulla prootica — шаровидное вздутие на prooticum, в котором помещены отростки переднего конца плавательного пузыря; оно снаружи на черепе слабо развито у Sardinops и еще менее у Sardinella; более всего развито у Sprattus и Clupeonella. Bulla pterotica (squamosa) — такое же, но обычно меньшее по размерам вздутие на pteroticum, заметное у одних (Clupea, Sardina) сбоку и сверху на этой кости, у других (Sardinops, Sardinella) только сверху, у третьих (например у Sprattus и Clupeonella) отсутствующее не только снаружи, но и внутри (рис. 11). Полости bulla prootica и bulla pterotica каждой стороны посредством узких костных каналов соединяются в верхней части ocdipitalia lateralia несколько выше отверстия, через которое каждый отросток плавательного пузыря входит в череп. Это отверстие расположено впереди отверстия n. vagus и отделено от последнего гребнем, идущим от основания occipitale laterale и отчасти прикрывающим сверху продольное углубление в этой кости, в котором расположен каждый отросток плавательного пузыря. У некоторых (Clupea) это углубление целиком закрыто гребнем, образующим на occipitale laterale с каждой стороны черепа продолговатое вздутие; отверстия для отростков плавательного пузыря расположены снизу этой кости. Относительные размеры bullae prootica и pterotica у разных родов семейства различны — у одних крупнее bulla prootica, у других bulla pterotica. Соотношение размеров их отчетливо видно на рентгеноснимках, по которым оно и установлено. Foramen temporale (височное отверстие) расположено с каждой стороны в верхней части черепа; оно ограничено снизу и сверху в передней части лобной и теменной костями и в задней части теменной. Fossa praeepiotica — большая ямка непосредственно позади foramen temporale с каждой стороны в верхней части сбоку черепа; образована спереди, сверху и сзади посредством теменной и epioticum и снизу посредством pteroticum. Fossa temporalis posterior — меньшая по размерам ямка с каждой стороны сзади черепа, имеющаяся у Etrumeus, Sardinella, Sardinops, Sardina и Clupanodon, почти не развитая у Sprattus, Clupea, Harengula и Clupeonella и отсутствующая у Alosa, Pomolobus, Brevoortia и др.; образована сверху посредством epioticum и снизу посредством occipitale laterale. Fossa subtemporalis (lateralis) — еще меньшая по размерам ямка с каждой стороны черепа сзади над отростком opisthoticum между этой костью и pteroticum; она есть сравнительно у немногих (Clupea, Sardinella, Sardinops, Alosa и некоторых других). Prooticum у переднего края bulla prootica сверху пронизано отверстием, n. trigeminus; такое же отверстие, обычно меньшее по размерам, имеется у переднего нижнего края или снизу bulla prootica. В occipitale laterale с каждой стороны сбоку черепа располагается по большому отверстию n. vagus. Каждое ethmoidale laterale пронизано отверстием n. olfactorius. По такому же отверстию имеется в basisphenoideum с каждой стороны от нисходящего отростка этой кости. Задняя часть парасфеноида вилкообразно раздваивается, образуя крылья парасфеноида, расширенные в вертикальной плоскости и образующие боковые стенки миодома; у основания каждого крыла парасфеноида расположено по небольшому отверстию, ведущему в миодом. Над сочленовной поверхностью hyomandibulare с каждой стороны черепа расположено по одному отверстию в лобной кости и по 3 в pteroticum, которые ведут в каналы системы боковой линии, соединяющиеся впереди в надглазничный канал. Этот канал, идущий вдоль наружного края в каждой из лобных костей, имеет в передней половине сверху по отверстию; впереди каждый канал из лобных костей выходит над ethmoidalia lateralia. Между передними концами лобных костей и мезэтмоидом обычно есть большая или меньшая по размерам фонтанель. Окологлазничное окостенение состоит из 7—8 костей: 2, иногда 1 (у Dussumieriinae) supraorbitalia, 1 praeorbitale (lacrimale), 4 infraorbitalia и небольшой dermosphenoticum; сверху supraorbitalia расположена небольшая nasale.
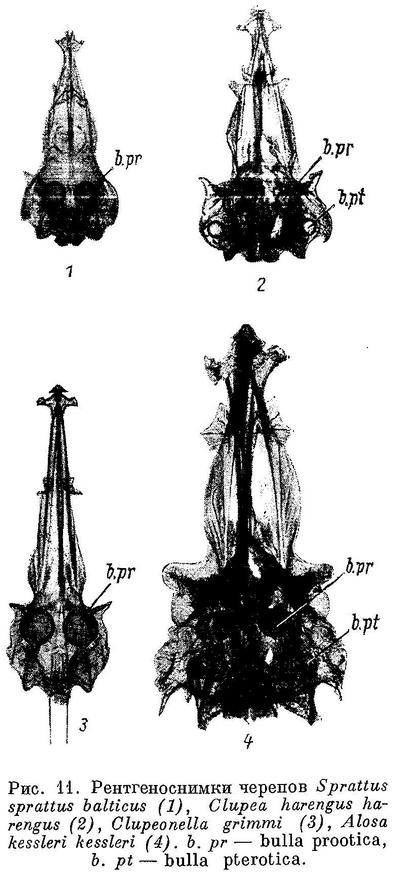
Измерение сельдей производилось по схеме (рис. 12), предложенной Бергом (1913 : 8—9): абсолютная длина тела (а—b) — расстояние от конца рыла (т. е. во всех случаях от переднего конца нижней челюсти при закрытом рте) до перпендикуляра, восстановленного от конца нижней лопасти хвостового плавника; длина тела по Смитту (а—с) — расстояние от конца рыла до конца средних лучей хвостового плавника; антеанальное расстояние (a—d) — расстояние от конца рыла до начала основания анального плавника; антедорсальное расстояние (а—е) — расстояние от конца рыла до начала основания спинного плавника; антевентральное расстояние (а—g) — расстояние от конца рыла до основания брюшного плавника; длина грудного плавника (f—h); длина брюшного плавника (g—i); высота тела наибольшая (е—k); высота тела наименьшая (l—m); расстояние между концом анального и началом хвостового плавников (n—o) (Оно характеризует длину хвостового стебля, который обычным способом, как у других рыб и как это рекомендует в своем «Руководстве к изучению рыб» И. Ф. Правдин, у сельдей измерить невозможно.); длина головы (а—р) — расстояние от конца рыла до заднего края жаберной крышки (обычно совпадает с местом соединения operculum с interoperculum, перепонка в расчет не принимается); длина рыла (а—r) — расстояние от вершины нижней челюсти при закрытом рте до переднего края глаза; диаметр глаза (r—s) — диаметр глазного яблока (продольный равен вертикальному; у рыб с жировым веком последнее надрезалось и частично удалялось); заглазничный отдел головы (s—р) — расстояние от заднего края глаза до заднего края жаберной крышки (как при измерении головы); высота головы (t—u) — расстояние по вертикали от верхнего края supraoccipitale; ширина межглазничного промежутка — ширина frontalia над серединой зрачка в том месте, где располагаются задние концы supraoccipitalia (кожные покровы в расчет не принимаются).
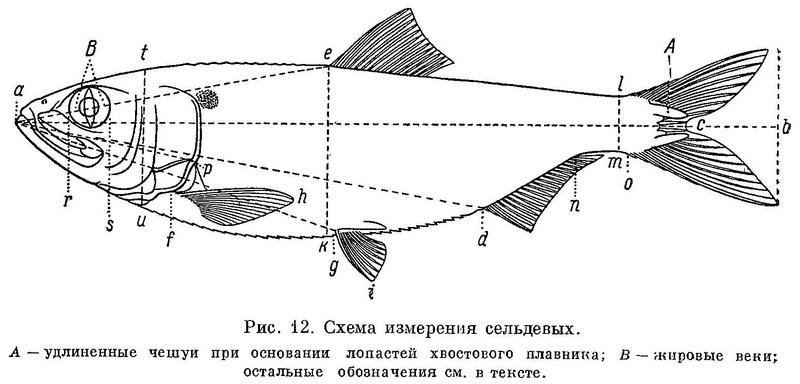
Измерения частей головы выражены в процентах ее длины, измерения частей тела — в процентах абсолютной длины тела, кроме сельдей из родов Clupeonella (Так как род Clupeonella относится к подсем. Clupeinae, у всех родов которого длину тела прицято измерять до конца хвостового плавника, то для этого рода в подстрочных примечаниях приведены некоторые признаки, выраженные в процентах и к последней длине.) и Alosa, у которых измерения принято выражать в процентах длины тела до конца средних лучей хвостового плавника. Расчисление темпа роста сельдей ведется по длине тела до конца средних лучей хвостового плавника, в тех же случаях, когда говорится о размерах рыбы, обычно принято употреблять абсолютную длину тела. К сожалению, этого правила придерживаются не все исследователи, и поэтому в тех случаях, когда приводится длина тела, измеренная каким-либо иным образом, каждый раз делается специальная оговорка. В частности, у сельдей из родов Clupeonella и черноморских и каспийских видов Alosa длина тела в последнее время преобладающим большинством исследователей измеряется только до конца средних лучей хвостового плавника. Поэтому для обоих этих родов, когда говорится о длине тела, то имеется в виду длина тела до конца средних лучей хвостового плавника. При просчете жаберных тычинок принимались во внимание и зачаточные тычинки на концах жаберных дуг. Количество позвонков, если это особо не оговорено, всюду приводится вместе с уростилем. В некоторых случаях к просчетам позвонков другими авторами, не принимавшими во внимание уростиль, прибавлялся один позвонок. Чешуи не просчитывались вследствие того, что они на теле сельдевых сохраняются плохо, а просчет их по кармашкам влечет к ошибкам.
Ввиду различий в методах измерения сельдей, применявшихся разными исследователями, причем измерения зачастую производились по рыбам, не всегда хорошо законсервированным, иногда даже по засоленным, из результатов исследований сельдей другими авторами здесь приводятся лишь счетные признаки.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
(В определительной таблице характеристика подсемейств касается лишь родов встречающихся в фауне СССР.)
1 (2). Брюшных килевых чешуй нет. Брюхо закругленное. Брюшные плавники позади заднего края основания спинного плавника. Жировое веко полностью закрывает глаз. (Подсем. Dussumieriinae). 1. Etrumeus
2 (1). Брюшные килевые чешуи есть, обычно хорошо развитые, реже слабые и менее заметные. Брюхо сжатое с боков и заостренное в виде киля или закругленное. Брюшные плавники под спинным или впереди него.
3 (20). Брюшных килевых чешуй на горле нет. Анальный плавник менее чем с 30 лучами, начало его далеко позади заднего конца спинного, брюшные плавники позади начала спинного или реже немного впереди.
4 (19). Последний луч спинного плавника не удлинен. Supramaxillaria 2.
5 (18). Верхняя челюсть без заметной медиальной вырезки. Сочленение нижней челюсти с черепом впереди вертикали заднего края глаза или под ней. (Подсем. Clupeinae).
6 (11). Последние два луча анального плавника не удлинены в виде лопасти. Удлиненных чешуй при основании хвостового плавника на лопастях его нет.
7 (10). На вертикальной части ключицы двухлопастного кожистого выроста нет.
8 (9). Брюшные килевые чешуи хорошо развитые. Брюхо сжатое с боков, с явственным килем на всем протяжении от горла до анального плавника. Брюшные плавники впереди или под началом спинного. Небольшие рыбы длиной до 18 см. 2. Sprattus
9 (8). Брюшные килевые чешуи слабые. Брюхо впереди брюшных плавников закруглено, позади несколько сжатое с боков, со слабым килем. Брюшные плавники позади начала спинного. Крупные рыбы, в половозрелом состоянии от 10—20 см и более. 3. Clupea
10 (7). На вертикальной части ключицы двухлопастный кожистый вырост. Последние два луча анального плавника слегка удлинены, но не образуют выдающейся свободной лопасти. 4. Harengula
11 (6). Последние два луча анального плавника удлинены в виде лопасти.
12 (17). Удлиненные чешуи (ala) при основании лопастей хвостового плавника есть.
13 (14). На вертикальной части ключицы двухлопастный кожистый вырост. На крышечной кости радиальных бороздок нет. 5. Sardinella
14 (13). На вертикальной части ключицы двухлопастного кожистого выроста нет. На крышечной кости радиальные бороздки.
15 (16). Жаберные тычинки нижней части жаберной дуги укороченные в углу дуги, на них налегают сверху тычинки верхней части дуги. Рот большой, сочленение нижней челюсти с черепом позади вертикали середины глаза, задний конец верхнечелюстной кости достигает или почти достигает середины глаза, у крупных простираясь и далее. Чешуи одинакового размера. 6. Sardinops
16 (15). Жаберные тычинки нижней части жаберной дуги в углу дуги не укороченные, одной длины с тычинками верхней части дуги, не налегающими на них сверху. Рот небольшой, сочленение нижней челюсти с черепом впереди вертикали середины глаза, задний конец верхнечелюстной кости не достигает середины глаза. Чешуи на боках тела неодинакового размера — крупные и скрытые под ними мелкие. 7. Sardina
17 (12). Удлиненных чешуй (ala) при основании лопастей хвостового плавника нет. Двухлопастного кожистого выроста на вертикальной части ключицы и радиальных бороздок на крышечной кости нет. 8. Clupeonella
18 (5). На верхней челюсти есть медиальная вырезка. Сочленение нижней челюсти с черепом за вертикалью заднего края глаза. При основании лопастей хвостового плавника есть удлиненные чешуи (ala). (Подсем. Alosinae). 9. Alosa
19 (4). Последний луч спинного плавника вытянут в длинную нить. Supramaxillare 1. (Подсем. Dorosomatinae). 10. Clupanodon
20 (3). Брюшные килевые чешуи есть и на горле. Анальный плавник длинный, более чем с 30 лучами, начало его под спинным плавником или сразу за задним концом его. Брюшные плавники малы, расположены впереди начала спинного. (Подсем. Pristigasterinae). 11. Ilisha
Рыбы пресных вод СССР и сопредельных стран. Л.С.Берг 1948 г.
Семейство CLUPEIDAE. — Сельдевые
Regan, Ann. Mag. Nat. Hist. (8), XIX, 1917, p. 297. — Берг, 1. с., стр. 223.
Тело покрыто чешуей. Голова голая. Брюхо сжатое или закругленное; если сжатое, то нередко снабжено килеватыми чешуйками. Рот конечный, реже нижний, челюсти почти одинаковой длины. Рыло не выдающееся. Край верхней челюсти сбоку образован верхнечелюстными костями, а посреди — межчелюстными; последние не выдвижные. На верхнечелюстной кости есть добавочная кость. Верхнечелюстная кость умеренной длины. Зубы небольшие, иногда отсутствуют. Жаберные перепонки свободны. Жаберные тычинки длинные, тонкие. Горловой пластинки нет. Жирового плавника нет. Боковой линии нет. (Точнее, в боковой линии всего от 2 (Sardina) до 5 (Clupea) чешуй.) Кости жаберной крышки нормальные (в числе 4). Спинной плавник по середине тела или несколько позади (но не над анальным), обычно есть. Анальный плавник удлиненный или умеренной длины. Хвостовой сильно выемчатый. Брюшные плавники умеренной величины или маленькие, иногда отсутствуют. Усиков нет. Плавательный пузырь есть. Postcleithrum на внешней стороне ключицы.
Во всех морях и океанах; многие входят в реки, некоторые постоянно живут в пресной воде.
В нижеследующем дан обзор главнейших сельдевых как пресноводных, так и морских, свойственных нашим водам.
1 (18). Последний луч спинного плавника не вытянут в длинную
нить.
2 (5). По середине верхней челюсти заметная вырезка. Рот большой:
сочленение нижней челюсти с черепом заходит за вертикаль заднего края глаза.
3 (4). Зубы на сошнике есть. — Черное и Каспийское моря.
1. Caspialosa Berg
4 (3). Зубов на сошнике нет. — Балтийское море, изредка Черное.
2. Alosa Cuvier
5 (2). По середине верхней челюсти или совсем нет вырезки, или едва
заметная. Сочленение нижней челюсти с черепом не доходит до вертикали заднего края глаза.
6 (11). Последние два луча анального плавника не удлинены.
7 (8). Брюшные шипы слабые, едва заметные. На сошнике есть зубы
(слабые). Брюшные плавники у взрослых под передней третью спинного. Рыбы средней величины — 13 см и более. 4. Clupea L.
8 (7). Брюшные шипы явственные. На сошнике нет зубов. Мелкие
рыбы — до 13 см (редко до 15 см).
9 (10). Брюшные плавники или под самым началом спинного или даже
немного впереди спинного. Позвонков 45—50.
5. Sprattus Girgensohn
10 (9). Брюшные плавники несколько позади вертикали начала спинного.
Позвонков 39—44 (45). (Harengula Valenc.)
11 (6). Последние два луча анального плавника удлинены. На сошнике
нет зубов.
12 (13). На хвостовом плавнике нет удлиненных чешуй. Крышечная
кость гладкая. Брюшные шипы выдающиеся. Позвонков 40—49. 6. Clupeonella Kessler
13 (12). На хвостовом плавнике есть удлиненные чешуи.
14 (15). На крышечной кости нет радиальных бороздок. На заднем
крае жаберной камеры, на ключице, 2 кожистых выроста (на вертикальной части ключицы). Позвонков 44—48.
(Sardinella Valenc.)
15 (14). На крышечной кости есть радиальные бороздки. Таких выступов на вертикальной части ключицы нет. Позвонков 47—54.
16 (17). Жаберные тычинки верхней части жаберной дуги не перекрывают тычинок нижней части. — Атлантический океан, Средиземное море. (Sardina Antipa)
17 (16). Жаберные тычинки верхней части жаберной дуги перекрывают
тычинки нижней части. — Тихий океан, берега южн. Африки. (Sardinops Hubbs)
18 (1). Последний луч спинного плавника вытянут в длинную нить.—
Дальний Восток. 3. Clupanodon Lacepède
При счете числа позвонков за последний позвонок считается тот, который явственно загибается кверху перед началом хвостового плавника; с этим позвонком соединены последние рудиментарные позвонки в уростиль (см. Heincke, 1898, р. 77). Словом, за последний позвонок признается тот, который на рис. 5 обозначен номером 79. При счете килевых чешуй за чешуи перед брюшными плавниками считаются чешуи, лежащие впереди передней точки основания брюшных; чешуи за этой точкой считаются лежащими за брюшными плавниками. Длина тела определяется согласно нижеприведенной схеме (фиг. 88) у Caspialosa, Alosa, Clupeonella; у Clupea же и Sprattus все измерения отнесены к абсолютной длине тела, т. е. к длине от конца рыла до вертикали заднего конца нижней лопасти хвостового плавника.