Eleginus nawaga (Walbaum, 1792)
Навага, Обыкновенная навага, Северная навага, Ледовитоморская навага, Европейская навага, Беломорская навага, Карская навага, Мезенская навага, Онежская навага, Двинская навага, Югорская навага, Обская навага, Элегинус навага (Rus),
Navaga, Arctic saffron, Atlantic navaga, Arctic cod
(Eng)
Синонимы:
Eleginus navaga (Walbaum, 1792)
Eleginus navaga karaensis Essipov, 1941
Eleginus navaga navaga (Walbaum, 1792)
Eleginus nawaga karaensis Esipov, 1941
Gadus callarias nawaga Walbaum, 1792
Gadus navaga Walbaum, 1792
Gadus nawaga Koelreuter, 1770
Gadus nawaga Walbaum, 1792
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
Навага европейская — Eleginus navaga (Pallas, 1811)
Наваги характеризуются наличием расширений на концах поперечных отростков (ребер) позвонков. Тело как бы сплюснутое (раздавленное) в районе брюшка (в сечении подобно треугольнику). Рот нижний. Усик нормально развит. Боковая линия слабо изогнута. D1 12—16. D2 14—21. D3 20—22. А1 19—23. A2 19—26. Спина и верхняя часть боков тела окрашены в серовато-оливковый цвет, брюхо беловатое. Длина тела порядка 30 см. Обитает в морских и солоноватых водах северных морей до западной части Карского моря на востоке.
Словарь названий пресноводных рыб СССР. Г.У.Линдберг и А.С.Герд 1972 г.
19.3.2. Eleginus navaga (Pallas, 1811) — Навага (Б. : 956).
[1] русск. Голодай Белое м., Голодайка Белое м. — А. Голодная навага Белое м. — Осиновский : 22. Навага — В., 1916 : 474. ~ Навага — Сл. Р. Джемса; Торговый устав 1724 г.; Лепехин, I : 517, IV : 328; Паллас, III, 1 : 40; Начерт. : 389; САР; Сл. 1847 г. [2] укр. Навага — Тат. : 85. [18] англ. Atlantic navaga Канада — Ricker : 73. [39] саам. Navag — Qvigstad : 375.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Северная навага (Eleginus navaga)
распространена в Белом, Печорском (юго-восточная часть Баренцева), Карском
морях, от Восточного Мурмана до Обской
губы. Она обитает вблизи берегов на
небольших глубинах, в преднерестовый
период массами подходит к берегам и
заходит с приливными течениями в реки.
Однако нерест происходит только в
морской воде в декабре — январе подо льдом
на глубине 8—10 м в проливах между
островами, в местах с сильными течениями
и каменистым или песчаным грунтом;
икринки придонные, но не прилипающие
к грунту. Длительность развития икры
до 4 месяцев. Навага достигает длины
до 30—35 см, в Карском море — до 40—42 см. Обычная длина промысловой
беломорской наваги меньше — 15—23 см: это
подошедшая для нереста рыба в возрасте
2—4 лет. В Мезенском заливе и у п-ова Канин ловят навагу в возрасте 6—7 лет,
имеющую длину 16—28 см. Основная
пища наваги — рачки бокоплавы и мизиды,
черви, частично молодь рыб. Навага —
вкусная промысловая рыба, ловят ее рюжами подо льдом во время массового
подхода к берегу для нереста, главным
образом с ноября по январь.
Рыбы северных морей СССР. А.П.Андрияшев 1954 г.
2. Eleginus navaga (Pallas) — Навага (рис. 90).
Gadus navaga Pallas, Zoogr. Rosso-Asiat., III, 1811 : 19 (Сев. Ледовит. океан на восток до устья Оби). — Smitt, Scand. Fish., I, 1893 : 481, fig. 119 (рис. экз. из Архангельска). — Книпович, Определитель, 1926 : 159, фиг. 121.
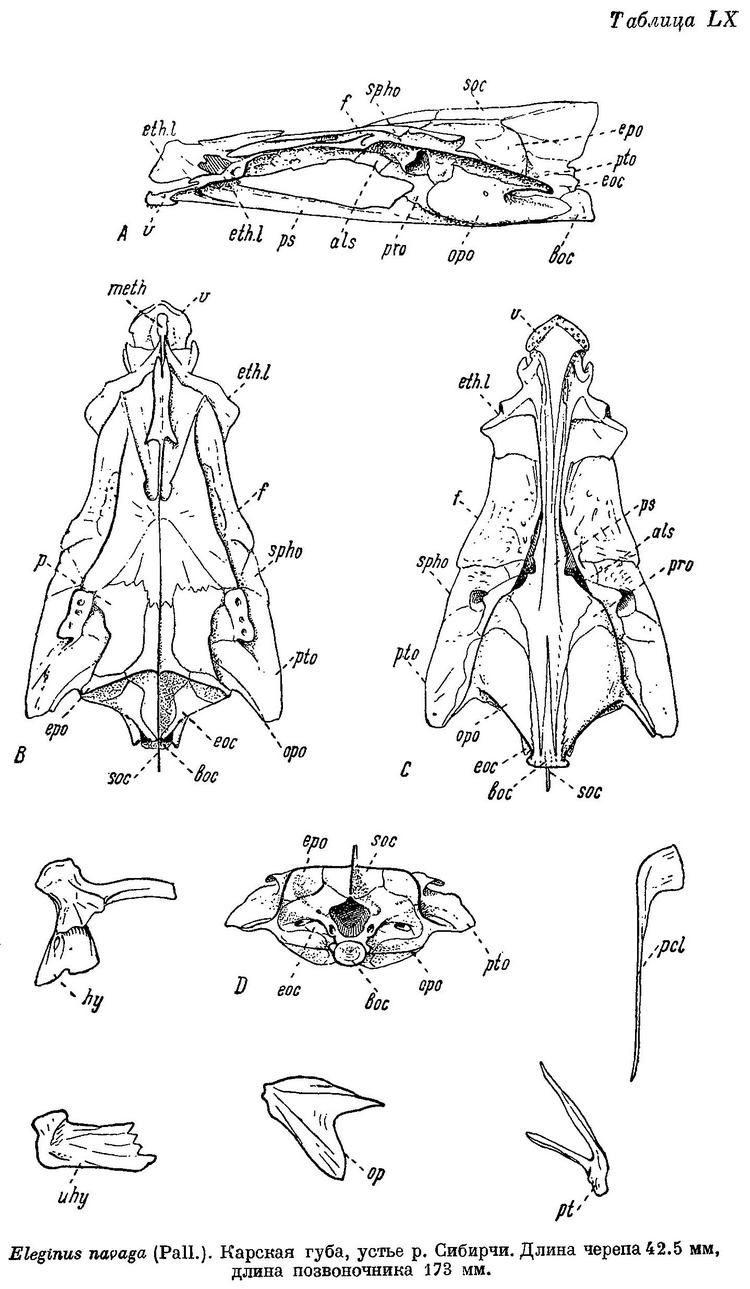
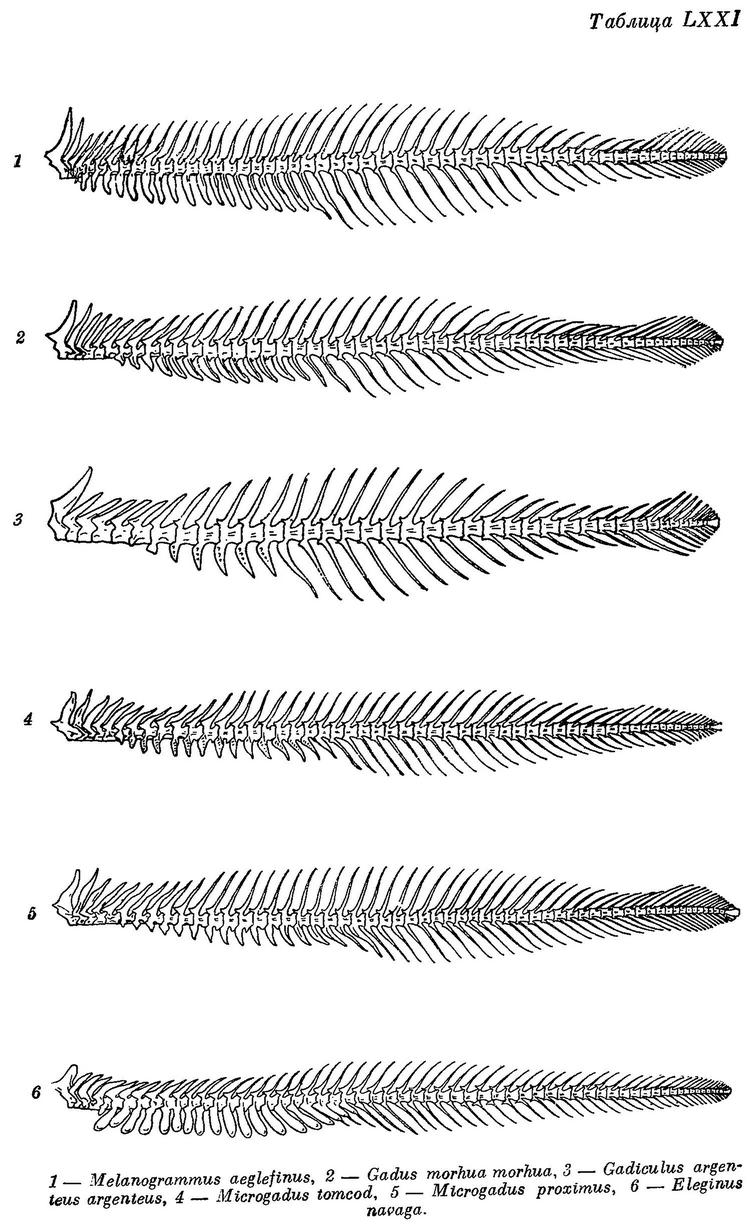
Eleginus navaga Шмидт, Рыбы вост. морей, 1904 : 221 (сравнение с вахней). — Ланшин, Русск. Зоолог. журн., VIII, 4, 1928 : 17 (Мезенск. зал.; биометрика). — Кузьмин-Караваев, Тр. Инст. рыбн. хоз., V, 4, 1930 : 57 (Белое м.; биометрика, возраст и темп роста). — Берг, Рыбы пресн. вод СССР, II, 1933 : 741, фиг. 714. — Световидов, 1948 : 193, фиг: 33—35, табл. XV, LX, LXXI (синонимия; остеология, сводка данных по систематике и биологии). — Клумов и Мантейфель в изд.: Промысл. рыбы СССР, 1949 : 525, атлас табл. 160 (сводка, цветн. рис.).
«navaga» Thielemann, Wiss. Meeresunters. (Helgol.), XIII, 2, 1921 : 213 (навага — это беломорский подвид трески).
Eleginus navaga navaga Есипов, Тр. Научно-иссл. инст. полярн. землед., сер. «Пром. хоз.», 15, 1941 : 144 (Белое м.).
Eleginus navaga karaensis Есипов, 1941 : 145 (Байдарацк. губа, Обск. губа, прол. Малыгина; биометрика).
Распространение, биология. Knipowitsch, Ежег. Зоолог. муз. Акад. Наук, II, 1897 : 150 (от Белого м. и вост. Мурмана до Нов. Земли). — Дерюгин, Фауна Кольск. зал., 1915 : 589 (по данным Мурм. научно-пром. эксп., 1 экз. в Кольск. зал.). — Солдатов, Тр. Плов. морск. научн. инст., 3, 1923 : 63 (горло Белого м., к западу от о. Вайгач, Байдарацк. губа). — Попов, Тр. Ленингр. общ. естеств., LVI, 1,1926 : 49 (беломорская навага отличается от югорской большей высотой тела и большей пятнистостью). — Пробатов, Мат. научно-пром. обслед. Карск. губы, 1934 : 78 (соотношение длины и веса). — Пробатов, Учен. зап. Пермск. унив., II, 3, 1936 : 155 (меристические признаки, рост, питание, плодовитость). — Сахно, За рыбн. индустр. Севера, 1, 1936 : 21; там же, 2, 1936 : 40 (биология, промысл, значение). — Халдинова, Зоолог. журн., XV, 2, 1936 : 321 (Онежск. зал., нерест, развитие). — Бурмакин, Тр. Научно-иссл. инст. полярн. землед., сер. «Пром. хоз.», 10, 1940 : 44 (сев. часть Обск. губы). — Расс, Геогр. параллелизмы в строен. развит. рыб, 1941 : 20 (сведения об икре и личинках). — Мантейфель, Навага Белого моря, изд. Полярн. инст. морск. рыбн. хоз. океаногр., Архангельск, 1945 : 1—44 (промыслово-биологич. очерк). — Алтухов в изд.: Промысл. рыбы Баренц. Белого м., 1952 : 154 (биология, промысел).

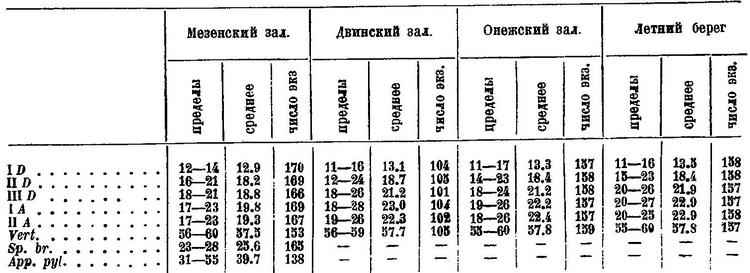
I D 12—16, II D 14—21, III D 20—22, I A 19—23, II A 19—26, P 19—22, позвонков (55) 56—60 (62, 63). Жаберных тычинок 23—29; пилорических придатков 31—55. Формула плавников, по Световидову. В Белом море, по Кузьмину-Караваеву, I D 11—17 (среднее 13.1—13.5), II D 12—24 (18.4—18.7), III D 18—26 (21.2—21.9), I А 18—28 (22.2—23.0), II А 18—26 (22.3—22.9), позвонков 55—60 (57.7—57.8). В Обской губе, по Есипову, I D 11—16 (12.9), II D 15—21 (17.8), III D 17—23 (19.5), I A 17—23 (19.8), II A 18—24 (20.7), позвонков 56—63 (58.9). Сходные с Есиповым данные получены Пробатовым для наваги Карской губы.
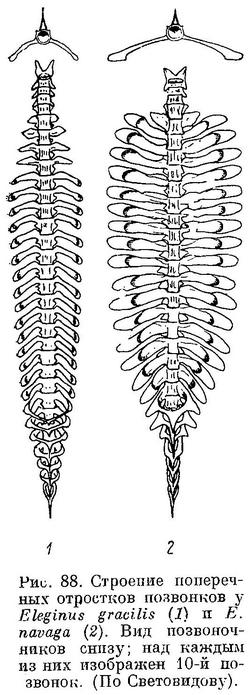
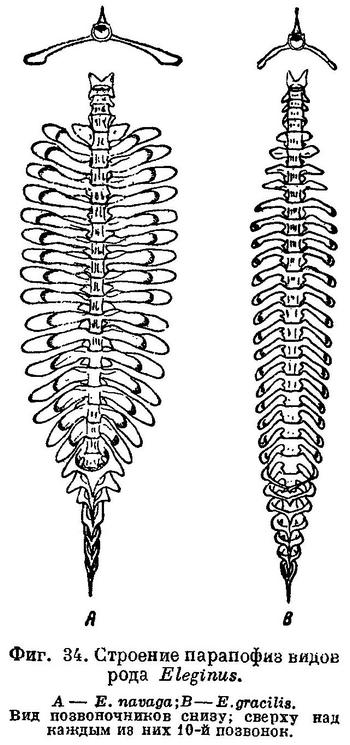
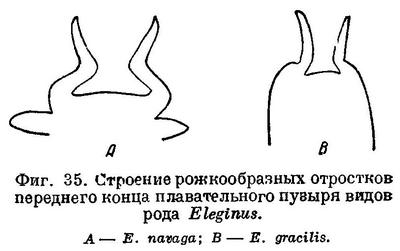
Первые 4 позвонка без поперечных отростков, на 5-м позвонке они короткие и без вздутия; начиная с 6-го позвонка поперечные отростки длинные и булавовидно вздутые (см. рис. 86, 2). Рожкообразные выросты переднего края плавательного пузыря сильно изогнуты. Верх тела буровато-серый с более темными пятнами. На боках окраска светлеет до серебристо-белой на брюхе. Длина до 30—35 см, обычно меньше; в Карском море до 40—42 см, чаще 21—28 см. Самый крупный экземпляр наваги был добыт И. Г. Юдановым (И. Юданов, газ. «Нырьян-Вындер», № 66, от 12 апреля 1944 г.) из р. Индиги в середине марта 1944 г. — посленерестовая («голодная») самка абсолютной длиной 47 см и весом 700 г.
Навага Карского моря и Мезенского залива отличается от беломорской наваги большим числом позвонков (в среднем 57.7—57.9 против 58.1—58.9) и меньшим числом лучей в непарных плавниках. (См. об этом у Есипова (1941) и замечания Световидова (1948 : 196).)
Распространение. От Белого моря на восток до Обской губы. Обычна у берегов вост. Мурмана, на запад единично до Кольского залива.
Биология. Обитает вблизи берегов на небольших глубинах, предпочитает низкие температуры (в зимне-весеннее время отрицательные), постоянно образуя значительные скопления подо льдом; при повышении температуры до 10° интенсивность питания снижается. Часто встречается в сильно опресненных районах, особенно в преднерестовый период, когда навага массами подходит к берегам и даже входит в устья рек. Нерест начинается в декабре (Карская губа) или в январе (Белое море) подо льдом на глубине до 10 м в местах с сильным течением и каменистым или песчаным грунтом. Икрометание порционное. Плодовитость колеблется от 6 до 88 (в среднем около 23) тыс. икринок диаметром 1.5— 1.9 мм. Икринки опускаются на дно или плавают в придонных слоях воды, но во всяком случае не прилипают к грунту. В икрометании в основном участвуют 2—4 (5)-годовалые особи. Рост в Карской губе, по Пробатову:

Навага Двинского залива растет значительно медленнее, достигая в 5 лет всего 16—17 см в длину. В питании наваги отмечены полихеты (Maldanidae, Phyllodocidae), мелкие ракообразные (Amphipoda, Mysidae), мелкие моллюски, икра рыб и рыбы (корюшка, мойва, песчанка, сайка, колюшка; молодь камбал, сигов, наваги, трески, Cottidae).
Eleginus navaga. Сезонные и возрастные изменения в питании беломорской наваги на большом материале изучены 3. Г. Паленичко (Изв. Карело-Финск. фил. АН СССР, 4, 1949 : 52—81). Основной пищей беломорской наваге служат полихеты, донные ракообразные и рыбы. Рыбное питание является наибольшим в период интенсивного откорма наваги (сентябрь, октябрь, март). Полихеты занимают первое место в месяцы с ослабленной интенсивностью питания (ноябрь, декабрь, январь), а также во время посленерестового откорма. Роль ракообразных уменьшается в зимнее время.
Хозяйственное значение имеет в Двинском, Онежском и Мезенском заливах, в Канинско-Чёшском и Печорском районах, в меньшей степени в Карской губе. Уловы, повидимому, не превышают 20—25 тыс. ц. Промышляется поздней осенью и зимой, когда она отличается наилучшими вкусовыми качествами.
Фауна СССР. Рыбы. Том IX, вып.4. Трескообразные. А.Н.Световидов 1948 г.
* 1. Eleginus navaga (Pallas) — Навага (табл. XV, LX, LXXI).
— Piscis e gadorum genere, russis navaga Koelreuter, Nov. Comment. Acad. Petrop., XIV, (1769), 1770 : 484, tabl. XII.
— Gadus navaga Pallas, Zoogr. rosso.-asiat., III, 1841 : 196 (Северный Ледовитый океан, на восток до Оби).
— Eleginus navaga G. Fischer, Mem. Soc. Nat. Moscou, IV, 1812—13 : 252, 257; Ланшин, Русск. зоол. журн., VIII, 4, 1928 : 17 (Мезенский залив, биометрика); Кузьмин-Караваев, Тр. Инст. рыбн. хоз., V, 4, 1930 : 57 (Белое море, систематическая характеристика, возраст, рост); Берг, Рыбы пресных вод, II, 1933 : 741.
— Eleginus navaga navaga Есипов, Тр. Научно-исслед. инст. полярн. землед., животнов. и промысл. хоз., сер. промысл. хоз., 15, 1941 : 44 {Белое море и его заливы).
— Eleginus navaga karaensis Есипов, ibid. : 145 (Карское море: Байдарацкая губа, Обская губа, прол. Малыгина).

Распространение, биология. Smitt, Scand. fish., I, 1893 : 481, f. 119 (Белое море); Knipowitsch, Ежегодн. Зоол. муз. Акад. Наук, II, 1897 : 150, (берега восточной части Мурмана, Белого моря, п-ова Канина, о. Колгуева, Югорского Шара, Новой Земли); Варпаховский, Рыбы басс. Оби, 1902 : 162, f. 11 (северная часть Обской губы); Михайловский, Ежегодн. Зоол. муз. Акад. Наук, VIII, 1903 : 57 (устья рек о. Колгуева); Шмидт, Рыбы восточных морей, 1904 : 221 (Сумский посад, Соловецкие о:ва, р. Индига, Югорский Шар); Дерюгин, Фауна Кольского залива, 1916 : 589 (Кольский залив, случайно); Солдатов, Тр. Плов. морск. научн. инст., III, 1923 : 63 (горло Белого моря, Баренцово море, Байдарацкая губа в Карском море); Ланшин, Русск. зоол. журн., IX, 3, 1929 : 103 (рост, плодовитость); Кузьмин-Караваев, Тр. Инст. рыб. хоз., V, 4, 1930 : 83 (Ивановская губа на Мурмане, мальки); Пробатов, Мат. по научно-пром. обслед. Карской губы и р. Кары, 1934 : 78 (Карская губа, низовья pp. Кары и Сибирчи); Халдинова, Зоол. журн., XV, 2, 1936 : 321 (Онежский залив, биология, места и время нереста, искусственное оплодотворение, литература по нересту); Пробатов, Учен. зап. Пермск. унив., II, 3, 1936 : 155 (биология); Сахно, За рыбн. индустр. Севера, 1936, 1 : 21; ibid., 2 : 40 (биология, промысловое значение); Агапов и Топорков, Пробл. Арктики, 2, 1937 : 107 (у берегов Новой Земли); Бурмакин, Тр. Научно-исслед. инст. полярн. землед., животнов. и промысл. хоз., сер. промысл. хоз., 10, 1940 : 44 (северная часть Обской губы, на юг до устья р. Тамбей, библиография); Мантейфель, Навага Белого моря и ее промысел, изд. Полярн. научно-исслед. инст. морск. рыбн. хоз. и океаногр., 1945 (биология).
I D 12—16, II D 14—21, III D 20—22, I A 19—23, II А 19—26, sp. br. 24—28, vert. 57—60. Количества лучей в плавниках, жаберных тычинок и позвонков у наваги из Мезенского залива (по данным Ланшина) и из Двинского и Онежского заливов и от Летнего берега (по данным Кузьмина-Караваева) таковы (Позвонки считались без последнего, к которому непосредственно присоединяются лучи.):
Без вздутий на концах лишь одна короткая пара парапофиз 5-го позвонка; все остальные парапофизы сильно удлинены, с большими вздутиями на концах (фиг. 34, А). Большие вздутия на каждой стороне на первых 4—5 гемальных дугах. Отростки плавательного пузыря, заходящие во вздутия парапофиз, длинные, рожкообразные выросты у переднего края плавательного пузыря сильно изогнуты (фиг. 35, А). Антедорсальное расстояние 25.8—28.7% длины тела, высота хвостового стебля 3.4—4.1%, длина головы 19.9—21.8% той же длины. В процентах к длине тела: антеанальное расстояние 42.0—46.2, длина грудного плавника 12.1—14.6, длина брюшного плавника 7.1—9.2, высота тела 14.3—17.3, длина хвостового стебля 11.9—13.9. В процентах к длине головы: длина рыла 32.6—37.1, продольный диаметр глаза 12.6—17.4; длина верхней челюсти 35.0—39.1, длина нижней челюсти 42.8—46.7, ширина лба 20.3—26.0. Окраска спины и боков серая, с бурыми или темными пятнами на спине, брюшко серебристо-белое. Обычные размеры в Белом море 15—23 см, но достигает 35 см и более, в Мезенском зал. и у западных берегов п-ова Канина — до 40—42 см. На протяжении своего ареала не везде идентична: например, навага Двинского залива мельче наваги из других беломорских заливов. Повидимому, в отдельных заливах образуются локальные расы или стада.
Распространение. Вдоль северного побережья Европы и Западной Сибири от Кольского залива на западе до Обской губы на востоке, Белое море, вокруг п-ова Канина и о. Колгуева, Чешская губа, у западных берегов Новой Земли (По любезному сообщению И. П. Савватимского, во время его работ у восточных берегов южного острова в августе—сентябре 1935 г. навага здесь не встречалась.) на север до Белушьей губы, в Байдарацкой, Карской губах и в самой северной части Обской губы. В наибольших количествах встречается в заливах Белого моря и в Чешской губе; в Кольском заливе и в северной части Обской губы редка.
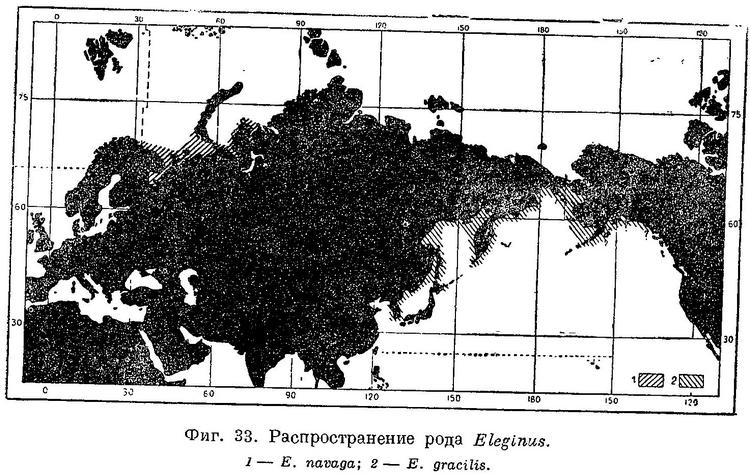
Биология. Холодолюбивая рыба, хотя и не в такой степени, как Boreogadus saida. Больших миграций, повидимому, не совершает. Встречается главным образом на небольших глубинах у берегов с мягкими илистыми грунтами, в открытом море и на глубинах не найдена. В зимнее время встречается в больших количествах подо льдом. Часто ловится в сильно опресненной и даже совершенно пресной воде, заходя в устья и в приливо-отливной зоне реки поднимаясь вверх. Зимой перед икрометанием скопляется в больших количествах у самых берегов, заходя в это время в устья и вверх по течению в реки, в части их, находящиеся под влиянием приливов и отливов. Для нереста навага отходит от берегов и устьев рек на большие глубины (8—10 м) с твердыми песчанистыми и каменистыми грунтами. Нерестилища наваги располагаются в проливах между островами или в углублении между берегом и мелководной банкой, где имеют место сильные приливо-отливные течения. В Белом море навага нерестится в заливах Онежском (около Шуй-острова, Мяг-острова, у мыса Глубокого) и Двинском (в Унской губе, в дельте Двины, у о. Ягры, в районе Сухого моря). Кроме того, известны нерестилища у берегов п-ова Канина, Мурмана (Ивановская губа, 60°22' с.ш., 38°40' в.д.) и, несомненно, имеются везде, где зимой наблюдаются подо льдом массовые скопления наваги. Нерест происходит с начала или середины, а иногда даже с конца января и заканчивается в основном приблизительно дней через 15, хотя отдельные зрелые особи встречаются и позднее. Плодовитость наваги исчисляется от 6 до 90 тыс. икринок. Икринки наваги не липкие, не являются пелагическими, а опускаются на дно. Указание на липкость ее (Вебель, За рыбн. индустр. Севера, 1935, 5 : 24) оспаривается (Сахно, l. c.). Развитие икринок происходит только в морской воде, в пресной они погибают. Указания на нерест наваги в реках (Пробатов, l. с.) ошибочны. Выход личинок из икры при температуре в среднем +2° С наблюдался на 39-й день, в естественных условиях при температуре около —1° С инкубационный период длится, повидимому, около 4 месяцев. Половой зрелости навага достигает на 3—4-м году. Основная масса мечущей икру наваги состоит из 2—4-годовалых рыб, размеры которых, по данным Кузьмина-Караваева, в это время (январь) таковы (в см):

Наиболее мелкими размерами и медленным ростом отличается навага в районе устья Сев. Двины, где она к 5 годам достигает 16—17 см против 20—25 см в других районах. Более крупных размеров и большего возраста достигает навага в Мезенском зал. и у западного побережья п-ова Канина, где основная масса ее ловится в возрасте 6—7 лет при длине 16—28 см. Основной пищей являются амфиподы, мизиды, черви, частично питается мелкой рыбой (корюшкой, мойвой, песчанкой, сайкой, мелкими треской, камбалой и навагой), которая большее значение имеет в питании крупной наваги. Летом при повышении температуры воды до 10° С и выше питается слабо.
Xозяйственное значение довольно велико. Главными местами лова являются Двинский, Онежский и Мезенский заливы Белого моря и Чешская губа. Промысловый лов происходит во время массового скопления у берегов подо льдом для нереста. В промысловом количестве начинает ловиться с ноября, наиболее интенсивный лов совпадает с разгаром нереста — в январе, после чего начинает ловиться «голодная», отнерестившаяся навага и уловы быстро падают. Полных и точных статистических сведений об уловах наваги нет. Лов производится, главным образом, у берегов Карело-Финской ССР, в Онежском, Двинском и Мезенском заливах, в Чешской губе и в Печорском районе. Общий улов с 1898 по 1944 гг. колебался от 1.6 (1920 г.) до 24.0 (1930 г.) тыс. ц (Мантейфель, l. c.), по некоторым указаниям (Скворцов, Бюлл. рыбн. хоз., 1926, 1: 16) достигал в прежние годы свыше 40 тыс. ц. Наиболее низкие уловы были в 1920—1923 гг., в последующие годы они опять возросли и достигли прежнего уровня. Повышение уловов произошло, главным образом, за счет развития промысла в Чешской губе и в Печорском районе. В прежние годы в наибольшем количестве навага ловилась в Мезенском зал. и по западному берегу п-ова Канина, но в последние годы количество ее здесь сильно уменьшилось. Главным орудием лова являются рюжи и отчасти уда.
Сравнительные заметки. Eleginus navaga karaensis, описанная В. К. Есиповым по экземплярам из Обской губы и выделенная им в подвид на основании сравнения с экземплярами из Белого моря, исследованными Г. А. Кузьминым-Караваевым, отличается от последних меньшим количеством лучей во 2-м и 3-м спинных и в обоих анальных плавниках. В остальных сравниваемых признаках (количества позвонков и лучей в 1-м спинном плавнике) существенных различий нет. Судя по меньшему количеству лучей в указанных плавниках у исследованных В. К. Есиповым экземпляров, можно полагать, что отличие их от беломорских обусловлено тем, что при просчете лучей кожные покровы при начале и конце этих плавников им не подрезались и скрытые в них лучи оказались не просчитанными (Соблюдалась ли эта необходимая предосторожность В. К. Есипов не указывает. Г. А. Кузьмин-Караваев кожные покровы удалял.). Как указывалось выше, без подрезания кожных покровов правильно количество лучей можно подсчитать только в 1-м спинном плавнике, по количеству лучей в котором экземпляры В. К. Есипова существенно не отличаются от экземпляров Г. А. Кузьмина-Караваева. Все подвиды тресковых (например Lota lota, Molva dipterygia, Trisopterus minutus, Gadus morhua и др.) значительно различаются краниологически, в частности шириной черепа. Таких различий В. К. Есиповым не указано; их, судя по всему, у наваги и нет. Несомненно, что навага, как указывалось выше, распадается на ряд более мелких таксономически, чем подвид, форм. Выделять эти формы следует с соблюдением единообразной методики просчетов и измерений, одним лицом, воздерживаясь от использования для сравнения данных, полученных другими исследователями.
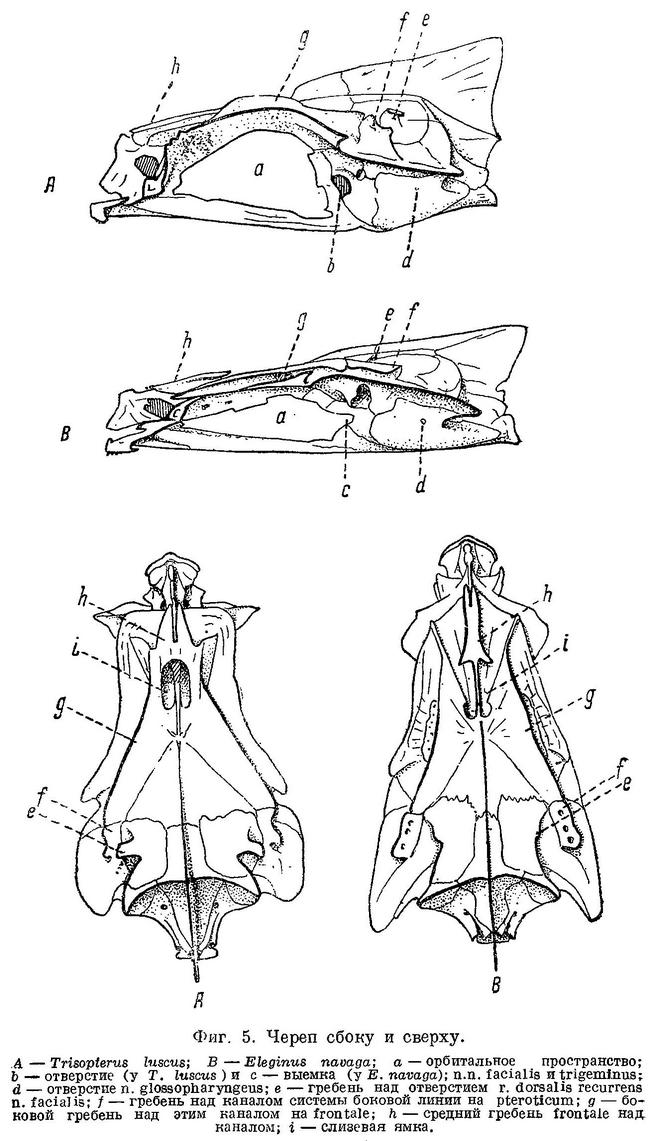
N. facialis выходит из черепа у тресковых вместе с n. trigeminus через отверстие или различной глубины выемку в передней части prooticum сбоку черепа (фиг. 5,b,с). У Brosme brosme нерв этот выходит через отверстие, а у следующего за ним по ряду Gaidropsarus mediterraneus — через глубокую выемку в prooticum, края которой настолько сближены, что почти замыкают ее спереди. У Ciliata mustela и далее следующих за ним в ряду рыб выемка становится спереди более широкой и менее глубокой. У Lota lota выемка так же глубока и края спереди сужены почти так же, как у Gaidropsarus mediterraneus, а у всех видов Molva и Merluccius n. facialis из черепа выходит опять через отверстие. Через отверстие n. facialis выходит и у следующих далее в ряду видов рода Trisopterus. При этом ширина костной перемычки у Trisopterus luscus несколько шире, чем у Trisopterus minutus, а у последней шире, чем у Trisopterus esmarkii. Odontogadus merlangus и далее следующие за ним представители семейства имеют в prooticum выемку, которая последовательно становится спереди более широкой и менее глубокой. У обеих форм Gadiculus argenteus выемки почти нет. Далее, у видов родов Eleginus, Microgadus, Boreogadus, Arctogadus и Theragra выемка постепенно углубляется, а края ее становятся более сближенными. Micromesistius имеет сбоку черепа отверстие. Таким образом, по этому признаку ряд тресковых, расположенных по степени дифференциации их непарных плавников, разделяется в свою очередь на два параллельных ряда, соответственно делению семейства на подсемейства. Такой же закономерности подчиняется изменение и некоторых других признаков.
Сверху черепа тресковых на каждом parietale расположено по отверстию, через которое выходит r. dorsalis recurrens n. facialis (фиг. 5,е и рис. В на табл. XXI—LXVII). У одних тресковых каждое из этих отверстий прикрыто у каждого вида в различной степени развитыми гребнями; у других гребень не развит, и отверстие сверху открыто. У Brosme brosme и далее следующих за ним в ряду Gaidropsarus mediterraneus, Ciliata mustela, Enchelyopus cimbrius гребни, прикрывающие эти отверстия, имеются. У Raniceps ranius и всех видов родов Phycis и Urophycis отверстия r. dorsalis recurrens n. facialis не прикрыты, так как гребни у них имеют вид едва заметных выростов, расположенных несколько сзади отверстий. У Lota, всех видов Molva и Merluccius гребни развиты и прикрывают отверстия. В пределах подсемейства Gadinae гребни на parietalia развиты сильнее. У всех видов родов Trisopterus, Odontogadus, Pollachius, Gadus и Melanogrammus гребни развиты, размеры их уменьшаются от Trisopterus к Melanogrammus. Нет гребней у Gadiculus. Далее гребень есть, постепенно увеличиваясь к концу ряда, у Eleginus, Microgadus, Boreogadus, Arctogadus, Theragra и Micromesistius, причем у одних особей Eleginus navaga гребни есть, у других они не развиты.
N. glossopharyngeus (Stensiö, 1921 : 155) выходит из черепа через небольшое отверстие на opisthoticum (фиг. 5,d и рис. А на табл.XXI—LXVII). Величина этого отверстия, изменяясь в пределах ряда, подчиняется той же закономерности. У Brosme brosme отверстие почти не заметно. У Gaidropsarus mediterraneus, Ciliata mustela, Enchelyopus cimbrius, Raniceps raninus оно небольшое, у видов родов Phycis и Uropkycis несколько больше, у Lota и Molva большое и у Merluccius вновь небольших размеров. Далее, у всех видов рода Trisopterus отверстие едва заметно, размеры его постепенно увеличиваются до Gadus и Melanogrammus и, начиная с Gadiculus, уменьшаются вновь.
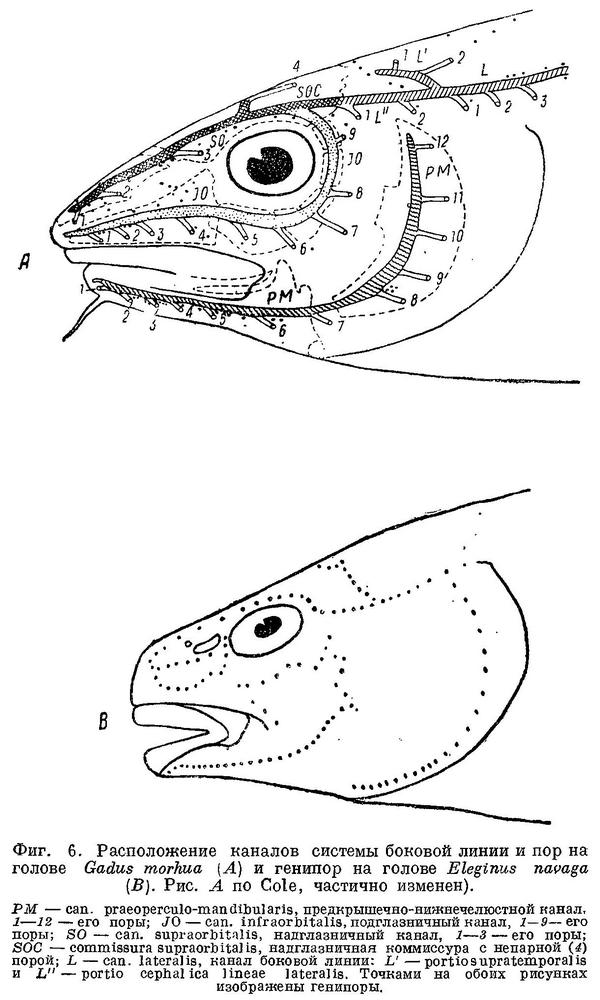
Боковая линия у тресковых образует на голове с каждой стороны каналы (фиг. 6): предкрышечно-нижнечелюстной, подглазничный и надглазничный с короткой комиссурой, соединяющей правый и левый каналы. Сама боковая линия имеет на голове надвисочную часть (L '), над которой образуется окостенение из 4—5 косточек ossa tabularia (= supratemporale). Каждый канал системы боковой линии на голове прикрыт сверху почти на всем своем протяжении гребнем тех костей, на которых эти каналы расположены (frontale, pteroticum, sub- и praeorbitale, nasale, praeoperculum, tabulare). Среди этих гребней наиболее существенное значение имеет средний гребень frontale (фиг. 5, h) и боковые гребни frontale и pteroticum (фиг. 5, f и g). Межглазничная комиссура надглазничного канала расположена в особом углублении frontale — слизевой ямке (фиг. 5, i). Слизевая ямка у одних представителей Gadidae спереди замкнута сросшимися краями боковых передних гребней над каналами системы боковой линии на frontale, у других широко открыта, так как края боковых и средних гребней frontalia укорочены и не срастаются впереди слизевой ямки. У Brosme brosme края боковых и средних гребней frontalia настолько сближены, что почти замыкают спереди небольших размеров слизевую ямку. У Gaidropsarus mediterraneus, Ciliata mustela и Enchelyopis cimbrius слизевая ямка спереди открыта, очень небольших размеров. Raniceps raninus имеет слизевую ямку спереди открытой. У одних видов родов Phycis и Urophycis слизевая ямка спереди открыта, у других закрыта; оба эти рода представляют в этом отношении небольшое отклонение от общего правила, о причинах чего будет сказано ниже. У Lota lota слизевая ямка спереди почти замкнута сближенными краями боковых и средних гребней frontalia, которые у видов рода Molva, срастаясь, образуют хорошо замкнутую ямку. Слизевая ямка у всех видов рода Merluccius представляет собой обширное углубление посередине frontalia и имеет несколько уклоняющийся от обычного для тресковых вид. Слизевая ямка замкнута далее, начиная с Trisopterus и кончая Gadiculus. У всех далее следующих в ряду видов слизевая ямка спереди открыта и замкнута вновь у стоящих в конце его Theragra chalcogramma и Micromesistius poutassou и Micromesistius australis. При этом следует отметить, что закрытая или открытая спереди слизевая ямка обычно свойственна всем видам любого рода тресковых, за исключением, как указывалось выше, родов Phycis и Urophycis. Кроме того, есть один вид — Eleginus navaga, — у которого сближенные края боковых и средних гребней frontalia иногда бывают сращены с одной стороны, а на одном из исследованных черепов и с обеих сторон. Таким образом, эта особенность находится в неустойчивом состоянии в первом случае в пределах рода, во вторых — в пределах вида, стоящих в середине каждого из параллельных рядов.
Подобная же закономерность наблюдается и в строении гребней над самыми каналами боковой линии на frontalia. У Brosme, Gaidropsarus, Ciliata, Enchelyopus, Molva, Trisopterus, Micromesistius, т. е. у рыб, стоящих в начале и конце каждого ряда, края гребней на frontalia тесно примыкают к этим костям, почти замыкая канал. Более того — у некоторых из этих рыб (Trisopterus) края гребней местами соединяются с frontalia при помощи костных перемычек. У всех остальных расположенных в ряду между этими рыбами тресковых края гребней над слизевыми каналами на frontalia обычно приподняты. Несколько уклоняется в этом признаке род Merluccius, у которого гребни развиты очень слабо.
Вдоль каждого из каналов системы боковой линии на голове расположены многочисленные группы нервных окончаний (фиг. 6, В) — генипор (pit-organs), или каналы эти открываются наружу рядом пор (фиг. 6, А), причем единичные генипоры имеются и в этом случае. Боковая линия на теле у тресковых тянется от головы до хвостового плавника в виде непрерывной или прерывистой, разделенной небольшими промежутками, линии. У Brosme brosme боковая линия тянется непрерывной полосой от головы до конца спинного плавника, становясь прерывистой лишь на хвостовом стебле. У Gaidropsarus mediterraneus, Ciliata mustela и Enchelyopus cimbrius боковая линия прерывиста на всем протяжении, у Raniceps raninus ее, повидимому, нет совершенно. У видов родов Phycis и Urophycis и у Lota lota боковая линия непрерывна почти до конца анального плавника, далее прерывиста; у видов рода Moiva она прерывиста сзади от конца анального плавника, у видов Merluccius — непрерывна на всем протяжении. У видов родов Trisopterus, Odontogadus, Pollachius и Melanogrammus боковая линия непрерывна на всем протяжении; у видов родов Gadus, Gadiculus (Экземпляры Gadiculus обычно бывают без чешуй, и поэтому о строении их боковой линии судить трудно, однако до середины третьего спинного плавника она тянется без перерывов.) и Microgadus она прерывиста сзади от середины или конца третьего спинного плавника, у видов рода Eleginus прерывиста уже сзади от начала второго спинного плавника и у видов родов Boreogadus и Arctogadus боковая линия прерывиста на всем протяжении. Далее, у Theragra chalcogramma боковая линия прерывиста лишь сзади от начала второго спинного плавника и у Micromesistius poutassou и Micromesistius australis без перерывов на всем протяжении.
Рыбы пресных вод СССР и сопредельных стран. Л.С.Берг 1948 г.
1. Eleginus navaga (Pallas). — Навага
Piscis e gadorum genere, russis nawaga Koelreuter, Nov. Comment. Acad. Petrop., XIV (1769), pt. I, 1770, p. 484 , pl. XII (местонахождение не указано).
Gadus navaga Pallas, Zoogr. rosso-asiat., III, 1811, p. 196 (Сев. Ледовитый океан на восток до устьев Оби). — Smitt, Scand. fishes, I, 1892, p. 481, fig. 119 (Белое море). — Knipowitsch, Ежегодн. Зоол. муз. Акад. Наук, II, 1897, стр. 150 (восточная часть Мурмана, Белое море, берега Канина, Колгуева, Новой Земли, Югорский шар). — Варпаховский, Рыбы басс. Оби, 1902, стр. 162, рис. 11 (северная часть Обской губы). — Михайловский, Ежегодн. Зоол. муз. Акад. Наук, VIII, 1903, стр. 57 (устья рек о. Колгуева).
Eleginus navaga Шмидт, Рыбы Вост. морей, 1904, стр. 221 (Сумский посад, Соловецкие острова, р. Индига, Югорский шар).
навага Н. Смирнов, Изв. Арханг. общ. изучения Русск. Сев., 1910, № 2 (Мезенский и Печорский уезды от устья р. Койды до Индиги).
Eleginus navaga Солдатов, Тр. Плов. морск. инст., III, 1923, стр. 63 (Баренцево море). — Ланшин, Русск. зоол. журн., VIII, № 4, 1928, стр. 17 (Мезенский залив; биометрика). — Кузьмин-Караваев, Тр. Инст. рыбн. хоз., V, вып. 4, 1930, стр. 57 (Белое море; биометрика, возраст, рост); стр. 83 (мальки в Ивановской губе, 38° 40' в. д., на Мурмане). — Пробатов, Матер. по научно-пром. обслед. Карской губы, М., 1934, стр. 78 (Карская губа, молодь в реках Каре и Сибирче не выше 5 км от устья, где вода солоноватая; вся длина до 395 мм, вес до 487 г); Уч. зап Пермск. унив., II, вып. 3, 1936, стр. 155—173 (там же; возраст, рост, плодовитость). — Сахно, За рыбную индустрию Севера, 1936, № 1, стр. 21 (Поморский берег Белого моря; уловы); № 2, стр. 40 (биология). — Халдинова, Зоол. журн., XV, 1936, стр. 321 (Поморский берег, о. Сосновец; биология). — Бурмакин, Тр. Инст. полярн. землед., вып. 10, 1940, стр. 44 (Обская губа на юг до устья р. Тамбей — 71.5° с. ш.).
Eleginus navaga karaensis Есипов, Тр. Инст. полярн. землед., вып. 15, 1941, стр. 141—145 (Байдарацкая губа, Обская губа, прол. Малыгина).
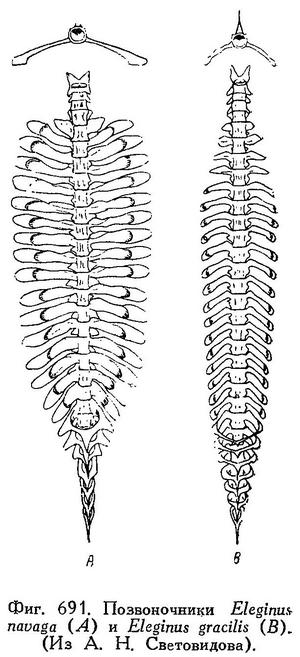
Eleginus navaga Б. Мантейфель, Навага Белого моря и ее промысел. Архангельск, 1945, 44 стр. (биология, уловы). — Световидов, Трескообразные, 1948, стр. 193, фиг. 33 (карта), 34 (парапофизы), 3) (плавательный пузырь), табл. XV, фиг 1, табл. LX (череп), табл. LXXI, фиг. 6 (позвоночник).
I D 12—14, средн. 13, II D 16—21, средн. 18, III D 18—21, средн. 19, I А 17—23, средн. 20, II А 17—23, средн. 19, Р 19—22, средн. 20, жаберных тычинок на 1-й дуге 23—28, средн. 26, позвонков, считая с последним, 57—61, средн. 58.5, пилорич. придатков 31—54, средн. 40.
Эти данные по экземплярам из Мезенского залива: число лучей у 166—170, тычинок у 165, позвонков у 153, пилорич. придатков у 138 (Ланшин; число позвонков увеличено мною на 1, так как Ланшин не считал позвонка с hypuralia). — У 25 экз. западного берега Белого моря (Колежма) позвонков 57—59, средн. 57.9 (по подсчету К. Ф. Телегина). По данным Г. А. Кузьмина-Караваева:
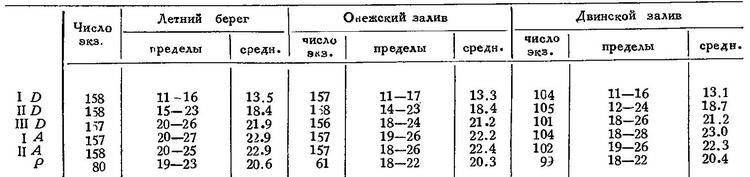
Объединяя крайние величины, получим: I D 11—17, II D 12—24, III D 18—26, I А 18—28, II А 18—26, Р 18—23. По сравнению с мезенской навагой получаются значительные расхождения, особенно в III D, I А и II А.
Число позвонков у беломорских, по Кузьмину-Караваеву, с последним, следующее:

Поперечные отростки позвонков, начиная с 6-го, удлинены, на концах вздуты; в этих вздутиях заключены ответвления плавательного пузыря; 5-й позвонок несет короткие поперечные отростки без вздутий. Рожкообразные выросты у переднего края плавательного пузыря короткие, сильно изогнутые. В процентах к абсолютной длине тела: длина головы 19.3—25.3 (средн. 22.3), наибольшая высота тела 12.7—19.6 (16.1), наименьшая высота тела 3.2—4.7 (4.0), длина рыла 6.6—8.9 (7.7), ширина лба 4.8—7.1 (5.8), антедорсальное расстояние (до I D) 26.1—33.0 (29.7), антеанальное расстояние (до I А) 40.1—43.0 (43.7), длина I D 9.0—13.9 (11.0), длина II D 13.0—18.8 (16.4), длина II А 12.6—17.4 (15.0). В процентах к длине головы: длина рыла 31.0—33.8 (34.4), горизонтальный диаметр глаза 14.5—21.6 (17.8), заглазье 44.9—52.3 (48.5), ширина лба 22.2—32.0 (25.9), длина верхнечелюстной 33.3—41.7 (37.3), длина нижнечелюстной 40.5—50.0 (45.5). В процентах к ширине лба горизонтальный диаметр глаза 50.0—88.9 (69.0) (по 170 экз. из Мезенского залива, 85 самцов, 81 самка, половых различий нет, длина тела не приведена; Ланшин). На спине бурые пятна.
Длина 210—315 мм, в среднем 248 мм. Самая крупная навага из Белого моря имела в длину 37 см. На зап. Ямале, в устье р. Юрибея А. Н. Пробатов наблюдал 19 марта 1946 г. громадных наваг: недавно выметавшая икру длиною 441 мм (абс.) весила 703 г, другая длиною 419 мм, весом 713 г, имела текучую икру диаметром 1.8 мм.
Берега Мурмана от Кольского залива (К. Дерюгин, Зап. Акад. Наук, XXXIV, № 1, 1915, стр. 589, единично.) на восток (в промысловых количествах только начиная с восточной части Мурмана), Белое море и на восток вплоть до Обской губы; Колгуев, Новая Земля по западному берегу. Перед икрометанием (недели за две) заходит на некоторое время в устья рек, но, повидимому, не поднимается выше области влияния прилива.
Навага — прибрежная арктическая рыба. В открытом море она почти не встречается. В с. Колемжа на Поморском берегу Белого моря в 1933 г. разгар нереста приходился на 5—20 января (Ранее, до эпохи потепления, нерест происходил в декабре (Мантейфель, стр. 16).); нерест закончился в общем в конце января, но отдельные особи с текучими половыми продуктами попадались в феврале и марте. Икра у наваги донная не липкая; в пресной воде она не развивается. Нерест происходит при солености не менее 27‰ (П. Сахво). Плодовитость у 109 самок из Карской губы в среднем 45 665 икринок (Пробатов). Массовый выход личинок в конце марта — начале апреля. Нерестилища находятся обычно в местах с сильными приливо-отливными течениями и с песчано-каменистым или галечным грунтом. В дельте Сев. Двины, между островами, таких мест много. Промысел наваги в Белом море производится со времени ледостава и до начала февраля. Основную массу промышляемой здесь наваги составляют особи, которым исполнилось два или три полных года, а более старые особи встречаются единицами. Средний вес беломорской наваги 50—100 г; самая мелкая навага — двинская (Кузьмин-Караваев). Средний вес карской наваги 197 г (366 экз.).
Возраст беломорской наваги, по данным Кузьмина-Караваева, таков:

3 Нерестующие особи.
Наблюденная средняя длина до конца средних лучей хвостового плавника в миллиметрах для тех же проб:
На Поморском берегу Белого моря возраст, рост и вес наваги таковы (3200 экз., П. Сахно):

Самок 54.5%, самцов 45.5%.
Относительно подвида karaensis А. Н. Световидов считает, что указываемое для этого подвида меньшее число лучей в II D и III D в I А и II А объясняется тем, что при подсчете лучей кожные покровы при начале и конце этих плавников не подрезались и скрытые в них лучи не просчитывались.