Alosa caspia caspia (Eichwald, 1838)
Каспийский пузанок, северокаспийский пузанок, северо-восточный каспийский пузанок, астраханская селедка, ильменный пузанок, обыкновенный пузанок, пузанок, севернокаспийский пузанок, среднекаспийский пузанок, астаринский пузанок, бакинский пузанок, саринский пузанок, суворовский пузанок (Rus),
Caspian shad
(Eng)
Синонимы:
Alosa caspia (Eichwald, 1838) Северокаспийский пузанок (Rus),
Alosa caspia caspia (Eichwald, 1838) Северокаспийский пузанок (Rus),
Alosa caspia salina (Svetovidov, 1963) Северо-восточный пузанок (Rus),
Alosa rossica Kessler, 1870
Caspialosa caspia Eichwald, 1838 Каспийский пузанок (Rus),
Caspialosa caspia aestuarina Berg 1932 Ильменный пузанок, обыкновенный пузанок (Rus),
Caspialosa caspia caspia n. aestuarina Svetovidov, 1944
Caspialosa caspia kaidakensis Kazancheev 1936
Caspialosa caspia knipowitschi saraica Berg, 1948 Бакинский пузанок, саринский пузанок (Rus),
Caspialosa caspia knipowitschi sarensis Meysner, 1936
Caspialosa caspia persica Iljin, 1927
Caspialosa caspia salina Svetovidov, 1963 Северо-восточный каспийский пузанок (Rus),
Caspialosa caspia suvorovi Kisselevitz, 1923
Caspialosa suworowi Berg, 1913 Суворовский пузанок (Rus),
Caspialosa tanaica palaeostomi Sadowsky, 1934
Clupea caspia Eichwald, 1838
Clupeonella caspia Berg, 1913
Clupeonella caspia m. elongata Berg, 1913
Clupeonella suworowi Berg, 1913
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
Пузанок каспийский — Alosa caspia caspia Eichwald, 1838
Тело довольно высокое. Зубы на челюстях почти не развиты. Отличается многочисленными (около 120) тонкими и длинными жаберными тычинками. Зоопланктофаг. Длина примерно 30 см. Полупроходной вид, обитающий в северо-западной части Каспийского моря и в дельте Волги.
Декоративное рыбоводство. А.М.Кочетов 1991 г.
При содержании рыб воду лучше подсаливать (5—18 ‰). В аквариуме содержат следующие виды:
Каспийский пузанок - Caspialosa caspia Eichwald, 1838, 26 см; есть семь подвидов.
Словарь названий пресноводных рыб СССР. Г.У.Линдберг и А.С.Герд 1972 г.
3.1.2 (1). Alosa caspia Eichwald, 1838 — Северокаспийский пузанок (Св., 1952 : 242).
[1] русск. Астраханская селёдка — Гримм, 1878 : 39. Ильменный пузанок — Св., 1952 : 242. Каспийский пузанок — Суворов, 1904 : 40. Пузанок — В., 1916 : 27. Севернокаспийский пузанок — В., 1932 : 88. Северокаспийский пузанок — Св., 1952 : 242. Среднекаспийский пузанок — В., 1932 : 86. [18] англ. Caspian shad Канада — Ricker : 102.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
По форме головы, клиновидно сжатой
с боков в нижней части, пузанки близки к
европейско-американским алозам. Черноморско-каспийский пузанок (Alosa caspia) —
это в основном солоноватоводный вид,
живущий в воде самой различной солености:
черноморско-азовские пузанки входят для
нереста в пресную воду, каспийские
размножаются как в пресной, так и в
солоноватой воде в море. Икра пузанков
полупелагическая, при слабом течении на
нерестилищах опускается на дно; диаметр
икринок от (1,3) 1,5 до 3 мм.
Пузанки имеют высокое, сжатое с
боков, укороченное в области хвоста тело;
с большими глазами. На боках тела
обычно одно темное пятно позади
жаберной щели, нередко ряд из 6—8 темных
пятен. Зубы у пузанков развиты очень
слабо, едва заметные; жаберных тычинок
от 50 до 180, тычинки тонкие и длинные;
позвонков 47—51. Пузанки растут
медленнее проходных алоз и меньше их по
размерам: черноморско-азовские имеют длину
до 20 см, каспийские — до 28 см.
Все пузанки, проходные, полупроходные
или солоноватоводные, — чисто планктоноядные формы, ведущие пелагический
образ жизни. Пузанки — один из самых
теплолюбивых видов рода алоз.
В Каспийском море живут четыре
подвида пузанков: два — в Северном
Каспии и два — в Южном Каспии.
Наиболее многочислен северокаспийский
пузанок (Alosa caspia caspia), возможно,
распадающийся на две морфологически
неразличимые формы (племена): собственно
северокаспийскую и среднекаспийскую, или
ильменную. Северокаспийский пузанок
достигает длины 28 см, обычная длина
в уловах 18—22 см. Жаберных тычинок
у него на первой дуге от 70 до 149, тычинки очень тонкие, густые и длинные.
Позвонков 47—52. Это наиболее широко
распространенная форма вида,
встречающаяся почти по всему Каспийскому
морю. Половой зрелости достигает главным
образом в трехлетнем возрасте. Живет
до 9 лет. Северокаспийский пузанок
зимует в южной части Каспийского моря,
держась в теплые зимы западнее, а в
холодные — восточнее, преимущественно на
глубине 24—33 м от поверхности при
температуре воды 9—11°С. Весной, начиная
с марта, совершает миграции на север
вдоль западного берега Каспия. В
Среднем Каспии подходит к западному берегу
в апреле и в мае при температуре воды
7,6—10,2°С и 10,8—14,0°С; при температуре
ниже 5° не встречается. В первом подходе
преобладают самцы, во втором — самки.
В Северном Каспии появляется в конце
марта — начале апреля, широко
распространяясь в западной части моря в мае.
Нерестует почти по всему мелководью
Северного Каспия, наиболее интенсивно
в северо-западной части, в предустьевом
пространстве Волги. Входит и в дельту
Волги, заходя в ильмени, поднимается
для икрометания и выше дельты, в
небольшом количестве до Волгограда и выше.
В Урал заходит в небольших количествах.
Места массового нереста располагаются
в предустьевом пространстве,
преимущественно на глубине 1—3 м, меньше до
6 м; нерест начинается с конца апреля —
середины мая и заканчивается в
середине — конце июня, происходит при
температуре от 13,8 до 24°С, главным образом
от 18 до 22°, в основном в пресной или
в осолоненной до 1—2‰ воде, частично
до 4—6 и даже 8,4‰. В дельту Волги
и выше пузанка заходит мало. Икринки
северокаспийского пузанка и ильменной
формы его различаются: у основной
формы икринки имеют большие размеры
(1,7—3,0 мм против 1,39—1,99), большее
кругожелтковое пространство (21,8—
31,3% против 13,5—26,5, в среднем 20%
диаметра икринки), наконец, оболочка
икринок основной формы тонкая и
нелипкая, как у всех видов основного подрода алоз, тогда как у
икринок ильменного пузанка оболочка плотная,
импрегнированная мелкими частицами ила,
по-видимому, так, как у американских помолобов.
Питается северокаспийский пузанок
главным образом мелкими веслоногими
рачками планктона, меньше мизидами;
интенсивность питания в зимний период
очень понижена. Жирность его
колеблется от 6,3—10,3% весной, до
18,1%—осенью. Растет пузанок очень медленно,
достигая в годовалом возрасте длины
11 — 12,4 см, в два года — 16,1—17,4,
в три — 18,9—20,9 и в четыре — 21,0—
23,0 см.
Северокаспийский пузанок — одна из
важнейших промысловых сельдей
Каспийского моря, дававшая от 40 до 75%
всего улова сельдей в этом водоеме.
В 1927—1930 гг. были сделаны попытки
акклиматизировать северокаспийского
пузанка в Аральском море. Они не
увенчались успехом.
Остальные подвиды каспийских
пузанков — северо-восточный (Alosa caspia salina),
энзелийский (Alosa caspia knipowitchi),
астрабадский (Alosa caspia persica) — имеют
гораздо меньшее значение, чем
северокаспийская форма. Северо-восточный
пузанок в 30-х годах был распространен в
восточных водах Каспия и нерестовал в
солоноватой воде на мелководье перед
входом в залив Мертвый Култук. Этот залив
высох в 40-х годах вследствие падения
уровня Каспия, а более поздних
исследований северо-восточного пузанка не
было.
Фауна СССР. Рыбы. Том II, вып.1. Сельдевые (Clupeidae). А.Н.Световидов 1952 г.
*3. Alosa caspia (Eiclrwald) s. l. — Каспийско-черноморские пузанки.
Литературу см. при подвидах. В числе работ по черноморским подвидам не упомянуты две статьи, опубликованные в недоступном автору издании (Borcea I., С. R. Acad. Sci. Roumanie, I, 1936 : 26—28, 216—217).
Жаберных тычинок 50—180, различное количество у разных форм, тычинки у всех форм тонкие и длинные, значительно длиннее жаберных лепестков, с необломанными концами, обычно образующими на нижней половине жаберной дуги выпуклую линию. Зубы развиты очень слабо, на верхней челюсти обычно слабо заметные на ощупь, иногда на нижней, реже и на верхней совершенно не заметные. Челюсти одинаковой длины или нижняя слегка выдается вперед, реже короче верхней. Голова клиновидно сжатая в нижней части с боков, довольно большая и высокая, высота ее 17.5—25% длины тела, значительно различающаяся у разных форм. Глаза большие, размеры их у разных форм неодинаковые. Тело типично пузанковое, высокое, укороченное в хвостовой области, с длинными грудными плавниками, высота его у разных форм различная. Темп роста даже у наиболее быстрорастущей типичной формы вида по сравнению с каспийскими проходными сельдями, крайне замедленный; при одном и том же возрасте, начиная со второго года, размеры у каспийских в среднем на 5 см и более меньше, чем у Alosa kessleri volgensis. Сравнительно небольшие по размерам рыбы, каспийские до 28 см, наичаще 18—22 см, черноморско-азовские не свыше 20 см, наичаще 14—16 см.
Широко евригалинный, в основном солоноватоводный вид, черноморские фррмы которого заходят для нереста в пресную воду, каспийские размножаются как в пресных водах, так и в солоноватой воде в море.
Распадается на ряд форм. Один из наиболее изменчивых экологически видов, занимая в этом отношении промежуточное положение между морскими и проходными сельдями, так как имеет формы как полупроходные, мечущие икру в пресной или слегка осолоненной воде, так и почти морские, размножающиеся в солоноватой или даже с океанической соленостью воде. Один из наиболее теплолюбивых видов рода, причем некоторые из самых теплолюбивых каспийских форм живут и размножаются в южной части Каспийского моря, другие для нереста идут в северную часть его. Черноморско-азовские формы несколько менее теплолюбивы, чем каспийские. Морфологически некоторые формы вида, в особенности черноморско-азовские, отличаются друг от друга не слишком сильно, что стоит в связи с тем, что по некоторым биологическим особенностям, обусловливающим морфологические различия у форм других видов рода, формы этого вида сходны — все они планктоноядны и ведут пелагический образ жизни, между тем как более сильно различающиеся морфологически формы других видов рода, например Alosa brashnikovi s. l., в отношении характера питания и образа жизни сильно различаются. Однако по ряду особенностей, в частности по количеству жаберных тычинок и позвонков, длине головы и высоте тела и некоторым другим и даже внешне, почти все формы отличимы довольно хорошо, а некоторые, как, например, Alosa caspia knipowitschi, от большинства форм отличаются резко. При этом следует отметить, что черноморско-азовские формы, отличаясь некоторыми особенностями (меньшие размеры, менее высокое тело, большие размеры глаз) от каспийских, между собой различаются незначительно, гораздо менее, чем каспийские. О разделении форм этого вида по антедорсальному и антеанальному расстоянию, длине головы, количеству позвонков и некоторым другим особенностям на две группы — северных и южных и по количеству жаберных тычинок на западных и восточных см. Световидов, 1950.
Распространение. Каспийское, Черное и Азовское моря. В Каспийском море некоторые формы, нерестящиеся в северной части его, распространены почти повсеместно, другие почти не выходят из пределов южной части.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДВИДОВ
В числе их не упомянута новая форма из пресных вод Турции, описанная в недоступном автору издании [(Battalgil F. Türkiyenin tatli su baliklari. Les poissons des eaux douces de la Turquie. (Collection de l'InsLitufc de Zoologie de l'Universite d'Istambul). Rev. Fac. Sci. Univ. Istambul, VI B, 1941 : 170—186). (Caspialosa tanaica etemi subsp. nov., оз. Маньяс, басс. Мраморного моря)].
1 (8). Глаза не особенно большие, 18—24.5%, в среднем около 21.5—22.5% длины головы. Довольно крупные рыбы длиной до 28 см, в половозрелом состоянии от 14—16 см. Каспийское море.
2 (5). Жаберных тычинок 70—180, наичаще 100—160, тычинки длинные, тонкие и густые. Западная половина Каспийского моря.
3 (4). Жаберных тычинок 70—150, наичаще 100—140. Позвонков 47—52, наичаще 49—51. Голова сравнительно небольшая, 25.5—28%, в среднем около 26.5% длины тела. Высота головы 18.5—22%, в среднем около 20.5% длины тела. Грудные плавники сравнительно короткие, 15.5—18%, в среднем около 16.5—17% длины тела. Тело не особенно высокое. Преимущественно в западной половине Каспийского моря, зимой в южной части и в восточной половине моря. *3f. Alosa caspia caspia (Eichwald)
4 (3). Жаберных тычинок 120—180, наичаще 130—160. Позвонков 43—49, наичаще 47—48. Голова большая, 25—30%, в среднем около 28% длины тела. Высота головы 21—25%, в среднем около 22.5—23% длины тела. Грудные плавники длинные, 16—19%, в среднем около 17.5% длины тела. Тело значительно выше. Южная часть Каспийского моря, преимущественно в западной половине, осенью единично вдоль западного берега средней части. *3d. Alosa caspia knipowitschi (Iljin)
5 (2). Жаберных тычинок 50—120, наичаще 80—100, тычинки короче, более грубые и редкие. Восточная половина Каспийского моря.
6 (7). Жаберных тычинок 60—120, наичаще 80—100. Позвонков 48—52, наичаще 49—51. Голова сравнительно небольшая, 24—26.5%, в среднем около 25.5% длины тела. Высота головы 17.5—21.5%, в среднем около 19.5% длины тела. Грудные плавники сравнительно короткие, 15—17.5%, в среднем около 16% длины тела. Тело не особенно высокое. Восточная половина Каспийского моря, весной и летом в восточной половине северной части моря. *3g. Alosa caspia salina (Svetovidov)
7 (6). Жаберных тычинок 50—110, наичаще 60—90. Позвонков 45—51, наичаще 47—49. Голова большая, 25.5—27%, в среднем около 27% длины тела. Высота головы 19.5%—20.5%, в среднем около 20% длины тела. Грудные плавники длинные, 16.5—18%, в среднем около 17% длины тела. Тело очень высокое. Южная часть Каспийского моря, преимущественно в восточной половине. *3е. Alosa caspia persica (Iljin)
8 (1). Глаза большие, 22—28.5%, в среднем около 24%—26% длины головы. Небольшие рыбы длиной не более 20 см, в половозрелом состоянии от 10 см. Черное и Азовское моря.
9 (12). Тело низкое, наибольшая высота его составляет 20%—27%, в среднем около 24.5% длины его.
10 (11). Глаза большие, 24—28.5%, в среднем около 26% длины головы.
Жаберных тычинок 62—85, в среднем около 76. Азовское море и
прилегающая часть восточной половины Черного моря. *3с. Alosa caspia tanaica (Grimm)
11 (10). Глаза несколько меньше, 22—27%, в среднем около 24.5% длины головы. Жаберных тычинок 66—88, в среднем около 80. Западная половина Черного моря. *3b. Alosa caspia nordmanni Antipa
12 (9). Тело выше, наибольшая высота его 25—31%, в среднем около 27.5% длины его. Юго-восточная часть Черного моря, главным образом у Поти и Анаклии. *3а. Alosa caspia palaeostomi (Sadowsky)
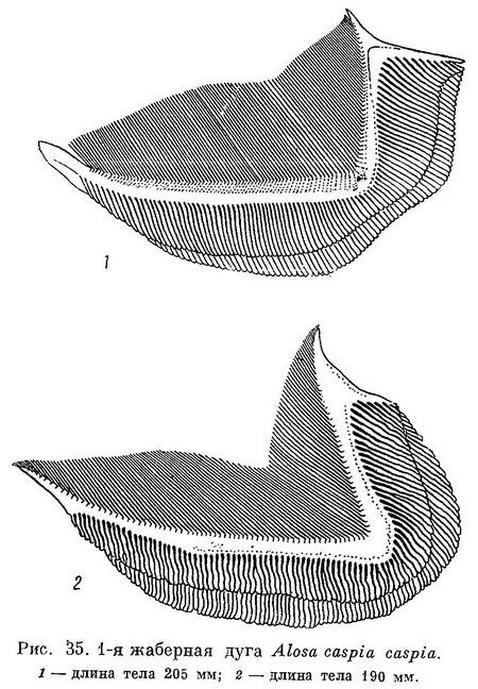
*3f. Alosa caspia caspia (Eichwald) — Северокаспийский (среднекаспийский) или ильменный пузанок (табл. XII, XIII).
— Clupea caspia Eichwald, Bull. Soc. Nat. Moscou, XI, 1838 : 134 (Каспийское м., к югу); Fauna caspio-caucasica, 1841 : 203, t. XXXII, f. 1 (Каспийское м., к югу).
— Alosa rossica Кесслер, Тр. СПб. общ. ест., I, 1870 : 76 (Волга, nomen nudum; согласно Кесслеру = Clupea caspia Eichwald, 1841).
— Alosa caspia Кесслер, там же : 282 (Волга у Самары, Казани, Ярославля (Один из этих экземпляров хранится в коллекциях кафедры ихтиологии Ленинградского Государственного университета, № 405, жаберных тычинок 127, длина 29.7 см.), сентябрь 1869 г.).
— Alosa caspica Яковлев, Тр. Общ. ест. Казанск. унив., I, 2, 1871 : 63—64 (жаберных тычинок свыше 100).
— Clupea caspia Кесслер, Тр. СПб. общ. ест., V, 1, 1874 : 301 (частью); Гримм, Астраханская селедка, СПб., 1887 : 67 (широкая разность).
— Clupea (Alosa) caspia Суворов, Тр. Касп. эксп. 1904 г., I, 1907 : 142, 164, 168—172 (среднекаспийский пузанок, западное побережье Каспийского м. от Махачкалы до Апшеронского п-ова; измерения), 146, 164, 178—180 (астаринский пузанок, от Апшеронского п-ова до устья р. Астары; измерения); там же, II, 1908 : 40 (частью).
— Clupeonella caspia Берг, (Несколько более подробные ссылки на некоторые синонимы и литературу более раннего периода см. в этой работе и в цитируемой ниже работе Киселевича (1923).) Мат. позн. русск. рыбол., II, 3, 1913 : 36, табл. I, фиг. 1, табл. II (n. caspia).
— Clupeonella caspia m. elongata Бepг, там же : 38 (n. aestuarina).
— Clupeonella caspia Киселевич, там же, III, 6, 1914 : 14, 40, 48—50 (северо-восточная часть Каспийского м.; измерения, ход).
— Caspialosa caspia Берг, там же, IV, 6, 1915 : 4; Рыбы пресных вод, 1916 : 27 (частью); Киселевич, Каспийско-волжские сельди, I, 1923 : 108, 135, 49—128 прилож. (n. aestuarina, частично n. caspia); Морозов, Тр. Научн. инст. рыбн. хоз., II, 4, 1928 : 47 (биометрическая обработка материалов Киселевича).
— Caspialosa caspia caspia Аверинцев и Сыч, Тр. Астрах. научн. рыбохоз. ст., VIII, 3, 1930 : 11 (количества позвонков, килевых чешуй, лучей в плавниках).
— Caspialosa caspia caspia Световидов, Докл. АН СССР, 1932 : 203 (количество пилорических придатков).
— Caspialosa caspia aestuarina Берг, Рыбы пресных вод, I, 1932 : 88 фиг. 94. За тип n. aestuarina следует считать экземпляр длиной 205 мм из ильменя Дамбур, 19 V 1912 г., описанный Бергом (1913 : 32, табл. 3).
— Caspialosa caspia Берг, там же : 86, фиг. 93; Дехтярева. Тр. Всесоюзн. Научно-исслед. инст. морск. рыбн. хоз. и океаногр., XIV, 1940 : 47 (описание сеголетков); Световидов, в: Промысловые рыбы СССР, 1949 : 95, табл. 23.

Распространение, биология. Арнольд, Вестн. рыбопр., XXI, 9—10, 1906 : 480 (Волга в 30 км ниже Симбирска, 30 VI 1906, 112 жаб. тыч.); Тр. Касп. эксп. 1904 г., I, 1907 : 226, фиг. 3 (зал. Чада; искусственное оплодотворение); Мейснер, Мат. позн. русск. рыбол., III, 4, 1914 : 97, фиг. VI—XVII (кавказское побережье; биология, промысел); Киселевич, там же, III, 6, 1914 : 3 (северо-восточная часть моря; биология); Вукотич, там же, IV, 6, 1915 : 21 (Кизил-бурун; плодовитость); Киселевич, там же, IV, 10, 1915 : 119 (n. aestuarina, дельта Волги; ход, нерест); Тр. Астрах. ихт. лаб., V, 1, 1923 : 37, 41 (n. aestuarina ?, у устьев Волги; плодовитость); Киселевич, там же, V, 2, 1922 : 22 (рост, возрастной состав); Фаресов, там же, 143 (низовье р. Эмбы); Киселевич, там же, VI, 1, 1924 : 70 и сл. (n. aestuarina ?, возраст и темп роста); Недошивин, Сборн. в честь Книповича, 1927 : 304 (искусственное оплодотворение); Мейснер, Бюлл. Всекасп. научн. рыбохоз. эксп., 1—2, 1932 : 50 (среднекаспийский и северокаспийский пузанок, миграции); там же, 5—6, 1932 : 20 (среднекаспийская раса); Чугунов, там же, 3—4, 1932 : 13 (распределение и миграции в море в весенне-летний и осенний периоды); там же, 5—6, 1932 : 191 (распределение и миграции в море весной); Леванидов, там же, 5—6, 1932 : 53 (содержание жира); Жуковский, Тр. Северн. инст. рыбн. хоз., III, 1932 : 37 и сл. (акклиматизация в Арале); Махмудбеков, Рыбн. хоз. СССР, 1935, 11 : 22 (половой состав и изменение его во время хода); Чаянова, Тр. I Всекасп. научн. рыбохоз. конфер., I, 1936 : 51 (питание); Дмитриев, Рыбн. хоз. СССР, 1936, 2 : 39 (миграции у западных берегов моря); Беляев, Первая научн. конфер. по изуч. сельдевых рыб Каспийского м., Астрах., 1937 : 34 (распространение, нерест, возраст); Керенцева, Учен. зап. Пермск. Гос. унив., III, 1, 1937 : 177 (у восточных берегов южной части Каспийского м., Тарта, Карши, Кизил-су; апрель); Киселевич, Сельди северного Каспия, Сталингр., 1937 : 25 (биология); Навозов-Лавров, Рыбн. хоз., 1938, 4—5 : 26 (Дагестан; условия подхода в береговую зону); Перцева, там же, 1938, 7 : 31, рис. 1—2 (n. caspia, места, время и условия нереста); Н. Танасийчук, там же, 1938, 11 : 33 (нерест в затонах Волги); Строганов, Учен. зап. Моск. Гос. унив., 33, 1939 : 201 (Кардуанские ильмени; нерест, развитие икры при разных соленостях); Перцева, Тр. Всесоюзн. Научно-исслед. инст. морск. рыбн. хоз. и океаногр., VIII, 1939 : 27 (развитие и морфология икринок и личинок); Дмитриев, Сборн., посвящ. деят. Книповича, 1939 : 266. (условия подходов в береговую зону); Махмудбеков, Изв. Азербайджанск. научно-исслед. рыбохоз. ст., 3, 1939 : 3 (расовый состав и термические условия подходов пузанков); там же : 27 (возраст и темп роста); Замахаев, Тр. Всесоюзн. Научно-исслед. инст. морск. рыбн. хоз. и океаногр., XIV, 1940 : 3 (нерестовые кольца); Казанова и Халдинова, там же : 77 (n. aestuarina и n. caspia, дельта Волги; икринки и личинки); Перцева, там же : 109 (n. caspia; нерест, литература); Чаянова, там же : 211 (питание); Махмудбеков, Рыбн. хоз., 1941, 3 : 18 (возрастной состав, перспективы промысла); Казанчеев, Рыбн. промышл. СССР, 3, 1945 : 59 (берега Мангышлака; летом); Екатерининская и Изосимов, Тр. Общ. ест. Казанск. Гос. унив., 1—2, 1945 : 37 (Волга выше Тетюшей; питание после нереста); Штейнфельд, там же : 76 (Волга у Тетюшей и выше Казани); Смирнов, Изв. Акад. наук Азерб. ССР, 7, 1945 : 71 (миграции); там же, II, 8, 1946 : 55 (биология); Докл. Акад. наук Азерб. ССР, II, 2, 1946 : 60 (влияние света на нерестовые миграции); Махмудбеков, Рыбн. хоз., 1946, 9 : 6 (состояние запасов); Зоолог. журн., XXVI, 2, 1947 : 143 (возраст наступления половой зрелости); Остроумов, Тр. Волго-каспийск. научн. рыбохоз. ст., IX, 1, 1947 : 3 (миграции); там же, X, 1948 : 97 (возрастной состав нерестового стада); В. Танасийчук, там же : 93 (нерест в Урале); Остроумов, Зоолог. журн., XXVIII, 5, 1949 : 447 (половое созревание); В. Танасийчук, Тр. Всесоюзн. Научно-исслед. инст. морск. рыбн. хоз. и океаногр., XVIII, 1951 : 20, рис. 3, 4 (колебание урожайности молоди); Перцева-Остроумова, там же : 53, рис. 9, 10 (описание икринок и личинок). Н. Танасийчук, промысл. рыбы Волго-Каспия, Пищепромиздат, 1951 : 39 (распространение, миграции, места нереста).

D III—IV 13—15 (13.7), общим числом 17—19 (17.7), A III (IV) 15—20 (18.3), общим числом 18—23 (21.3), sp. br. 68—135 (116.0), vert. S. 49—51 (50.05), caud. vert. 33—34 (33.26). Жаберные тычинки очень тонкие, густые и длинные, в 1.5—2 раза длиннее жаберных лепестков. Зубы очень слабые, на нижней челюсти почти не развитые и обычно не заметные на ощупь, на верхней челюсти едва заметные. Голова сравнительно небольшая и низкая, длина ее 25.5—28.1 (26.4)%, высота 18.8—22.0 (20.5)% длины тела. Глаза довольно большие, 19.5—24.4 (22.2)%. длины головы. Грудные плавники относительно короткие, 16.0—18.1 (16.7)% длины тела. Тело высокое, наибольшая высота его 22.8—29.6 (26.4)%, наименьшая — 6.8—8.9 (7.8)% длины. Спина и верхняя часть головы темные. За жаберной крышкой почти всегда с каждой стороны по темному пятну, сзади которого на боках иногда еще по нескольку пятен (3—7). Размеры до 28 см, наичаще 18—22 см. Описание дано по 50 экз. длиной от 15.1 до 22.1, в среднем 18.7 см, собранным весной, (во второй половине апреля — начале мая) у западных берегов средней части Каспийского моря (Сумгаит и Турали). По данным В. И. Мейснера, у n. caspia жаберных тычинок 70—149 (121.5), позвонков 47—52 (49.9), у n. aestuarina жаберных тычинок 92—149 (116.7), позвонков 45—52 (49.6), пилорических придатков (Световидов, 1932) 30—59 (40.8).
Евригалинная форма, размножается как в солоноватой, так и в пресной воде, иногда высоко подымаясь вверх по течению.
Возможно, распадается на 2 формы: Alosa caspia caspia n. caspia — среднекаспийский или точнее северокаспийский пузанок и
Alosa caspia caspia n. aestuarina — ильменный пузанок. Морфологически обо эти формы в сущности не различимы. По данным
B. И. Мейснера, у Alosa caspia caspia n. aestuarina несколько меньше количество жаберных тычинок, немного короче голова и крупнее глаза, но все эти различия крайне ничтожны, едва отличаясь в средних и совпадая в пределах колебаний. По материалам Н. Н. Коркиной, исследовавшей большое количество проб пузанка из разных районов северной части моря и из ильменей, реальные различия имеются между всеми пробами почти во всех исследованных ею признаках, кроме количества позвонков, но все эти различия не велики и имеют крайне неопределенный характер: пробы из ильменей почти не отличаются от проб из моря, причем по большинству исследованных признаков отличаются друг от друга пробы и из ильменей. Общее заключение из данных Н. Н. Коркиной таково, что различия между этими формами имеются только в темпе роста, чем, возможно, и обусловлены меньшие размеры и более удлиненная форма ильменного пузанка (Казанова и Халдинова, 1940). Относительно мест икрометания обеих форм указания довольно разноречивы (см. об этом у Киселевича, 1937 : 29 и сл.), но возможно, что северокаспийский пузанок мечет икру в море, а ильменный в ильменях (см. ниже). Имеются указания, что обе формы существенно различаются по строению икринок. У ильменного пузанка икринки значительно мельче, чем у северокаспийского (диаметр их у ильменного пузанка, как правило, не превышает 2 мм, между тем как у северокаспийского он обычно более 2 мм), оболочка плотнее и поверхность ее вследствие клейкости покрыта тонким слоем ила, чего не наблюдается у северокаспийского пузанка. Однако остается неясным, в пресной ли воде найдены эти икринки северокаспийского пузанка или в солоноватой. Таким образом, вопрос о существовании двух рассматриваемых форм пузанка требует еще дальнейшего исследования, и поэтому, учитывая также отсутствие отчетливых различий между ними, их никак нельзя признать равнозначными другим каспийским формам вида. По этим причинам ильменный пузанок принимается провизорно как natio и рассматривается, подобно другим формам с неопределенным характером отличий, совместно с типичной формой.
Alosa rossica Кесслера (1870) следует считать за Alosa caspia caspia, так как на стр. 77 цитируемой работы Кесслер указывает, что на рисунке Clupea caspia Eiclrwald (1841) изображена эта форма. К Clupea caspia Кесслер (1874) частично, судя по указываемым размерам (до 345 мм), относится и Alosa kessleri volgensis. Сюда же он относит и каспийские формы Alosa caspia (l. с. : 113).
Распространение. Каспийское море. Наиболее широко распространенная форма вида. Встречается почти по всему морю, за исключением, повидимому, самых северо-восточных районов северной части его. В наибольшем количестве встречается на путях к местам нереста вдоль западного побережья моря и на местах нереста. Икрометание почти по всему мелководью северной части моря от Аграханского зал. до Урало-эмбенского района, наиболее интенсивно в предустьевом пространстве Волги между течениями Главной и Белинской банок и восточнее в районе Жесткого и Новинского осередков, иногда к юго-западу от предустьевого пространства в районе Лагани. У юго-западных берегов северной части моря и в Урало-эмбенском районе нерест менее интенсивен. В небольшом количестве входит в дельту Волги, откуда для икрометания заходит в западные и отчасти в восточные подстепные ильмени. В годы с низкими паводками, когда ильмени не затапливаются, подымается для икрометания и выше дельты, в небольшом количестве заходит до Сталинграда и выше (до Куйбышева и выше (Коллекции Зоологического института Академии Наук СССР, № 30987, Волга, Климовский затон (110 км выше Куйбышева), 23 V 1939, жаберных тычинок 124, длина 49.3 см.) и Казани, в прежние годы даже до Ярославля (№ 405 коллекций Ленинградского Государственного университета, Ярославль, 1869, Кесслер. Об одном из этих экземпляров пишет Кесслер (1870). Это действительно Alosa caspia caspia.)), единично — в Каму выше Молотова. (Кама у с. Добрянки (60 км выше Молотова), 2 IX 1951, № 32875 коллекций Зоологического института Академии Наук СССР, доставлен Ю. А. Козьминым.) В небольшом количестве заходил в прежние годы в низовья Эмбы, заходит в Урал.
Биология. Нерест во второй половине мая, начинается он в зависимости от температуры воды с конца апреля — начала мая до середины мая и заканчивается в середине — конце июня. Зимует в южной части моря, причем в более теплые зимы основная масса держится против западных берегов и в меньшей степени против восточных, встречается и в средней части, в холодные же зимы концентрируется главным образом против восточных берегов и в меньшем количестве против западных, встречается, вероятно, и против южных берегов. Одна из наиболее теплолюбивых из нерестящихся в северной части моря сельдей, в море при температуре воды ниже 5° не встречается, при 5° держится в разреженном состоянии, основные концентрации в зимнее время наблюдаются при 9—11°. Поэтому, когда температура воды против западных берегов южной части моря ниже этих оптимальных температур зимних концентраций, то основная масса отходит в восточную, более теплую половину моря. В теплые зимы зимует далее к северу, в средней части моря (Н. Танасийчук, 1951). Наибольшие зимние скопления образует в местах резкого падения глубин (от 60 до 170 м и больше), где благодаря нисходящим токам создаются наилучшие условия аэрации слоев воды, причем основная масса держится в постоянном течении, вне его, в галистатической области встречается в небольшом количестве. С наступлением весны, в марте, начинает передвигаться на север, при этом, как показали исследования П. К. Дорошкова, миграции в основном происходят вдоль западного побережья, сначала южной, а затем средней частей моря в северную часть заходит у западных берегов (Остроумов, 1947). У восточных берегов средней части моря идет в значительно меньшем количестве и, возможно, не эта форма, a Alosa caspia salina. Относительно захода основной массы северокаспийского пузанка в северную часть моря у восточных берегов (Киселевич, 1914), а также и о фронтальном подходе его в северную часть (Чугунов, 1932) см. у Остроумова (1947). В весенний период миграций на север в центральных областях южной и средней частей моря встречается редко, и в открытом море значительных концентраций не образует, передвигаясь на север в основном в частях моря, не слишком удаленных от берега. В апреле подходит к восточным берегам (промысел Тарта, Карши, Кизил-су) в южной части моря (Керенцева, 1937). Подходы в прибрежную область в средней части моря у западных берегов происходят в апреле—мае, одновременно на большом протяжении, причем здесь наблюдаются две волны массовых подходов — первая в зависимости от условий весны обычно во второй половине апреля при температуре воды 7.6—10.2°, вторая — в первой половине мая при 10.8—44.0°. Первая волна обычно больше второй. В первой волне преобладают самцы (свыше 80%), идущие в начале хода, во второй самки (свыше 70%), идущие в конце хода. Каких-либо морфологических различий между пузанками этих двух волн подходов нет (Махмудбеков, 1935). Подходы в узкую прибрежную область обусловлены, с одной стороны, несколько более высокими температурами воды здесь, чем в открытом море, с другой стороны, меньшими скоростями течений, причем в зоне неводного облова миграции вдоль берегов совершает крайне ограниченные. При весенних миграциях на север вместе с половозрелыми идет и молодь, еще не достигшая половой зрелости. В мае миграции в южной и средней частях моря заканчиваются, подходы в прибрежную область становятся меньше, и в середине мая промысел обычно заканчивается. В северной части моря первые экземпляры обычно появляются в конце марта — начале апреля. Первое время держится разрозненно в глубинных, осолоненных и более тепловодных участках; в предустьевом пространстве появляется позднее, в западной половине скопления образует через 15—20 дней после появления здесь. В мае места концентрации занимают обширную площадь в западной половине, простирающуюся на восток до Забурунья (Остроумов, 1947). В период миграций в средней части моря имеет половые продукты в стадии III, III—IV и IV. Икрометание происходит при температуре воды от 13.8 до 24.1°, массовый нерест — при 18—22°, как в пресной, так и в слегка осолоненной воде с содержанием солей от 1—2‰; в небольшом количестве развивающиеся икринки были находимы и в более осолоненной воде, до 4—6‰ и даже при 8.4‰ (в восточной половине северной части моря против Гогольского Култука). Места массового нереста располагаются в предустьевом пространстве на глубине от 1 до 3 м) в меньшем количестве нерестует и на больших глубинах, до 6 м. В дельте Волги икринки среднекаспийского пузанка найдены единично, преимущественно в западных ильменях, при температуре воды в июне от 18.2 до 19.7°. Икринки ильменного пузанка найдены в западных ильменях, в мае при температуре воды 19.2—21.8°, в июне — при 22—23.4°, в предустьевом пространстве моря, а также в дельтовых протоках не встречены. В 1914 г. нерест происходил как в западных, так и в восточных ильменях, куда перестал заходить вследствие обмеления и зарастания. Встречается и в ильменях, расположенных выше, — в ильмене Кордонном в системе р. Ахтубы. Количество пузанка, заходящего в дельту и выше, не велико: в 1937 г. в предустьевом пространстве Волги в промысловых уловах сельдей он составлял 73.5%, а в дельте лишь 7.8% (Остроумов, 1947; см. также Киселевич, 1915). Однако временами в Волгу заходит в значительных количествах. Так, например, в 1938 г. в первой пятидневке мая у с. Никольского (240 км от Астрахани) составлял до 27.7% неводного улова, в среднем за май — 1.2% (Н. Танасийчук). В значительных количествах заходил, повидимому, в 60-х годах (Кесслер, 1870). Плодовитость от 12 до 41 тыс., в среднем около 24 тыс. икринок (Вукотич, 1915), рабочая плодовитость от 7 до 7.5 тыс. икринок (Жуковский, 1932). Инкубационный период при температуре воды около 20° длится от 42 до 52 час., в среднем 46 час. (Перцева, 1939). Молодь, по наблюдениям В. С. Танасийчук и А. И. Дехтяревой, в наибольших концентрациях встречается в июле (По сообщению Н. П. Танасийчука, молодь в 1938—1939 гг. была находима в отшнуровавшихся затонах нижнего течения Волги. По словам рыбаков, молодь пузанка здесь успешно перезимовывает.), держится на сравнительно неглубоких местах вблизи нерестилищ, глубже 5 м не найдена, по отношению к солености, а также, повидимому, и к температуре строгой приуроченности нет, но из северной части на юг уходит ранее других сельдей, в октябре и ноябре здесь уже не встречается. На юг мигрирует вдоль западных берегов, близко к берегу. В северной части моря взрослые остаются довольно долго, однако большая часть уже летом уходит в среднюю часть, где в значительных концентрациях обнаружена у восточного побережья (Н. Танасийчук, 1951). Обратные миграции из северной части в среднюю происходят как у западных берегов, так и в центральной части моря и у восточных берегов, причем у восточных берегов к югу, начиная приблизительно от зал. Кендырли, встречается в меньших количествах, повидимому, отходя отсюда в центральные части моря. В конце октября — начале ноября основные массы переходят из средней части в южную, при этом переход совершается главным образом у западных берегов, так как в восточной половине встречается в значительно меньших количествах и, возможно, не эта форма, a Alosa caspia salina. Вертикальное распространение известно мало. В зимнее время, по наблюдениям П. К. Дорошкова, основная масса держится на глубине 24.5—32.5 м от поверхности, более или менее глубоко встречается в меньших количествах. Весной держится в поверхностных слоях воды, опускаясь при сильном летнем нагревании воды опять в более глубокие слои. Осенью вновь поднимается к поверхности, и основная масса держится на глубине 0.5—16.5 м. Основу питания составляют планктонные ракообразные (Copepoda — более 70%, Mysidae — около 20%, Amphipoda и Cumacea — единицами процентов). Питание в разное время года существенно отличается. Различают четыре периода: 1) в зимний период во время пребывания на местах зимовки питается очень слабо, 2) в период преднерестовых миграций питается интенсивно, 3) в нерестовый период питается также интенсивно и 4) в посленерестовый период миграций на юг питается особенно интенсивно (Чаянова, 1940). Во время нереста питается и в ильменях, так как желудки его, по наблюдениям И. И. Кузнецовой, в этот период здесь полны пищи. В реке (Волга, выше Тетюшей) после нереста основную роль в питании играет Daphnia longispina (63—77%), Cyclops strenuus (20—30%) и другие ракообразные (Екатерининская и Изосимов, 1945). Половой зрелости самки и самцы достигают в двухгодовалом возрасте; самцы несколько в большем количестве, частично в четырех пятигодовалом, но основная масса тех и других созревает в трехгодовалом возрасте и в течение жизни нерестует 2—3 и даже 4 раза (Махмудбеков, 1947; Остроумов, 1948, 1949). В уловах у самцов преобладают двух- и трехгодовалые (до 94%), у самок — трех- и четырехгодовалые (81%). Самки растут несколько быстрее самцов. Возрастной состав из года в год изменяется. У берегов Азербайджана за ряд лет он был таков (Махмудбеков, 1941; в процентах):
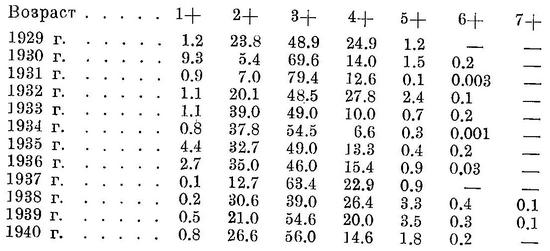
Параллельно со снижением возрастного состава за ряд последних лет уменьшились и средние размеры ловимого пузанка, которые к 1940 г. обнаружили тенденцию к повышению (Махмудбеков, 1941):

Значительно снизились и средние размеры возрастных групп, особенно, для 1934—1936 гг. (Махмудбеков, 1941; см):

В 1927—1930 гг. были предприняты попытки акклиматизации пузанка в Арал, не давшие, однако, положительного результата, но единичные взрослые экземпляры в Арале были пойманы. (Один экземпляр хранится в коллекциях Зоологического института Академии Наук СССР, № 25622, Уч-сай, 14 V 1931, жаберных тычинок 97, длина 18.8 см.) После черноспинки одна из самых жирных из каспийских сельдей. В начале весенних миграций у берегов Азербайджана и Дагестана содержит от 9.01 до 13.19% жира, в северной части моря содержание жира уменьшается до 6.34—10.3%. Наиболее жирной бывает осенью, у берегов Дагестана в сентябре содержит до 18.10% жира (Леванидов, 1932).
Хозяйственное значение. Одна из важнейших промысловых рыб Каспийского моря, стоящая по уловам на первом месте среди; каспийских сельдей. Уловы в 1936—1939 гг. составляли от 56 до 750 тыс. ц. Основная масса улова добывается у берегов Дагестана и Азербайджана и в северной части моря, весной во время миграций на север и на местах нереста и отчасти зимой в южной части моря.
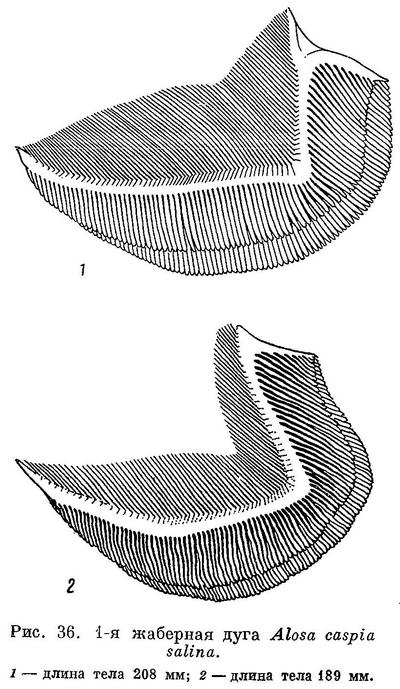
*3g. Alosa caspia salina (Svetovidov) — Северо-восточный каспийский пузанок (табл. XIV).
— Caspialosa caspia Суворов, Тр. I Всесоюзн. Геогр. съезда, 3, 1934 : 14 (у входа в зал. Мертвый Култук с восточной стороны м. Кызыл-каир).
— Caspialosa caspia salina Световидов, Докл. АН СССР, I (X), 1 (78), 1936 : 51 (в конце мая — в июне в северо-восточной части Каспийского моря (Тип № 25813 коллекций Зоологического института Академии Наук СССР, зал. Мертвый Култук в 10 км к западу от м. Кызыл-каир, 8 VI 1935, длина 208 мм.) у Прорвы, о. Дурнева, в заливах Мертвый Култук и Кайдак).
— Caspialosa caspia kajdakensis Казанчеев, Рыбн. хоз. СССР, 1936, 3 : 25 (в конце июня — начале июля в зал. Мертвый Култук и Кайдак).
— Caspialosa caspia salina Световидов, Тр. Касп. ком. АН СССР, I, 1937 : 192 (нерест, распространение в связи с соленостью); Зоолог. журн., XXII, 4, 1941 : 231.
Распространение, биология. Чумаевская-Световидова, Тр. Касп. ком. АН СССР, II, 1938 : 23 (возраст и темп роста); Клыков, там же : 77, 92 (ход у п-ова Мангышлак); Перцева-Остроумова, Тр. Всесоюзн. Научно-исслед. инст. морск. рыбн. хоз. и океаногр., XVIII, 1951 : 59, рис. 15, 16 (описание икринок и личинок); Xалдинова, там же : 103, рис. 4,5 (нерест, распределение икринок и личинок).

D III—IV 13—16 (13.2), общим числом 17—20 (17.2), А III (IV) 16—20 (17.7), общим числом 19—23 (20.7), sp. br. 61—117 (86.5), vert. S. 49—51 (50.33), caud. vert. 31—35 (33.17). Самая малотычинковая из каспийских форм вида. Тычинки тонкие, более редкие, чем у других форм, длинные, в 1.25—1.5 раза длиннее жаберных лепестков. Зубы более или менее развитые, на нижней челюсти чаще незаметные, реже имеются, на верхней челюсти обычно хорошо заметные на ощупь. Голова сравнительно небольшая и низкая, длина ее 24.0—26.6 (25.4)%, высота 17.3—21.5 (19.6)% длины тела. Глаза довольно большие, 16.3—20.3 (18.5)% длины головы. Грудные плавники сравнительно короткие, 15.1—17.3 (16.1)% длины тела. Тело высокое, наибольшая высота его 22.3—30.5 (26.2)%, наименьшая — 7.1—9.0 (8.0)% длины тела. Спина и голова сверху темные. За жаберной крышкой с каждой стороны всегда по черному пятну, сзади которого иногда на боках тела по ряду таких же пятен числом до 7. Наряду с обычно окрашенными иногда встречаются особи с интенсивно окрашенной верхней частью головы и спиной. Размеры до 24 см, наичаще 18—21 см. Описание дано по 93—102 экз. длиной от 16 до 24, в среднем 20.2 см, собранным в конце мая — в июне 1935 г. в зал. Мертвый Култук и на мелководье перед входом в этот залив. Количество жаберных тычинок по 205 экз. из этих же мест длиной от 16 до 24, в среднем 20.4 см.
Солоноватоводная форма, размножалась в солоноватой воде в море.
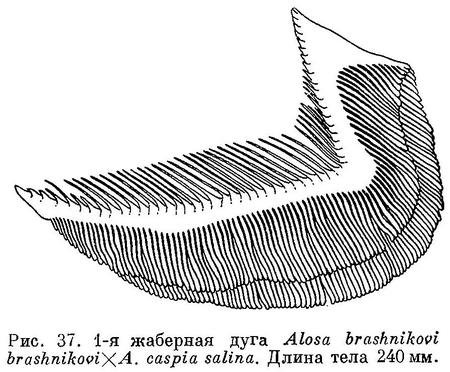
От близкого по внешности Alosa caspia caspia отличается меньшим количеством жаберных тычинок и их строением. На местах нереста нередко встречаются особи, по внешности напоминающие эту форму, но со значительно меньшим количеством жаберных тычинок, и наоборот — внешне похожие на Alosa brashnikovi brashnikovi, но с количеством тычинок значительно большим (до 53 и выше, рис. такой сельди см. на табл. XV).

Распространение довольно широкое. Встречается, насколько известно, в восточной половине Каспийского моря. Наиболее обычен и многочислен весной на местах нереста, которые до обмеления были расположены в восточной половине северной части моря — на мелководье перед входом в зал. Мертвый Култук между о. Дурнева и м. Кызыл-каир к северу от п-ова Бузачи, в зал. Мертвый Култук у м. Кызыл-каир и, повидимому, в районе Прорва — Жилая Коса. Как далеко заходит на запад, не известно. Места нереста в настоящее время достоверно не известны.
Биология изучена недостаточно хорошо. Икрометание в мае—июне в осолоненной или морской воде. Места нахождения в зимнее время не известны, но, как и другие формы вида, зимует, по-видимому, в южной части моря. Равным образом не выяснены и пути миграций к местам нереста, хотя, несомненно, можно предполагать, что они расположены у восточных берегов. Весной в северной части моря у о. Долгого появлялся в половине апреля, при температуре воды ниже 15° ход плохой при повышении ее выше 15°, ход усиливается (Клыков, 1938). В период с 13 по 23 мая 1935 г. в небольшом количестве ловился в Прорве, с конца мая единично, а в июне в значительном количестве ловился у входа в зал. Мертвый Култук. Икринки встречались несколько в более соленой и теплой воде, чем это наблюдается у других форм вида, в зал. Мертвый Култук обнаружены при солености 22.0—32.2‰ и температуре воды 19.2—24.5°. Развитие икринок при температуре воды от 19.2 до 24.5° длится от 39 до 53 час. Предличинки обнаружены на мелководье перед входом в зал. Мертвый Култук и при входе в зал. Кайдак при температуре воды от 17 до 23.6° и солености от 27.6 до 34.9‰ на глубине от 0.75 до 2 м. Личинки найдены на мелководье к востоку от о. Дурнева, по всему зал. Мертвый Култук и в северной части зал. Кайдак. Южная граница массового распространения личинок в зал. Кайдак по западному берегу Кызыл-бай, по восточному — южнее м. Донус-мурун, южнее личинки найдены у входа в зал. Кара-кичу. Личинки встречались при температуре воды от 17 до 21.1° и солености от 21.4 до 39.8‰, у входа в зал. Кара-кичу — при 45.6‰, над глубинами от 0.8 до 5 м (Халдинова, 1951). На местах нереста в июне 1935 г. ловились почти исключительно самки; это обусловлено, повидимому, тем обстоятельством, что наблюдениями был охвачен конец нереста, а вылов производился сетями с крупными ячеями, не задерживавшими более мелких самцов. Выносит значительные осолонения, но все же меньшие, чем долгинка, встречающаяся в самом конце зал. Кайдак при солености 47.7‰; в зал. Кайдак заходит по западному берегу до м. Кызыл-бай (35.8‰) и до м. Донус-мурун (34.1‰) по восточному. Вместе с половозрелыми в северную часть моря заходит и молодь. В июне 1935 г. молодые особи предыдущего года вывода размерами от 87 до 127 мм в особенно большом количестве ловились у входа в зал. Мертвый Култук на мелководье между о. Дурнева и м. Кызыл-каир, южная граница нахождения в зал. Кайдак по восточному берегу несколько южнее мыса Донус-мурун, по западному у Кызыл-бая при содержании солей 21.4—39.8‰. В северной части держится до осени, в сентябре и октябре в районе Прорвы в значительном количестве встречается, по-видимому, эта форма. Половая зрелость наступает по достижении уже двухгодовалого возраста, но на местах нереста преобладают трехгодовалые, двухгодовалые встречаются в незначительном количестве. В 1935 г. возрастной состав на нерестилищах был таков (в процентах):
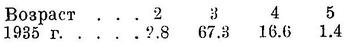
Рост несколько более замедленный, чем у основной формы вида (по исследованиям Чумаевской-Световидовой, 1938):
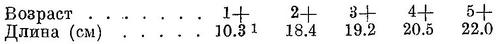
1 Здесь приведены средние размеры возрастных групп, которые близко совпадают с цифрами, полученными обратным расчислением.
Хозяйственное значение небольшое. У форта Шевченко — о. Долгого весной во время хода не ловился. У Прорвы составлял основную массу уловов.
*3d. Alosa caspia knipowitschi (Iljin) — Энзелийский, саринский пузанок (табл. XI).
— Caspialosa knipowitschi Ильин, Сборн. в честь Книповича, 1927 : 69, фиг. (Энзелийский зал.). Тип не указан, но часть экземпляров, по которым дано первоописание, сохранилась. За тип следует считать один из этих экземпляров длиной 212 мм, Энзели, 15 IV 1915, № 31892 коллекций Зоологического института Академии Наук СССР.
— Caspialosa caspia knipowitschi Берг, Рыбы пресных вод, I, 1932 : 89, фиг. 95 (на север до Астары, входит в р. Кумбашинку); Световидов, Тр. Касп. ком. Акад. Наук, I, 1937 : 194, 195 (сравнение с другими формами вида); Зоолог. журн., XXII, 4, 1943 : 231.
— Caspialosa caspia knipowitschi n. saraica Световидов, там же : 231 (по Бергу); Берг, Рыбы пресных вод, I, 1948 : 144 (по n. sarensis Мейснер). Тип не указан. За тип следует считать экземпляр длиной 192 мм, о. Сара, 25 IV 1939, № 32183 коллекций Зоологического института Академии Наук СССР. Отсюда происходят и экземпляры, по которым сделано первоописание.
— Caspialosa caspia knipowitschi Берг, l. с. : 143.
Распространение, биология. Мейснер, Бюлл. Всекасп. научн. рыбохоз. эксп., 5—6, 1932 : 21, фиг. 3 (n. knipowitschi и n. saraica, распространение); Махмудбеков, Зоолог. журн., XXVI, 2, 1947 : 143 (возраст наступления половой зрелости); Дмитриев, там же, XXVI, 6, 1947 : 559 (размеры, возрастной состав, ход у берегов Ирана).
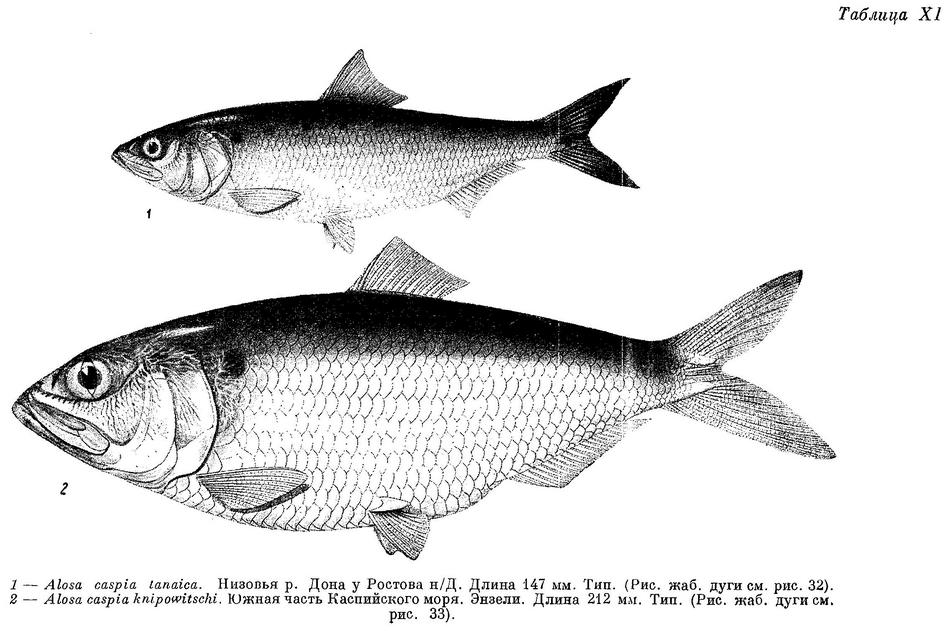
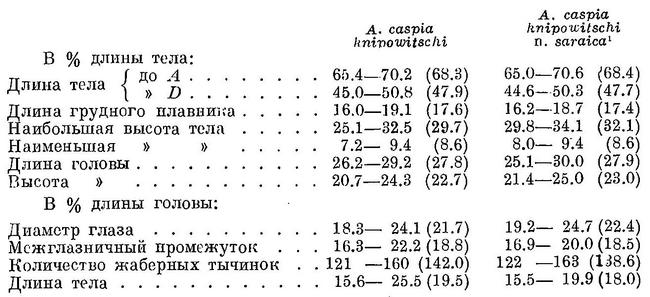
D III—IV 12—14 (13.4), общим числом 16—18 (17.4), A III (IV) 16—20 (18.7), общим числом 19—23 (21.7), sp. br. 121—160 (142.0), vert. S. 47—49 (47.33), caud. vert. 31—33 (31.70). Со срощенными позвонками среди исследованных рыб была одна, имевшая 47 позвонков. Жаберных тычинок наибольшее количество из всех форм вида. Тычинки очень тонкие, нежные, частые и длинные, в 1.5—2 раза длиннее жаберных лепестков. Зубы развиты очень слабо, на нижней челюсти обычно их нет, на верхней едва заметные, или иногда совсем незаметные на ощупь. Голова большая и высокая, длина ее 18.3—24.1 (21.7)%, высота 20.7—24.3 (22.7)% длины тела. Глаза довольно большие, 18.3—24.1 (21.7)% длины головы. Грудные плавники длинные, 16.0—19.1 (17.6)% длины тела. Тело очень высокое, наибольшая высота его 25.1—32.5 (29.7)%, наименьшая — 7.2—9.4 (8.6)% длины тела. Спина и голова сверху темные. За жаберной крышкой сверху с каждой стороны по черному пятну, на боках тела иногда по ряду таких же пятен числом до 7. Размеры до 25.5 см (по Ильину до 29.6 см), наичаще 18—20 см. Описание дано по 42 экз. длиной от 15.6 до 25.5, в среднем 19.5 см, собранным в апреле (26 экз.), июле—августе (15 экз.) и в декабре (1 экз.) 1915 г. в зал. Пехлеви. Это часть тех экземпляров, по которым сделано описание этой формы Б. С. Ильиным. По данным В. И. Мейснера, у n. knipowitschi жаберных тычинок 122—168 (144.7), позвонков 43—49 (47.4), у n. saraica жаберных тычинок 110—180 (145.1), позвонков 45—51 (47.5).
Полупроходная форма, размножается в пресной и слегка осолоненной воде.
От северокаспийских форм вида внешне отличается характерным высоким телом с более длинными плавниками и головой. От астаринского пузанка резко отличается количеством жаберных тычинок. Повидимому, распадается на две формы: Alosa caspia knipowitschi n. knipowitschi (Iljin) — энзелийский пузанок и Alosa caspia knipowitschi n. saraica (Berg) — саринский пузанок. Отличия между этими формами крайне ничтожны, внешне они, подобно формам Alosa caspia caspia, почти не различаются:
1 Описание дано по 50 экз. длиной от 15.5 до 19.9, в среднем 17.9 см, собранным 25—29 IV 1939 в южной части Каспийского моря у о. Сара. Позвонков у них 47—48 (47.69).
У саринского пузанка лишь немного выше тело и больше глаза, причем и по этим признакам обе формы значительно перекрывают одна другую. По исследованиям же В. И. Мейснера, саринский пузанок отличается от энзелийского по другим признакам: большей длиной грудных плавников, более крупными глазами и более длинной головой, причем различия в этих признаках ничтожны и различать по ним обе формы в сущности невозможно. Некоторые различия между этими формами, наблюдаемые в темпе роста, указывают на возможную изоляцию их друг от друга и локализацию по местам икрометания. Возможно, что обе формы являются лишь стадами. Биологически Alosa caspia knipowitschi отличается от Alosa caspia knipowitschi n. saraica, повидимому, так же, как Alosa caspia caspia n. aestuarina от Alosa caspia caspia. Alosa caspia knipowitschi n. saraica и Alosa caspia caspia мечут икру в море, в опресненных частях его, заходят в устья рек, в то время как Alosa caspia knipowitschi и Alosa caspia caspia n. aestuarina размножаются в пресной воде — в дельте Волги и ильменях и в Пехлевийском зал. Эти различия в условиях размножения сказываются в небольших различиях в числе позвонков, что видно из нижеследующего (в скобках приведены данные В. И. Мейснера):

Формы, размножающиеся в море, имеют несколько большее число позвонков, что, судя по всему, обусловлено различиями в температуре воды в период образования позвоночника, повидимому несколько более низкой в море.
Распространение очень узкое, в пределах южной части Каспийского моря, преимущественно в западной его половине, в районе Пехлевийского (Энзелийского) зал., Астары и Бакинского архипелага, осенью, у северных берегов Апшеронского п-ова, откуда вдоль западных берегов в это время заходит до Худата. На восток вдоль южных берегов осенью и зимой в небольшом количестве заходит до Астрабадского зал. К северу от Астары встречается n. saraica. Икрометание типичной формы подвида в Пехлевийском зал. и, возможно, в устье р. Чемхалы (к востоку от р. Сефид-руд), n. saraica нерестует у о. Сара и частично, возможно, в устье р. Кумбашинки.
Биология изучена мало. Нерест в мае—июне в пресной или слегка осолоненной воде. Места пребывания в южной части моря в зимнее время точно не известны. Наряду с Alosa caspia persica одна из самых теплолюбивых форм вида и вообще каспийских сельдей. Основная масса нерестующих рыб у типичной формы состоит из четырех- и пятигодовалых, но встречаются и значительно более младшие, точно так же как и более старшие возрасты, от двух- до девятигодовалых включительно. У n. saraica основную массу нерестующих рыб составляют пятигодовалые, причем и здесь встречаются особи от двух- до восьмигодовалых и даже девяти- и десятигодовалых. Половой зрелости впервые частично достигает в двухгодовалом возрасте, но основная масса, судя по всему, в более старшем (Махмудбеков, 1947). В промысловых уловах в 1932 г., по данным В. И. Мейснера, преобладали пяти- и шестигодовалые, составлявшие свыше 76% улова; позднее преобладали четырех- и пятигодовалые, которые в 1934 г. составляли около 76%, в 1935 г. — около 63% (по наблюдениям А. М. Шуколюкова), в 1945 г. — около 82.2% (Дмитриев, 1947). Темп роста у обеих форм крайне замедленный, энзелийский пузанок, по исследованиям Туркменской научной рыбохозяйственной станции, в первые годы растет несколько быстрее саринского, а затем последний его обгоняет, как это видно из сопоставления средних размеров возрастных групп обеих форм (по данным В. И. Мейснера):

Хозяйственное значение небольшое. Промыслом запасы обеих форм почти не затронуты, о чем свидетельствует высокий возрастной состав уловов и крайне замедленный темп роста. Саринский пузанок вылавливается в большей степени, чем энзелийский, причем вылавливаются более старшие возрастные группы, так как лов производится крупноячейными орудиями лова, благодаря чему, повидимому, и наблюдается увеличение приростов после 5 лет. Ловится у о. Сара, осенью составляет до 30% в уловах Alosa caspia caspia у северных берегов Апшеронского п-ова.
Caspialosa suworowi Berg — Суворовский пузанок.
— Clupea sp. Суворов, Тр. Касп. эксп. 1904 г., I, 1907 : 161.
— Clupeonella suworowi Берг, Мат. позн. русск. рыбол., II, 3, 1913 : 28, табл. XII, фиг. 2; Киселевич, там же, III, 6, 1914 : 24.
— Caspialosa suworowi Берг, там же, IV, 6, 1915 : 6.
— Caspialosa caspia suvorovi Киселевич, Изв. Отд. научно-промысл. исслед., I, 1, 1918 : 17, 18, 29; Киселевич, Каспийско-волжские сельди, I, 1923 : 112, 128—139.
— Caspialosa suworowi Берг, Рыбы пресных вод, 1923 : 498; Киселевич, Промысловые рыбы Волго-каспийского района, Астрах., 1926 : 26.
— Caspialosa caspia suvorovi Аверинцев и Сыч, Тр. Астрах. научн. рыбохоз. ст., VII, 3, 1930 : 9.
— Caspialosa suvorovi Киселевич, Сельди северного Каспия, Сталингр., 1937 : 34.
— Caspialosa suworowi Световидов, Тр. Касп. ком. АН СССР, I, 1937 : 191, табл. I, фиг. 2; Берг, Рыбы пресных вод, I, 1932 : 77; Световидов, Зоолог. журн., XXII, 4, 1943 : 231.
По внешности сходен с Alosa caspia и Alosa saposhnikovi. Жаберных тычинок по Киселевичу 32—76 (55.3), по Бергу 32—68 (55). Зубы развиты слабо или сильно. На нижней челюсти небольшой бугорок, иногда слабо развитый или отсутствующий. Длина головы 22.7—28.0 (25.5)% длины тела, высота головы 17.9—22.0 (19.8)% той же длины. Диаметр глаза 20.0—28.2 (23.4)% длины головы. Высота тела 22.0—29.9 (25.9)% длины его. Длина от 12.9 до 26.8, в среднем 19.1 см. Под этим названием, как показало исследование типа и других экземпляров, хранящихся в коллекциях Зоологического института Академии Наук и определенных К. А. Киселевичем как Caspialosa caspia suvorovi, был описан экземпляр и принимались в дальнейшем помеси между разными видами каспийских сельдей. Экземпляр (№ 15927 коллекций Зоологического института Академии Наук), послуживший Л. С. Бергу для описания этого вида с целью обратить на эту форму внимание будущих исследователей, был первоначально принят Е. К. Суворовым (1907) за помесь, причем им было указано на сходство его с Caspialosa caspia, с одной стороны, и с Caspialosa brashnikovi, с другой. Берг (1913 : 28) сближал Caspialosa suworowi с Caspialosa kessleri auct., а позднее (1915 : 6) с Caspialosa saposhnikovi. Киселевич (1923 : 11) считал этот вид промежуточной формой между Caspialosa caspia и Caspialosa saposhnikovi. К этому же виду отнес К. А. Киселевич и описанную Бергом (1915 : 6) помесь между Caspialosa caspia (вернее одной из ее южнокаспийских форм) и Caspialosa brashnikovi leucocephala (= Alosa brashnikovi grimmi), а также и ряд экземпляров из коллекций Зоологического института Академии Наук (№№ их перечислены в работе Киселевича, 1923 : 112), отнесенных предыдущими исследователями к разным видам (Caspialosa kessleri, Caspialosa saposhnikovi, Caspialosa brashnikovi). Позднее, как мог установить автор, принимая участие в работах Всекаспийской научной рыбохозяйственной экспедиции весной 1931 г. и из бесед с лицами, участвовавшими в сборах и обработке материалов по сельдям этой экспедиции, к Caspialosa suworowi обычно относились такие сельди, в определении которых встречались затруднения. Чаще всего это были экземпляры, промежуточные по своим признакам между обычными видами и формами каспийских сельдей. Условия, благоприятные для образования помесей между видами и формами каспийских сельдей, весьма нередкое явление. Так, например, в конце нереста Alosa brashnikovi brashnikovi на тех же местах начинает метать икру Alosa caspia salina и встречаемые иногда здесь сельди, по внешности напоминающие первую форму, но с большим количеством жаберных тычинок и, наоборот, сельди, внешне напоминающие вторую, но с меньшим количеством тычинок, являются, повидимому, помесями между формами этих двух видов (см. табл. XV и рис. 37). Точно так же на местах икрометания Alosa brashnikovi kisselevitshi в самый разгар нереста бывали находимы единичные самки и самцы Alosa brashnikovi grimmi с еще не вполне выметанными икрой и молоками. Возможности для образования помесей, принимаемых за Caspialosa suworowi, имеются, несомненно, и у других видов и форм сельдей, например у Alosa caspia caspia, Alosa caspia salina и Alosa saposhnikovi. Следует также отметить, что сельди, принимаемые за Caspialosa suworowi, встречаются почти по всему Каспийскому морю, но, как и помесь между Alosa kessleri kessleri и Alosa kessleri volgensis, в небольшом количестве и, что самое существенное, мы до сего времени не располагаем достоверными сведениями об образе жизни этой сельди. В настоящее время достаточно хорошо известны места и условия нереста и строение икринок и личинок в сущности всех сельдей, размножающихся в северной части Каспийского моря, даже таких редко встречаемых, как, например, Alosa sphaerocephala. Мы не располагаем такими сведениями лишь о Caspialosa suworowi, относительно которой имеются крайне неопределенные данные лишь у Киселевича [1923 : 115 — «Икрометание происходит во второй половине апреля и в мае в море у устья Волги» и в такой же неопределенной форме позднее (1937 : 35): «Мечет икру в тех же местах, где и пузанки большеглазый и каспийский, главным образом у устьев Волги и Урала, в одинаковой мере перенося и полное опреснение и слабое осолонение»], без каких-либо указаний, кем и когда наблюдался нерест. Никаких других сведений относительно этого нет, несмотря на то что в отмеченных К. А. Киселевичем местах в последние годы были произведены довольно детальные исследования.
Рыбы пресных вод СССР и сопредельных стран. Л.С.Берг 1948 г.
Caspialosa caspia Eichwald. — Пузанок. Каспийский (среднекаспийский) пузанок
Clupea caspia Eichwald, Bull. Sac. Nat. Moscou, XI, 1838, p. 134 (Каспийское море «к югу»; длина всего тела 190 мм).
среднекаспийский пузанок Суворов, Тр. Касп. эксп. 1904 г., I, 1907, стр. 163 («основной тип»), стр. 143 (западное побережье Каспийского моря от Петровска до Апшеронского полуострова), стр. 168—172 (измерения: Караман [немного севернее Петровска — Махачкалы], Буйнак, Белиджи, Килязи).
? астаринский пузанок Суворов, там же, стр. 146 (от Баку до устья Астары), стр. 164, стр. 178—180 (измерения: Астара и севернее).
Clupeonella caspia Берг, Мат. к позн. русск. рыбол., II, вып, 3, 1913, стр. 36, табл. I, фиг. 1 (Сумгаит), табл. II (близ устья Сулака). — Meйенер, там же, III, вып. 4, 1914, стр. 97 сл., графики VI—XVII (Дагестан; биология). — Киселевич, там же, III, вып. 6, 1914, стр. 14 (северная часть моря: Синее морцо и восточнее), стр. 40, 48—50 (измерения).
Caspialosa caspia Берг, Рыбы пресн. вод России, М., 1916, стр.27. — Мейснер, Раб. Туркменск. рыбохоз. ст., 1936 (нерестится частью в опресненном пространстве северного Каспия, частью входит для нереста в низовья Волги, но, повидимому, не заходит в западные и восточные подстепные ильмени). — Первая научн. конфер. по изуч. сельдевых рыб Касп. моря, 1937, стр. 34 (частью). — Киселевич, Сельди сев. Каспия, 1937, стр. 25. — Перцева, Тр. Инст. морск. рыбн. хоз., VIII, 1939, стр. 27—61 (икра, личинки). — Махмудбеков, Изв. Азербайдж. рыбохоз. ст., III, 1939, стр. 3 (зап. побережье сред. Каспия), стр. 27 (возраст). — Дехтерева, Тр. Инст- морск. рыбн. хоз., XIV, 1940, стр. 68, рис. 11 (сеголетки). — Казакова и Халдинова, там же, стр. 81 (икринки). — Перцева, там же, стр. 109 (нерест). — Чаянова, там же, стр. 211 (питание). — А. А. Остроумов, Тр. Волго-касп. ст., IX, вып. 1, 1947, стр. 3 (миграции).
D III—IV 13—15, А III 16—20, squ. 49—54. Жаберных тычинок на 1-й дуге у половозрелых 70—149, наичаще 115—130, в среднем 122; позвонков 47—52, в среднем (по 2339 экз.) 49.9, наичаще 50 (по данным Туркменской станции). Тычинки тонкие, длинные; длина их в полтора-два раза превосходит длину самых длинных жаберных лепестков. Жаберные дуги тонкие, слабые, легко извлекаемые. Зубы слабые, слабее, чем у всех прочих видов рода Caspialosa, на нижней челюсти: у взрослых обыкновенно совсем нет зубов (у молодых — есть); на верхней— слабые зубы. По наблюдениям Д. Ф. Замахаева, у пузанка с большим количеством жира, что бывает в среднем и южном Каспии, чешуя легко опадает; напротив, в сев. Каспии, куда пузанок приходит уже исхудавшим, а особенно на местах нереста, чешуя держится сравнительно плотно. По данным Т. А. Перцевой, диаметр зрелой , неоплодотворенной икринки 1.11—1.38 мм, диаметр оплодотворенной, вполне разбухшей икринки в среднем 2.1—2.8 мм (пределы колебаний 1.92—2.91), оболочка ее прозрачная, не инкрустированная частицами ила.
Тело умеренно высокое, сильно сжатое с боков, высота его в процентах длины тела (по Суворову — для среднекаспийского, т. е. к северу от Апшеронского полуострова):

1Двухгодовалые, по Суворову.
2Трехгодовалые, по Суворову.
3Четырехгодовалые и выше, по Суворову.
Пузанки к югу от Баку («астаринский пузанок») несколько выше (по Суворову):

Мне попадались особи из среднего Каспия (Буйнак) с высотой до 30%. Глаз большой, диаметр его в процентах к длине тела вышеуказанных групп:
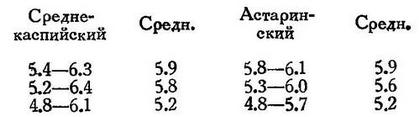
Длина головы в длине тела у тех же групп:

Весною появляется темное пятно за жаберной крышкой; иногда за ним на боках тела 3—7 темных пятен (наличие этих пятен не стоит в связи ни с полом, ни с возрастом). Это мелкая сельдь: вся длина от 160 до 260 мм, иногда до 320 мм. По данным Туркменской станции, средняя длина (до конца средних лучей С) колеблется за отдельные годы между 18 и 22 см, в среднем равняясь 20 см. Средний вес 100—120 г.
Развитие икринок типичной Caspialosa caspia возможно и в пресной и в солоноватой воде. При 18.4° мальки выклевываются через 44—58 часов. Икринки придонные, полупелагические. Икринки Caspialosa caspia отличаются от икринок Caspialosa volgensis меньшим диаметром (у волжской средний диаметр оплодотворенной икринки 3.4 мм, у пузанка 1.9—2.7). Только что выклюнувшиеся личинки имеют в длину 3.9—4.2 мм (у волжской 4.0—4.9 мм); пигментированы они слабее, чем личинки волжской (Перцева, 1939).
Все Каспийское море. Нерестится перед устьями Волги и Урала и частью в дельте Волги, но также в море. По сообщению Т. С. Расса (апрель 1938 г.), икра типичного каспийского пузанка встречается от предустьевых пространств Волги и Урала до Дагестана (южнее исследования не производились), будучи приурочена к глубинам в 2—3 м. В дельте Волги икра типичного пузанка попадается гораздо реже, чем икра ильменного; она была обнаружена преимущественно в низовых районах, но частью в довольно отдаленных ильменях. В море, по данным Т. С. Расса, нерест типичного пузанка во много раз интенсивнее, чем в дельте. Отдельные экземпляры, как исключение, подымаются высоко вверх по Волге; так, в 1869 г. пузанок был добыт у Ярославля, в 1906 г. у Шиловки (выше Сенгилея); тело у обоих высокое, 29—27% (см. Берг, 1913, стр. 38, 40). В небольшом количестве входит в дельту Урала. На зиму каспийский пузанок уходит из сев. Каспия в среднюю и южную части моря, причем в холодные зимы сосредоточивается в районе Красноводской косы. Весною снова направляется для икрометания в северную часть моря — как по западному, так и по восточному берегу. У Дербента он появляется в марте; разгар же лова пузанка здесь бывает в середине и второй половине апреля; заканчивается ход обычно к половине мая. К берегам Мангышлака и к Кулалам он подходит в конце апреля. Икрометание происходит в пресной или сильно опресненной воде сев. Каспия, перед устьями Волги и Урала, частью же в дельте Волги и выше, до Сталинграда. Главные нерестилища расположены в предустьевом пространстве Волги, где глубины 2—3 м; нерест начинается здесь с конца апреля — второй половины мая и продолжается до половины или до конца июня. Часть пузанка нерестится в самой Волге; ход в дельту начинается в конце апреля — начале мая и заканчивается в конце мая — середине июня. После нереста молодь уходит сначала в сев. Каспий, а в августе — сентябре она появляется уже у берегов Дагестана. Каспийский пузанок нерестится, по данным Д. Ф. Замахаева, два-три раза в жизни.
Темп роста, по данным Туркменской станции, таков:
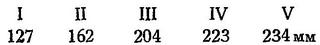
Чем дальше к югу (по западному побережью), тем процент пузанка среди прочих сельдей делается больше, доходя у Апшеронского полуострова до 90%; на севере же, у Сулака, пузанка всего 50%, около 30% Caspialosa saposhnikovi, остальное падает на Caspialosa brashnikovi, Caspialosa volgensis, Caspialosa kessleri.
Пузанок одна из важнейших промысловых рыб Каспия; уловы пузанка составляют около половины уловов всех сельдей Каспия. Из общего улова каспийского пузанка около половины приходится на северный Каспий и около половины на западное побережье среднего Каспия. В среднем за 1936—1939 гг. годовой улов пузанка (включая все формы Caspialosa caspia) составлял 396 тыс. ц, в иные годы превышая 700 тыс. ц.
Caspialosa caspia aestuarina Berg. — Ильменный пузанок
Clupea caspia Арнольд, Тр. Касп. эксп. 1904 г., I, 1907, стр. 226, рис. 3 (зал. Чада в дельте Волги; икрометание, оплодотворение икры).
Clupeonella caspia, Clupeonella caspia m. elongata, ильменный пузанок Берг, Мат. к позн.. русск. рыбол., II, вып. 3, 1913, стр. 38.
Clupeonella caspia Киселевич, там же, IV, вып. 10, 1915, стр. 125 (дельта Волги; биология).
Caspialosa caspia Киселевич, Касп.-волжск. сельди, М., 1923, стр. 108 (частью: северная часть Каспия, дельта Волги), стр. 135 (устья Эмбы), прил., стр. 49—128 (промеры, частью).
Caspialosa caspia aestuarina, севернокаспийский пузанок Берг, Рыбы пресн. вод СССР, I, 1932, стр. 88 (тип из подстепных ильменей дельты Волги). — Мейснер, Раб. Туркменск. рыбохоз. ст., 1936, (по всему Каспию, нерест в подстепных ильменях дельты Волги). — Казанова и Халдинова, Тр. Инст. морск. рыбн. хоз. XIV, 1940, стр. 82 (икрннки).
Caspialosa caspia caspia n. aestuarina Световидов, 1, с., 1944.
Главным отличием этой формы от типичного каспийского пузанка служит, по данным Т. А. Перцевой, И. И. Казановой и Н. А. Халдиновой, меньшая (1.5—1.8 мм) величина оплодотворенных икринок и характер их оболочек: у ильменного пузанка оболочка икринки, плотная (у типичного пузанка она очень тонкая), мутная, инкрустированная частицами ила, с ясно выраженными мелкими капельками липкого секрета. Кроме того, ильменный пузанок отличается от типичной формы более удлиненным телом и меньшим количеством жаберных тычинок. Высота тела в среднем 25—26% длины тела (но попадаются экземпляры и более высокие). Число жаберных тычинок, по данным Туркменской станции, 92—149, наичаще 110—127, в среднем 116.7; с возрастом число тычинок увеличивается. Описание Киселевича (1923) основано главным образом на этой форме. Приводим из его работы (стр. 111) следующие средние данные:

Длина (по Смитту) половозрелых 15—30 см, обычно 19—20 см. Число позвонков у этой формы наичаще 50 (т. е. низкое, как и вообще у всех Caspialosa caspia), с колебаниями от 48 до 52. Кроме темного пятна за жаберной крышкой (тоже обычно отсутствующего), у этого пузанка не бывает пятен на боках.
По данным Туркменской станции, распространен по всему Каспию совместно с типичным пузанком, но держится более или менее самостоятельными косяками. Зимует в южн. Каспии, весною же (апрель—май) движется на север вдоль западного и восточного берегов. Мечет икру в июне, в западных и восточных подстепных ильменях дельты Волги, но также, по сообщению Т. С. Расс, в приверхе дельты и в р. Ахтубе. «Нерест ильменного пузанка в предустьевой зоне, видимо, не происходит; он полностью приурочен к дельте и низовым частям Волги» (Pacс). 2 нюня 1904 г. (н. ст.) И. Н. Арнольд произвел оплодотворение пузанка в ильмене Чада в пресной воде при температуре воды 16 °С, а затем и в морской воде. Развитие совершалось одинаково хорошо как в пресной, так и в соленой воде. Мальки начали выклевываться через 76 часов по оплодотворении (Тр. Касп. эксп. 1904 г., I, 1907 г., стр. 226—227). При 22° С мальки выклевывались уже через 45 часов. Икрометание пузанка наблюдалось в середине июня в ильмене Харбута, где вода совершенно пресная. — Н. С. Строганов (Учен. зап. Моск. унив., XXXIII, 1939, стр. 201—212) наблюдал воздействие воды разной солености на оплодотворенную икру пузанка, взятого из Кардуанской группы подстепных ильменей. Нерест в 1936 г. происходил здесь с 12 по 24 июня. Опыты показали, что развитие икры происходит нормально при соленостях от обычной для речной воды вплоть до 8 pro mille и даже выше. Часть ильменных пузанков задерживается в ильменях после нереста, по спаде воды, и перезимовывает здесь подо льдом до будущего года (Первая научн. конфер. по изуч. сельдевых рыб Касп. моря, 1937, стр. 36.). По сообщению А. А. Клыкова, летом 1936 г. им ловился в больших количествах (500—1000 штук) в Волге, в районе Камышин — Вольск, нерестующий ильменный пузанок (определение Туркменской станции); он шел после черноспинки. В 1937 и 1938 гг. А. В. Лукин обнаружил в затонах Волги у Казани пузанка (№ 28036), возможно, принадлежащего к ильменной форме.
В 1930 г. икра ильменного пузанка из ильменей Дальча, Мечетного и др. была перевезена на аэропланах на Аральское море и выпущена там (Н. Д. Жуковский. Искусственное разведение сельдевых рыб. Тр. Сев. инст. рыбн. хоз., III, 1932.); из нее вывелись пузанки, которые достигли порядочной величины, но затем они, очевидно, все погибли.
Caspialosa caspia salina Svetovidov. — Северо-восточный пузанок
Caspialosa caspia salina Световидов, Доклады Акад. Наук, 1936, I, № 1, стр. 41 (Мертвый култук, Кайдак; представлено в ноябре 1935 г., опубликовано в апреле 1936 г.).
Caspialosa caspia kaidakensis Казанчеев, Рыбн. хоз. СССР, 1936, № 3, стр. 25 (Кайдак; опубликовано летом 1936 г.).
Caspialosa caspia salina Световидов, Тр. Касп. ком. Акад. Наук, I, вып. 1, 1937, стр. 192, табл. II, рис. 3 и 4 (последний — интенсивно пигментированный в черный цвет) (Мертвый култук, Кайдак). — Чумаевская-Световидова, «Заливы Каспийского моря». Рыбы и рыбные промыслы, 1938, стр. 23 (возраст и рост).
D III—IV 13—16, А III 16—20. Жаберных тычинок 61—117, в среднем 86.5. Позвонков 48—52, в среднем 50. Всегда есть зубы на верхней челюсти, иногда и на нижней. Тело высокое, в среднем 26.5% длины тела. Глаза в среднем 5.5% длины тела. За жаберной крышкой темное пятно; иногда за ним до 7 таких же пятен. Длина 16—24, в среднем 20 см (по 446 экз., до конца средних лучей хвостового плавн.). (Световидов). Этот пузанок по числу тычинок наиболее близок к subsp. persica, но отличается от него меньшей высотой тела и большим числом позвонков.
Заливы Каспийского моря Мертвый култук и Кайдак, где нерестится в июне и частью в июле. Где проводит зиму, неизвестно.
Рост северо-восточного пузанка таков (длина по Смитту):

Caspialosa caspia knipowitschi natio saraica Berg, nom. nov. — Бакинский пузанок
Caspialosa caspia knipowitschi n. sarensis Meйснep, Раб. Туркменок, рыбохоз, ст., 1936 (nomen paeoccupatum; тип с о. Сара).
Отличается от Caspialosa caspia knipowitschi несколько меньшей длиной трудных плавников, меньшим глазом и несколько большей головой. Жаберных тычинок наичаще 140—150, в среднем 145 (у 227 экз.). Позвонков 45—51, наичаще 47—48, в среднем 47.5 (у 228 экз.). Длина (по Смитту) до 26 см, в среднем около 20 см.
Юго-западная часть Каспия от Астары до северного побережья Апшеронского полуострова, отдельные особи до Худата (к югу от устья Самура). Нерестится в мае в районе о. Сара в опресненной воде; входит в р. Кумбашинку.
Caspialosa suworowi (Berg). — Суворовский пузанок
Clupea sp. Суворов, Тр. Касп. эксп. 1904 г., I, 1907, стр. 161 (12-футовый рейд).
Clupeonella suworowi Берг, Мат. к позн. русск. рыбол., II, вып. 3, 1913, стр. 28, табл. XII, фиг. 2 (там же, тот же экземпляр). — Киселевич, там же, III, вып. 6, 1914, стр. 24 (северо-восточная часть Каспийского моря).
Caspialosa suworowi Берг, там же, IV, вып. 6, 1915, стр. 6 (северо-восточная часть моря, зал. Кочак, у о. Тюленьего, к югу от Каралата, зал. Кендырли и др.). — Киселевич, Изв. Отд. рыбовод., I, вып. 1, 1918, стр. (18), (29).
Caspialosa caspia suvorovi Киселевич, Касп.-Волжск, сельди, 1923, стр. 112 (северная часть Каспия, дельта Волги: Кривой Бузан).
D III—IV (V) (12) 13—15, А (II) III (IV) 16—19 (20). По внешности сходен с пузанком и с Caspialosa saposhnikovi, помельче первого и крупнее второго: длина от 129 до 268 мм, в среднем 191 мм (до конца средних лучей С). Жаберных тычинок у взрослых обычно 50—55, с колебаниями 32—68, в среднем 55 (у экземпляра-типа 53). По данным Туркменской станции, тычинок от 40 до 83, наичаще 55—62, в среднем 61; тычинки в полтора раза длиннее жаберных лепестков, довольно грубые. Зубы развиты то слабо, то сильно. Тело высокое, высота его 22—30% (в среднем 26%) длины тела. Голова длинная, 22.7—28.0% длины тела (в среднем 25.5). Высота головы 69.5—84.5% (в среднем 78%) ее длины и 17.9—22.0% длины тела. Глаз большой 5.1—7.6% (в среднем 6.0) длины тела и 20.0—23.2% (в среднем 23.4) длины головы. Длина Р 14.5—18.7% (в среднем 16.7%) длины тела. Позвонков 49—52, наичаще 51. Окраска как у пузанка.
Единичными экземплярами заходит в нижнюю часть дельты Волги вместе с пузанком (Caspialosa caspia aestuarina). Еще Гримм добыл эту сельдь в дельте Волги, в Кривом Бузане, у Красного Яра. В Каспийском море обнаружена в северной части, к северу от пареллели Буйнака, главным же образом к востоку от волжского предустьевого пространства и отчасти внутри его. Появляется одновременно с Caspialosa saposhnikovi. Единичными экземплярами встречается по всему Каспию. Икрометание в мае, в море, близ устьев Волги. Рыбаками не отличается от пузанка.
По мнению А. Н. Световидова (Зоол. журн., 1943, стр. 231), под именем Caspialosa suworowi описывались помеси разных видов каспийских сельдей.