Царство Бактерии, Настоящие бактерии, Эубактерии (Bacteria Woese et al., 1990), Regnum Bacteria Woese et al., 1990 (Bacterias, Bacteriums, Eubacterias, True bacterias) 25 типов
Материал подготовил Фактистов И. Е., 2020 г. Сайт www.fishbiosystem.ru
Надтипы и типы:
• Тип Актинобактерии (Actinobacteria)
• Тип Aquificae
• Надтип группа Bacteroidetes/Chlorobi
• Тип Caldiserica
• Надтип группа Chlamydiae/Verrucomicrobia
• Тип Chloroflexi
• Тип Chrysiogenetes
• Тип Цианобактерии (Cyanobacteria)
• Тип Deferribacteraceae
• Тип Deinococcus-Thermus
• Тип Dictyoglomi
• Тип Elusimicroba
• Надтип группа Fibrobacteres/Acidobacteria
• Тип Фирмикуты (Firmicutes)
• Тип Фузобактерии (Fusobacteria)
• Тип Gemmatimonadetes
• Тип Nitrospirae
• Тип Planctomycetes
• Тип Протеобактерии (Proteobacteria)
• Тип Спирохеты (Spirochaetes)
• Тип Synergistetes
• Тип Тенерикуты (Tenericutes)
• Тип Thermodesulfobacteria
• Тип Thermotogae
Бактерии (эубактерии (Eubacteria), др.-греч. палочка) — домен (надцарство) прокариотных (безъядерных) микроорганизмов, чаще всего одноклеточных. К настоящему времени описано около десяти тысяч видов бактерий и предполагается, что их существует свыше миллиона, однако само применение понятия вида к бактериям сопряжено с рядом трудностей.
Изучением бактерий занимается раздел микробиологии — бактериология.
Термин.
До конца 1970-х годов термин «бактерия» был синонимом прокариотов, но в 1977 году на основании данных молекулярной биологии прокариоты были разделены на домены архебактерий и эубактерий. Впоследствии, чтобы подчеркнуть различия между ними, они были переименованы в архей и бактерий соответственно. Хотя до сих пор под бактериями часто понимают всех прокариотов, в данной статье описаны лишь эубактерии. Однако, эти две группы схожи, и многие положения статьи справедливы также для архей — в подобных случаях используется термин «прокариоты» или сочетание «бактерии и археи».
В экологических и микробиоценотических исследованиях под бактериями часто понимают лишь нефотосинтезирующие немицелиальные прокариоты, противопоставляя их по функциям актиномицетам и цианобактериям.
История изучения.
Впервые бактерии увидел в оптический микроскоп и описал в 1676 году голландский натуралист Антони ван Левенгук. Как и всех микроскопических существ, он назвал их «анималькули».
Название «бактерии» ввёл в употребление в 1828 году Христиан Эренберг.
В 1850-х годах Луи Пастер положил начало изучению физиологии и метаболизма бактерий, а также открыл их болезнетворные свойства.
Дальнейшее развитие медицинская микробиология получила в трудах Роберта Коха, которым были сформулированы общие принципы определения возбудителя болезни (постулаты Коха). В 1905 году он был удостоен Нобелевской премии за исследования туберкулёза.
Основы общей микробиологии и изучения роли бактерий в природе заложили М. В. Бейеринк и С. Н. Виноградский.
Изучение строения бактериальной клетки началось с изобретением электронного микроскопа в 1930-е. В 1937 году Э. Чаттон предложил делить все организмы по типу клеточного строения на прокариот и эукариот, и в 1961 году Стейниер и Ван Ниль окончательно оформили это разделение. Развитие молекулярной биологии привело к открытию в 1977 году К. Вёзе коренных различий и среди самих прокариот: между бактериями и археями.
Строение.
Подавляющее большинство бактерий (за исключением актиномицетов и нитчатых цианобактерий) одноклеточны. По форме клеток они могут быть округлыми (кокки), палочковидными (бациллы, клостридии, псевдомонады), извитыми (вибрионы, спириллы, спирохеты), реже — звёздчатыми, тетраэдрическими, кубическими, C- или O-образными. Формой определяются такие способности бактерий, как прикрепление к поверхности, подвижность, поглощение питательных веществ. Отмечено, например, что олиготрофы, то есть бактерии, живущие при низком содержании питательных веществ в среде, стремятся увеличить отношение поверхности к объёму, например, с помощью образования выростов (т. н. простек).
Из обязательных клеточных структур выделяют три:
• нуклеоид
• рибосомы
• цитоплазматическая мембрана (ЦПМ)
С внешней стороны от ЦПМ находятся несколько слоёв (клеточная стенка, капсула, слизистый чехол), называемых клеточной оболочкой, а также поверхностные структуры (жгутики, ворсинки). ЦПМ и цитоплазму объединяют вместе в понятие протопласт.
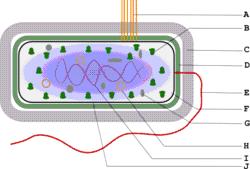
Схема строения грамположительной бактерии: A — пили, B — рибосомы, C — капсула, D — слой пептидогликана, E — жгутик, F — цитозоль, G — запасные вещества, H — плазмида, I — нуклеоид, J — цитоплазматическая мембрана.
Строение протопласта.
ЦПМ ограничивает содержимое клетки (цитоплазму) от внешней среды. Гомогенная фракция цитоплазмы, содержащая набор растворимых РНК, белков, продуктов и субстратов метаболических реакций, названа цитозолем. Другая часть цитоплазмы представлена различными структурными элементами.
Одним из основных отличий клетки бактерий от клетки эукариот является отсутствие ядерной мембраны и, строго говоря, отсутствие вообще внутрицитоплазматических мембран, не являющихся производными ЦПМ. Однако у разных групп прокариот (особенно часто у грамположительных бактерий) имеются локальные впячивания ЦПМ, выполняющие в клетке разнообразные функции и разделяющие её на функционально различные части. У многих фотосинтезирующих бактерий существует развитая сеть производных от ЦПМ фотосинтетических мембран. У пурпурных бактерий они сохранили связь с ЦПМ, легко обнаруживаемую на срезах под электронным микроскопом, у цианобактерий эта связь либо трудно обнаруживается, либо утрачена в процессе эволюции. В зависимости от условий и возраста культуры фотосинтетические мембраны образуют различные структуры — везикулы, хроматофоры, тилакоиды.
Вся необходимая для жизнедеятельности бактерий генетическая информация содержится в одной ДНК (бактериальная хромосома), чаще всего имеющей форму ковалентно замкнутого кольца (линейные хромосомы обнаружены у Streptomyces и Borrelia). Она в одной точке прикреплена к ЦПМ и помещается в структуре, обособленной, но не отделённой мембраной от цитоплазмы, и называемой нуклеоид. ДНК в развёрнутом состоянии имеет длину более 1 мм. Бактериальная хромосома представлена обычно в единственном экземпляре, то есть практически все прокариоты гаплоидны, хотя в определённых условиях одна клетка может содержать несколько копий своей хромосомы, а Burkholderia cepacia имеет три разных кольцевых хромосомы (длиной 3,6; 3,2 и 1,1 млн пар нуклеотидов). Рибосомы прокариот также отличны от таковых у эукариот и имеют константу седиментации 70 S (80 S у эукариот).
Помимо этих структур, в цитоплазме также могут находиться включения запасных веществ.
Клеточная оболочка и поверхностные структуры.
Клеточная стенка — важный структурный элемент бактериальной клетки, однако необязательный. Искусственным путём были получены формы с частично или полностью отсутствующей клеточной стенкой (L-формы), которые могли существовать в благоприятных условиях, однако иногда утрачивали способность к делению. Известна также группа природных не содержащих клеточной стенки бактерий — микоплазм.
У бактерий существует два основных типа строения клеточной стенки, свойственных грамположительным и грамотрицательным видам.
Клеточная стенка грамположительных бактерий представляет собой гомогенный слой толщиной 20—80 нм, построенный в основном из пептидогликана с меньшим количеством тейхоевых кислот и небольшим количеством полисахаридов, белков и липидов (так называемый липополисахарид). В клеточной стенке имеются поры диаметром 1—6 нм, которые делают её проницаемой для ряда молекул.
У грамотрицательных бактерий пептидогликановый слой неплотно прилегает к ЦПМ и имеет толщину лишь 2—3 нм. Он окружён наружной мембраной, имеющей, как правило, неровную, искривлённую форму. Между ЦПМ, слоем пептидогликана и внешней мембраной имеется пространство, называемое периплазматическим, и заполненное раствором, включающим в себя транспортные белки и ферменты.
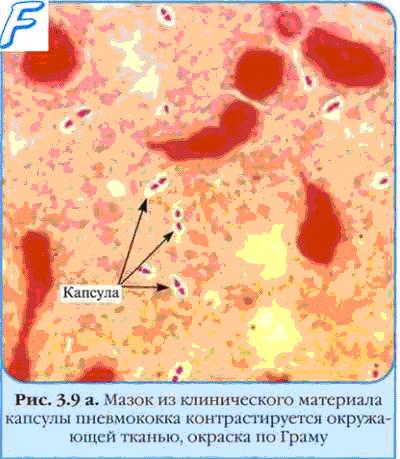
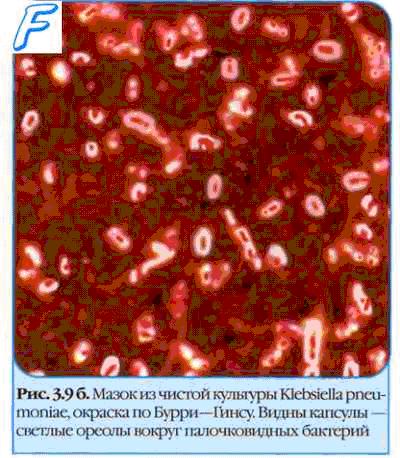
С внешней стороны от клеточной стенки может находиться капсула — аморфный слой, сохраняющий связь со стенкой. Слизистые слои не имеют связи с клеткой и легко отделяются, чехлы же не аморфны, а имеют тонкую структуру. Однако между этими тремя идеализированными случаями есть множество переходных форм.
Бактериальных жгутиков может быть от 0 до 1000. Возможны как варианты расположения одного жгутика у одного полюса (монополярный монотрих), пучка жгутиков у одного (монополярный перитрих или лофотрихиальное жгутикование) или двух полюсов (биполярный перитрих или амфитрихиальное жгутикование), так и многочисленные жгутики по всей поверхности клетки (перитрих). Толщина жгутика составляет 10—20 нм, длина — 3—15 мкм. Его вращение осуществляется против часовой стрелки с частотой 40—60 об/с.
Помимо жгутиков, среди поверхностных структур бактерий необходимо назвать ворсинки. Они тоньше жгутиков (диаметр 5—10 нм, длина до 2 мкм) и необходимы для прикрепления бактерии к субстрату, принимают участие в транспорте метаболитов, а особые ворсинки — F-пили, нитевидные образования, более тонкие и короткие (3—10 нм х 0,3—10 мкм), чем жгутики, — необходимы клетке-донору для передачи реципиенту ДНК при конъюгации.
Размеры.
Размеры бактерий в среднем составляют 0,5—5 мкм. Escherichia coli, например, имеет размеры 0,3—1 на 1—6 мкм, Staphylococcus aureus — диаметр 0,5—1 мкм, Bacillus subtilis — 0,75 на 2—3 мкм. Крупнейшей из известных бактерий является Thiomargarita namibiensis, достигающая размера в 750 мкм (0,75 мм). Второй является Epulopiscium fishelsoni, имеющая диаметр 80 мкм и длину до 700 мкм и обитающая в пищеварительном тракте хирурговой рыбы Acanthurus nigrofuscus. Achromatium oxaliferum достигает размеров 33 на 100 мкм, Beggiatoa alba — 10 на 50 мкм. Спирохеты могут вырастать в длину до 250 мкм при толщине 0,7 мкм. В то же время к бактериям относятся самые мелкие из имеющих клеточное строение организмов. Mycoplasma mycoides имеет размеры 0,1—0,25 мкм, что соответствует размеру крупных вирусов, например, табачной мозаики, коровьей оспы или гриппа. По теоретическим подсчётам, сферическая клетка диаметром менее 0,15—0,20 мкм становится неспособной к самостоятельному воспроизведению, поскольку в ней физически не могут поместиться все необходимые биополимеры и структуры в достаточном количестве.
При линейном увеличении радиуса клетки её поверхность возрастает пропорционально квадрату радиуса, а объём — пропорционально кубу, поэтому у мелких организмов отношение поверхности к объёму выше, чем у более крупных, что означает для первых более активный обмен веществ с окружающей средой. Метаболическая активность, измеренная по разным показателям, на единицу биомассы у мелких форм выше, чем у крупных. Поэтому небольшие даже для микроорганизмов размеры дают бактериям и археям преимущества в скорости роста и размножения по сравнению с более сложноорганизованными эукариотами и определяют их важную экологическую роль.
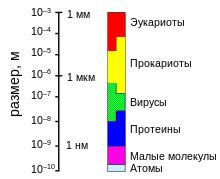
Шкала относительных размеров эукариот, прокариот, вирусов, протеинов, молекул и атомов.
Многоклеточность у бактерий.
Одноклеточные формы способны осуществлять все функции, присущие организму, независимо от соседних клеток. Многие одноклеточные прокариоты склонны к образованию клеточных агрегатов, часто скреплённых выделяемой ими слизью, эти агрегаты получили название биоплёнки. Чаще всего это лишь случайное объединение отдельных организмов, но в ряде случаев временное объединение связано с осуществлением определённой функции, например, формирование плодовых тел миксобактериями делает возможным развитие цист, при том что единичные клетки не способны их образовывать. Подобные явления наряду с образованием одноклеточными эубактериями морфологически и функционально дифференцированных клеток — необходимые предпосылки для возникновения у них истинной многоклеточности.
Многоклеточный организм должен отвечать следующим условиям:
• его клетки должны быть агрегированы,
• между клетками должно осуществляться разделение функций,
• между агрегированными клетками должны устанавливаться устойчивые специфические контакты.
Многоклеточность у прокариот известна, наиболее высокоорганизованные многоклеточные организмы принадлежат к группам цианобактерий и актиномицетов. У нитчатых цианобактерий описаны структуры в клеточной стенке, обеспечивающие контакт двух соседних клеток — микроплазмодесмы. Показана возможность обмена между клетками веществом (красителем) и энергией (электрической составляющей трансмембранного потенциала). Некоторые из нитчатых цианобактерий содержат помимо обычных вегетативных клеток функционально дифференцированные: акинеты и гетероцисты. Последние осуществляют фиксацию азота и интенсивно обмениваются метаболитами с вегетативными клетками.
Способы передвижения и раздражимость.
Многие бактерии подвижны. Имеется несколько принципиально различных типов движения бактерий. Наиболее распространено движение при помощи жгутиков: одиночных бактерий и бактериальных ассоциаций (роение). Частным случаем этого также является движение спирохет, которые извиваются благодаря аксиальным нитям, близким по строению к жгутикам, но расположенным в периплазме. Другим типом движения является скольжение бактерий, не имеющих жгутиков, по поверхности твёрдых сред и движение в воде безжгутиковых бактерий рода Synechococcus. Его механизм пока недостаточно изучен; предполагается участие в нём выделения слизи (проталкивание клетки) и находящихся в клеточной стенке фибриллярных нитей, вызывающих «бегущую волну» по поверхности клетки. Наконец, бактерии могут всплывать и погружаться в жидкости, меняя свою плотность, наполняя газами или опустошая аэросомы.
Бактерии активно передвигаются в направлении, определяемом теми или иными раздражителями. Это явление получило название таксис. Различают хемотаксис, аэротаксис, фототаксис.
Метаболизм.
Конструктивный метаболизм.
За исключением некоторых специфических моментов биохимические пути, по которым осуществляется синтез белков, жиров, углеводов и нуклеотидов, у бактерий схожи с таковыми у других организмов. Однако по числу возможных вариантов этих путей и, соответственно, по степени зависимости от поступления органических веществ извне они различаются.
Часть из них может синтезировать все необходимые им органические молекулы из неорганических соединений (автотрофы), другие же требуют готовых органических соединений, которые они способны лишь трансформировать (гетеротрофы).
Наибольшей степенью гетеротрофности отличаются внутриклеточные паразиты. Если при этом они способны существовать на богатых искусственных средах, они называются факультативными (факультативными иногда также называют паразитов, способных проделывать весь свой жизненный цикл во внешней среде, без участия хозяина). Некоторые облигатные (обязательные) внутриклеточные паразиты утратили часть путей биосинтеза и получают многие органические вещества, вплоть до АТФ, из клеток хозяина. Велика степень зависимости от хозяев также многих бактерий-эндосимбионтов. Большинство бактерий принадлежит к сапрофитам: они не питаются непосредственно веществами других организмов, но используют синтезированные ими органические вещества после их смерти. Существует также ряд бактерий, требующих наличия в среде небольшого круга определённых органических веществ (аминокислот, витаминов), которых они не могут синтезировать самостоятельно и, наконец, гетеротрофы, которые нуждаются лишь в одном довольно низкомолекулярном источнике углерода (сахар, спирт, кислота). Некоторые из них отличаются высокой специализацией (Bacillus fastidiosus может использовать только мочевую кислоту), другие в качестве единственного источника углерода и энергии могут использовать сотни различных соединений (многие Pseudomonas).
Удовлетворять потребности в азоте бактерии могут как за счёт его органических соединений (подобно гетеротрофным эукариотам), так и за счёт молекулярного азота (как и некоторые археи). Большинство бактерий используют для синтеза аминокислот и других азотсодержащих органических веществ неорганические соединения азота: аммиак (поступающий в клетки в виде ионов аммония), нитриты и нитраты (которые предварительно восстанавливаются до ионов аммония). Фосфор они способны усваивать в виде фосфата, серу — в виде сульфата или реже сульфида.
Энергетический метаболизм.
Способы же получения энергии у бактерий отличаются своеобразием. Существует три вида получения энергии (и все три известны у бактерий): брожение, дыхание и фотосинтез.
Брожение — серия окислительно-восстановительных реакций, в ходе которых образуются нестабильные молекулы, с которых остаток фосфорной кислоты переносится на АДФ с образованием АТФ (субстратное фосфорилирование). При этом возможно внутримолекулярное окисление и восстановление.
Дыхание — окисление восстановленных соединений с переносом электрона через локализованную в мембране дыхательную электронтранспортную цепь, создающую трансмембранный градиент протонов, при использовании которого синтезируется АТФ (окислительное фосфорилирование). В то время как эукариоты в конечном итоге почти всегда «сбрасывают» электрон на кислород (лишь в редких случаях акцептором электронов могут служить нитраты), бактерии могут использовать вместо него окисленные органические и минеральные соединения (фумарат, углекислый газ, сульфат-анион, нитрат-анион и др.; см. анаэробное дыхание), а вместо окисляемого органического субстрата использовать минеральный (водород, аммиак, сероводород и др.), что часто бывает сопряжено с автотрофной фиксацией CO2 (см. хемосинтез).
Фотосинтез бактерий может быть двух типов — бескислородный, с использованием бактериохлорофилла (зелёные, пурпурные и гелиобактерии) и кислородный с использованием хлорофилла (цианобактерии (хлорофилл a), прохлорофиты (a и b)). Цианобактерии, глаукоцистофитовые, красные и криптофитовые водоросли — единственные фотосинтезирующие организмы, содержащие фикобилипротеины. У архей встречается бесхлорофилльный фотосинтез с участием бактериородопсина (правда, энергия света используется при этом не для фиксации CO2, а непосредственно для синтеза АТФ, так что в строгом смысле это не фотосинтез, а фотофосфорилирование).
Бактерии, осуществляющие только бескислородный фотосинтез, не имеют фотосистемы II. Во-первых, это пурпурные и зелёные нитчатые бактерии, у которых функционирует только циклический путь переноса электронов, направленный на создание трансмембранного протонного градиента, за счёт которого синтезируется АТФ (фотофосфорилирование), а также восстанавливается НАД(Ф)+, использующийся для ассимиляции CO2. Во-вторых, это зелёные серные и гелиобактерии, имеющие и циклический, и нециклический транспорт электронов, что делает возможным прямое восстановление НАД(Ф)+. В качестве донора электрона, заполняющего «вакансию» в молекуле пигмента в бескислородном фотосинтезе используются восстановленные соединения серы (молекулярная, сероводород, сульфит) или молекулярный водород.
Существуют также бактерии с весьма специфическим энергетическим метаболизмом. Так, в октябре 2008 года в журнале Science появилось сообщение об обнаружении экосистемы, состоящей из представителей одного единственного ранее неизвестного вида бактерии — Desulforudis audaxviator, которые получают энергию для своей жизнедеятельности из химических реакций с участием водорода, образующегося в результате распада молекул воды под воздействием радиации залегающих вблизи нахождения колонии бактерий урановых руд. Некоторые колонии бактерий, обитающие на дне океана, используют для передачи энергии своим собратьям электрический ток.
Типы жизни.
Объединить типы конструктивного и энергетического метаболизма можно в следующей таблице:
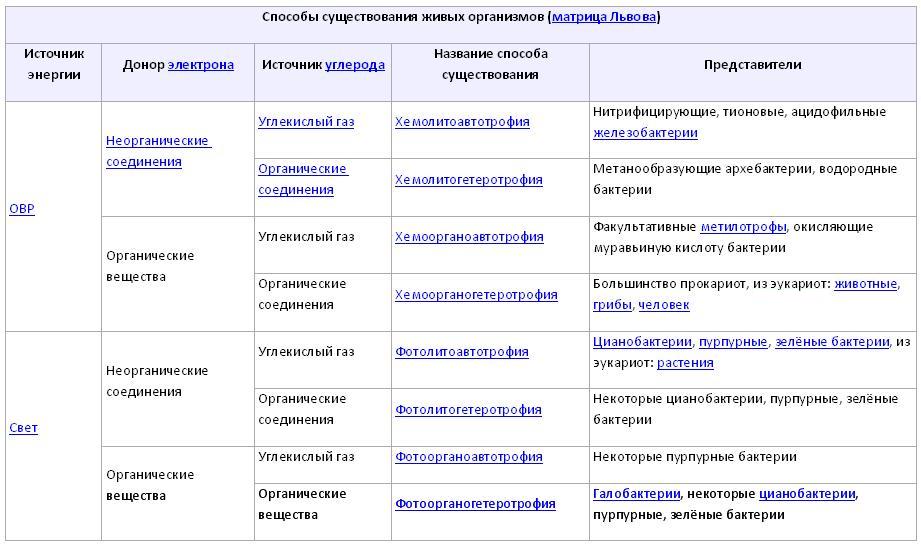
Из таблицы видно, что разнообразие типов питания прокариот гораздо больше, чем у эукариот (последние способны лишь к хемоорганогетеротрофии и фотолитоавтотрофии).
Размножение бактерий.
Некоторые бактерии не имеют полового процесса и размножаются лишь равновеликим бинарным поперечным делением или почкованием. Для одной группы одноклеточных цианобактерий описано множественное деление (ряд быстрых последовательных бинарных делений, приводящий к образованию от 4 до 1024 новых клеток). Для обеспечения необходимой для эволюции и приспособления к изменчивой окружающей среде пластичности генотипа у них существуют иные механизмы.
При делении большинство грамположительных бактерий и нитчатых цианобактерий синтезируют поперечную перегородку от периферии к центру при участии мезосом. Грамотрицательные бактерии делятся путём перетяжки: на месте деления обнаруживается постепенно увеличивающееся искривление ЦПМ и клеточной стенки внутрь. При почковании на одном из полюсов материнской клетки формируется и растёт почка, материнская клетка проявляет признаки старения и обычно не может дать более 4 дочерних. Почкование имеется у разных групп бактерий и, предположительно, возникало несколько раз в процессе эволюции.
У других бактерий кроме размножения наблюдается половой процесс, но в самой примитивной форме. Половой процесс бактерий отличается от полового процесса эукариот тем, что у бактерий не образуются гаметы и не происходит слияния клеток. Однако главнейшее событие полового процесса, а именно обмен генетическим материалом, происходит и в этом случае. Это называется генетической рекомбинацией. Часть ДНК (очень редко вся ДНК) клетки-донора переносится в клетку-реципиент, ДНК которой генетически отличается от ДНК донора. При этом перенесённая ДНК замещает часть ДНК реципиента. В процессе замещения ДНК участвуют ферменты, расщепляющие и вновь соединяющие цепи ДНК. При этом образуется ДНК, которая содержит гены обеих родительских клеток. Такую ДНК называют рекомбинантной. У потомства, или рекомбинантов, наблюдается заметное разнообразие признаков, вызванное смещением генов. Такое разнообразие признаков очень важно для эволюции и является главным преимуществом полового процесса.
Известны 3 способа получения рекомбинантов. Это — в порядке их открытия — трансформация, конъюгация и трансдукция.
Генетический аппарат.
Гены, необходимые для жизнедеятельности и определяющие видовую специфичность, расположены у бактерий чаще всего в единственной ковалентно замкнутой молекуле ДНК — хромосоме (иногда для обозначения бактериальных хромосом, чтобы подчеркнуть их отличия от эукариотических, используют термин генофор (англ. genophore)). Область, где локализована хромосома, называется нуклеоид и не окружена мембраной. В связи с этим новосинтезированная мРНК сразу доступна для связывания с рибосомами, а транскрипция и трансляция сопряжены.
Отдельная клетка может содержать лишь 80 % от суммы генов, имеющихся во всех штаммах её вида (т. н. «коллективный геном»).
Помимо хромосомы, в клетках бактерий часто находятся плазмиды — также замкнутые в кольцо ДНК, способные к независимой репликации. Они могут быть настолько велики, что становятся неотличимы от хромосомы, но содержат дополнительные гены, необходимые лишь в специфических условиях. Специальные механизмы распределения обеспечивают сохранение плазмиды в дочерних клетках так что они теряются с частотой менее 10-7 в пересчёте на клеточный цикл. Специфичность плазмид может быть весьма разнообразной: от присутствия лишь у одного вида-хозяина до плазмиды RP4, встречающейся почти у всех грамотрицательных бактерий. В плазмидах кодируются механизмы устойчивости к антибиотикам, разрушения специфических веществ и т. д., nif-гены, необходимые для азотфиксации также находятся в плазмидах. Ген плазмиды может включаться в хромосому с частотой около 10-4 — 10-7.
В ДНК бактерий, как и в ДНК других организмов, выделяются транспозоны — мобильные сегменты, способные перемещаться из одной части хромосомы к другой, или во внехромосомные ДНК. В отличие от плазмид, они неспособны к автономной репликации и содержат IS-сегменты — участки, которые кодируют свой перенос внутри клетки. IS-сегмент может выступать в роли отдельной транспозоны.
Горизонтальный перенос генов.
У прокариот может происходить частичное объединение геномов. При конъюгации клетка-донор в ходе непосредственного контакта передаёт клетке-реципиенту часть своего генома (в некоторых случаях весь). Участки ДНК донора могут обмениваться на гомологичные участки ДНК реципиента. Вероятность такого обмена значима только для бактерий одного вида.
Аналогично бактериальная клетка может поглощать и свободно находящуюся в среде ДНК, включая её в свой геном в случае высокой степени гомологии с собственной ДНК. Данный процесс носит название трансформация. В природных условиях протекает обмен генетической информацией при помощи умеренных фагов (трансдукция). Кроме этого, возможен перенос нехромосомных генов при помощи плазмид определённого типа, кодирующих этот процесс, процесс обмена другими плазмидами и передачи транспозон.
При горизонтальном переносе новых генов не образуется (как то имеет место при мутациях), однако осуществляется создание разных генных сочетаний. Это важно по той причине, что естественный отбор действует на всю совокупность признаков организма.
Клеточная дифференциация — изменение набора белков (обычно также проявляющееся в изменении морфологии) при неизменном генотипе.
Образование покоящихся форм.
Образование особо устойчивых форм с замедленным метаболизмом, служащих для сохранения в неблагоприятных условиях и распространения (реже для размножения) является наиболее распространённым видом дифференциации у бактерий. Наиболее устойчивыми из них являются эндоспоры, формируемые представителями Bacillus, Clostridium, Sporohalobacter, Anaerobacter (образует 7 эндоспор из одной клетки и может размножаться с их помощью) и Heliobacterium. Образование этих структур начинается как обычное деление и на первых стадиях может быть превращено в него некоторыми антибиотиками. Эндоспоры многих бактерий способны выдерживать 10-минутное кипячение при 100 °C, высушивание в течение 1000 лет и, по некоторым данным, сохраняются в почвах и горных породах в жизнеспособном состоянии миллионы лет.
Менее устойчивыми являются экзоспоры, цисты (Azotobacter, скользящие бактерии и др.), акинеты (цианобактерии) и миксоспоры (миксобактерии).
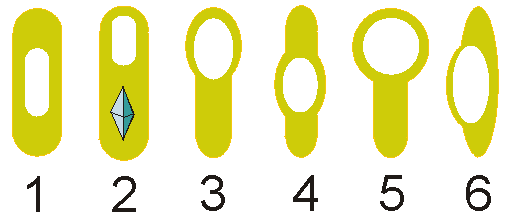
Расположение эндоспор: 1, 4 — центральное, 2, 3, 5 — терминальное, 6 — латеральное.
Другие типы морфологически дифференцированных клеток.
Актиномицеты и цианобактерии образуют дифференцированные клетки, служащие для размножения (споры, а также гормогонии и баеоциты соответственно). Необходимо также отметить структуры, подобные бактероидам клубеньковых бактерий и гетероцистам цианобактерий, служащие для защиты нитрогеназы от воздействия молекулярного кислорода.
Классификация.
Наибольшую известность получила фенотипическая классификация бактерий, основанная на строении их клеточной стенки, включённая, в частности, в IX издание Определителя бактерий Берги (1984—1987). Крупнейшими таксономическими группами в ней стали 4 отдела: Gracilicutes (грамотрицательные), Firmicutes (грамположительные), Tenericutes (микоплазмы; отдел с единственным классом Mollicutes) и Mendosicutes (археи).
В последнее время всё большее развитие получает филогенетическая классификация бактерий, основанная на данных молекулярной биологии. Одним из первых методов оценки родства по сходству генома был предложенный ещё в 1960-х годах метод сравнения содержания гуанина и цитозина в ДНК. Хотя одинаковые значения их содержания и не могут дать никакой информации об эволюционной близости организмов, их различия на 10 % означают, что бактерии не принадлежат к одному роду. Другим методом, произведшим в 1970-е настоящую революцию в микробиологии, стал анализ последовательности генов в 16s рРНК, который позволил выделить несколько филогенетических ветвей эубактерий и оценить связи между ними. Для классификации на уровне вида применяется метод ДНК-ДНК гибридизации. Анализ выборки хорошо изученных видов позволяет считать, что 70 % уровень гибридизации характеризует один вид, 10—60 % — один род, менее 10 % — разные рода.
Филогенетическая классификация отчасти повторяет фенотипическую, так, группа Gracilicutes присутствует и в той и в другой. В то же время систематика грамотрицательных бактерий была полностью пересмотрена, архебактерии и вовсе выделены в самостоятельный таксон высшего ранга, часть таксономических групп разбита на части и перегруппирована, в одни группы объединены организмы с совершенно разными экологическими функциями, что вызывает ряд неудобств и недовольство части научного сообщества. Объектом нареканий становится и то, что проводится фактически классификация молекул, а не организмов.
Происхождение, эволюция, место в развитии жизни на Земле.
Бактерии наряду с археями были одними из первых живых организмов на Земле, появившись около 3,9—3,5 млрд лет назад. Эволюционные взаимоотношения между этими группами ещё до конца не изучены, есть как минимум три основные гипотезы: Н. Пэйс предполагает наличие у них общего предка протобактерии, Заварзин считает архей тупиковой ветвью эволюции эубактерий, освоившей экстремальные местообитания; наконец, по третьей гипотезе археи — первые живые организмы, от которых произошли бактерии.
Эукариоты возникли в результате симбиогенеза из бактериальных клеток намного позже: около 1,9—1,3 млрд лет назад. Для эволюции бактерий характерен ярко выраженный физиолого-биохимический уклон: при относительной бедности жизненных форм и примитивном строении, они освоили практически все известные сейчас биохимические процессы. Прокариотная биосфера имела уже все существующие сейчас пути трансформации вещества. Эукариоты, внедрившись в неё, изменили лишь количественные аспекты их функционирования, но не качественные, на многих этапах циклов элементов бактерии по-прежнему сохраняют монопольное положение.
Одними из древнейших бактерий являются цианобактерии. В породах, образованных 3,5 млрд лет назад, обнаружены продукты их жизнедеятельности — строматолиты, бесспорные свидетельства существования цианобактерий относятся ко времени 2,2—2,0 млрд лет назад. Благодаря им в атмосфере начал накапливаться кислород, который 2 млрд лет назад достиг концентраций, достаточных для начала аэробного дыхания. К этому времени относятся образования, свойственные облигатно аэробной Metallogenium.

Докембрийский строматолит
Появление кислорода в атмосфере (кислородная катастрофа) нанесло серьёзный удар по анаэробным бактериям. Они либо вымирают, либо уходят в локально сохранившиеся бескислородные зоны. Общее видовое разнообразие бактерий в это время сокращается.
Предполагается, что из-за отсутствия полового процесса, эволюция бактерий идёт по совершенно иному механизму, нежели у эукариот. Постоянный горизонтальный перенос генов приводит к неоднозначностям в картине эволюционных связей, эволюция протекает крайне медленно (а, возможно, с появлением эукариот и вовсе прекратилась), зато в изменяющихся условиях происходит быстрое перераспределение генов между клетками при неизменном общем генетическом пуле.
Экология.
Многие бактерии вызывают болезни человека, животных и растений, другие играют исключительно важную роль в функционировании биосферы, например, лишь бактерии способны ассимилировать азот атмосферы. Бактерии являются одними из наиболее просто устроенных живых организмов (кроме вирусов). Полагают, что они — первые организмы, появившиеся на Земле.
Экологические и биосферные функции.
Количество клеток прокариот оценивается в (4—6)*1030, их суммарная биомасса составляет 350—550 млрд т., в ней запасено 60—100 % от углерода всех растений, а запас азота и фосфора в виду их большего относительного содержания в бактериях существенно превосходит запас этих элементов в фитомассе Земли. В то же время бактерии характеризуются коротким жизненным циклом и высокой скоростью обновления биомассы. Уже на основании этого можно оценить их вклад в функционирование основных биогеохимических циклов.
Бактерии способны расти как в присутствии свободного кислорода (аэробы), так и при его отсутствии (анаэробы). Участвуют в формировании структуры и плодородия почв, в образовании полезных ископаемых и разрушении растительной и животной мортмассы; поддерживают запасы углекислого газа и кислорода в атмосфере.
Патогенные бактерии.
Патогенными называются бактерии, паразитирующие на других организмах. Бактерии вызывают большое количество заболеваний человека, таких как чума (Yersinia pestis), сибирская язва (Bacillus anthracis), лепра (проказа, возбудитель: Mycobacterium leprae), дифтерия (Corynebacterium diphtheriae), сифилис (Treponema pallidum), холера (Vibrio cholerae), туберкулёз (Mycobacterium tuberculosis), листериоз (Listeria monocytogenes) и др. Открытие патогенных свойств у бактерий продолжается: в 1976 обнаружена болезнь легионеров, вызываемая Legionella pneumophila, в 1980-е—1990-е было показано, что Helicobacter pylori вызывает язвенную болезнь и даже рак желудка, а также хронический гастрит. Бактериальным инфекциям подвержены также растения и животные. Многие бактерии, являющиеся в норме безопасными для человека или даже обычными обитателями его кожи или кишечника, в случае нарушения иммунитета или общего ослабления организма могут выступать в качестве патогенов. Многие патогенные бактерии образуют скопление в организме в виде биоплёнок, скреплённых и защищённых слизью, что делает их недоступными для проникновения антибиотиков.
Опасность бактериальных заболеваний была сильно снижена в конце XIX века с изобретением метода вакцинации, а в середине XX века с открытием антибиотиков.
Бактерии в мутуалистических отношениях с другими организмами.
Многие бактерии находятся в симбиотических, в том числе в мутуалистических отношениях с другими организмами. Растения, например, выделяют значительную долю созданной в процессе фотосинтеза органики поверхностью корней. Преобразованная таким образом часть почвы (ризосфера) благоприятна для развития бактерий, в том числе азотфиксирующих. Увеличение интенсивности азотфиксации (называемой в таком случае ассоциативной) улучшает условия минерального питания растений. Бактерии-азотфиксаторы обитают также в клубеньках бобовых и других групп растений. В симбиозе со многими морскими животными (прежде всего, губками и асцидиями, а также с некоторыми растениями (например, водным папортником азолоой) и грибами (в составе лишайников) живут и цианобактерии. Хемоавтотрофные бактерии живут в симбиозе с рифтиями и многими другими видами беспозвоночных и протистов, населяющих сообщества гидротерм и сообщества тиобиоса. Есть и много других примеров симбиоза бактерий с самыми разными группами организмов.
Бактерии населяют желудочно-кишечный тракт животных и человека и необходимы для нормального пищеварения. Особенно они важны для травоядных, которые питаются не столько растительной пищей, сколько продуктами её бактериального преобразования, а частично переваривают и самих бактерий.
Бактерии и человек.
Тысячелетиями человек использовал молочнокислых бактерий для производства сыра, йогурта, кефира, уксуса, а также квашения.
В настоящее время разработаны методики по использованию фитопатогенных бактерий в качестве безопасных гербицидов, энтомопатогенных — вместо инсектицидов. Наиболее широкое применение получила Bacillus thuringiensis, выделяющая токсины (Cry-токсины), действующие на насекомых. Помимо бактериальных инсектицидов, в сельском хозяйстве нашли применение бактериальные удобрения.
Бактерии, вызывающие болезни человека, используются как биологическое (бактериологическое) оружие; кроме того, в качестве такого оружия могут использоваться бактериальные токсины.
Благодаря быстрому росту и размножению, а также простоте строения, бактерии активно применяются в научных исследованиях по молекулярной биологии, генетике, генной инженерии и биохимии. Самой хорошо изученной бактерией стала Escherichia coli. Информация о процессах метаболизма бактерий позволила производить бактериальный синтез витаминов, гормонов, ферментов, антибиотиков и др.
Перспективным направлением является обогащение руд с помощью сероокисляющих бактерий, очистка бактериями загрязнённых нефтепродуктами или ксенобиотиками почв и водоёмов.
В кишечнике человека в норме обитает от 300 до 1000 видов бактерий общей массой до 1 кг, а численность их клеток на порядок превосходит численность клеток человеческого организма. Они играют важную роль в переваривании углеводов, синтезируют витамины, вытесняют патогенные бактерии. Можно образно сказать, что микрофлора человека является дополнительным «органом», который отвечает за пищеварение и защиту организма от инфекций.
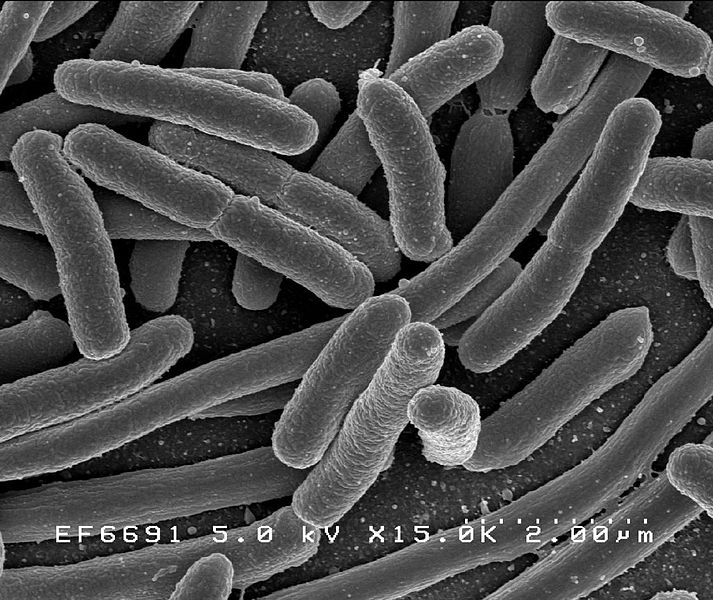
Кишечная палочка (Escherichia coli)
Бактерии в повседневной жизни.
По данным южнокорейского Бюро защиты прав потребителей, количество бактерий на ручках (без антибактериального покрытия) тележек крупных магазинов достигает 1100 колоний на 10 см2. Второе место занимают компьютерные «мышки» в интернет-кафе (690 колоний на ту же площадь). Ручки кабинок общественных уборных содержат лишь 340 колоний вредных микроорганизмов на 10 см2.
Для того, чтобы уберечься от всех видов микроорганизмов, которые были обнаружены на предметах общественного пользования в ходе исследования, достаточно регулярно мыть руки.
Систематика бактерий.
Для идентификации и систематизации бактерий применены критерии, отражающие особенности их физиологии, морфологии, антигенных и других свойств.
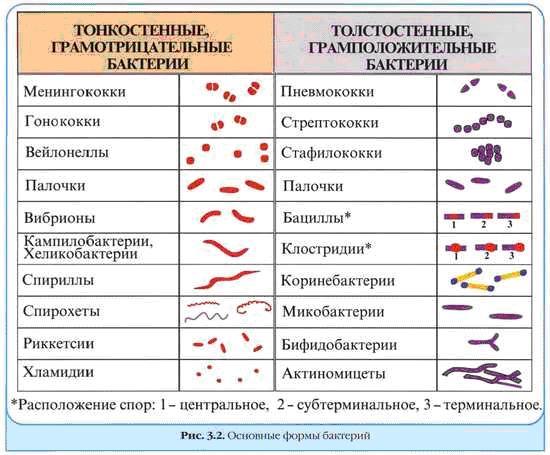
Морфологические и тинкториальные свойства бактерий.
Наиболее общие критерии для важных с медицинской точки зрения бактерий: величина, форма, агрегация (образование нитей, тетрад, пакетов), наличие капсулы, эндоспор, жгутиков, пигментов и способность окрашиваться красителями (то есть тинкториальные свойства). Наиболее распространена окраска по Граму — простое по технике выполнения окрашивание, основанное на способности воспринимать и удерживать внутри клетки красящий комплекс генцианового фиолетового с йодом либо терять его после обработки этанолом.
Грамположительные бактерии хорошо удерживают комплекс генцианового фиолетового с йодом и устойчивы к обесцвечиванию спиртом. После обработки фуксином они окрашиваются в фиолетово-пурпурный цвет.
Грамотрицательные бактерии обесцвечиваются спиртом, то есть теряют комплекс генцианового фиолетового с йодом, и хорошо поглощают фуксин. В мазках они окрашиваются в малиново-красный цвет.

Кислотоустойчивые бактерии. Клеточная стенка некоторых бактерий содержит большое количество липидов и восков, делающих их устойчивыми к последующему после окрашивания обесцвечиванию кислотами, щелочами или этанолом (например, виды Mycobacterium или Nocardia). Подобные бактерии называют кислотоустойчивыми, их трудно окрашивать по Граму (хотя кислотоустойчивые бактерии рассматривают как грамположительные). Для их окраски применяют метод Циля-Нильсена.
Окрашивание по Граму или Цилю-Нильсену имеет диагностическую ценность в отношении бактерий, обладающих прочной клеточной стенкой. Они неприемлемы для окраски микоплазм (нет клеточной стенки) или спирохет (клеточная стенка тонкая и легко разрушается при окрашивании). Для изучения последних применяют различные методы нанесения на их поверхность контрастных субстратов (например, серебрение).
Подвижность бактерий.
Важным дифференцирующим признаком является подвижность. В соответствии со способом передвижения выделяют скользящие бактерии, перемещающиеся за счёт волнообразных сокращений тела, и плавающие бактерии, движение которых обеспечивают жгутики или реснички.
Способность к спорообразованию бактерий.
Для классификации некоторых бактерий учитывают их способность к спорообразованию, размеры спор и их расположение в клетке.
Физиологическая активность бактерий.
Физиологическая активность является не менее важным отличительным признаком. Бактерии разделяют по способу питания, по типу получения энергии (дыхание, брожение, фотосинтез), по отношению к рН с указанием пределов устойчивости и оптимума роста и т.д. Наиболее важным критерием считают отношение к кислороду.
Аэробные бактерии используют молекулярный O2 в качестве конечного акцептора электронов в процессе дыхания. Большинство бактерий обладают связанной с мембраной цитохром-С-оксидазой, играющей ведущую роль в электронотранспортной цепи. Для выявления фермента применяют оксидазный тест, основанный на способности бесцветного вещества N, N-диметил-р-фенилендиамина приобретать при восстановлении малиновый цвет.
Анаэробные бактерии не утилизируют молекулярный O2 в качестве конечного акцептора электронов. Подобные бактерии получают энергию либо в процессе брожения, где конечными акцепторами электронов служат органические соединения, либо при анаэробном дыхании, используя отличный от кислорода акцептор электронов (например, NO -, SO4 - или Fe3+). Факультативные бактерии могут получать энергию либо в процессе дыхания, либо при брожении в зависимости от наличия или отсутствия кислорода в окружающей среде.
Биохимические свойства бактерий.
Для дифференцировки бактерий изучают их способность ферментировать углеводы, образовывать различные продукты (сероводород, индол) или гидролизовать белки.
Антигенные свойства бактерий.
Антигенные свойства бактерий различных бактерий специфичны и связаны с особенностями строения клеточных структур, распознаваемых специальными антисыворотками как антигенные детерминанты. Типирование бактерий по антигенной структуре проводят в реакции агглютинации (РА), смешивая каплю антисыворотки с каплей бактериальной суспензии. При положительной реакции появляются отдельные агрегированные комочки в исходно однородной бактериальной суспензии. Выделяют следующие типы:
• родоспецифичные, выявляемые у всех представителей конкретного рода, включая отдельные штаммы;
• видоспецифичные, выявляемые у отдельных видов и штаммов микроорганизмов;
• сероваро- (штаммо-) специфичные, выявляемые у представителей различных подгрупп (штаммов) внутри конкретного вида.
Чувствительность бактерий к бактериофагам.
Поскольку вирусы бактерий взаимодействуют только с чувствительными бактериями, то в ряде случаев для дифференцировки бактерий применяют их чувствительность к бактериофагам.
Химический состав бактерий.
Важный классификационный признак — суммарный химический состав бактериальных клеток. Обычно определяют содержание и состав Сахаров, липидов и аминокислот в клеточных стенках.
Генетическое родство бактерий.
Для филогенетической классификации бактерий лучший и наиболее информативный показатель — генетическое родство. При систематизации бактерий на основании генетического родства учитывают ряд показателей.
• Способность обмениваться генетической информацией (например, в процессе трансформации или конъюгации), возможной только между организмами одного рода или вида.
• Состав оснований ДНК (отношение гуанин-цитозин : аденин-тимин).
• Сходство нуклеиновых кислот, выявляемое методом гибридизации.
• "Золотой стандарт" в определении степени родства среди бактерий — установление сходства в последовательности нуклеиновых кислот. Однако в результате обмена генетической информацией генотип бактерий может существенно изменяться, поэтому подобный подход к систематике оказывается на практике неудобным.

Определитель Берджи.
Определитель Берджи систематизирует все известные бактерии по нашедшим в практической бактериологии наибольшее распространение принципам идентификации бактерий, основанным на различиях в строении клеточной стенки и отношении к окраске по Граму. Определитель выделяет четыре основных категории бактерий — Gracillicutes [от лат. gracilis, изящный, тонкий, + cutis, кожа] — виды с тонкой клеточной стенкой, окрашивающиеся грамотрицательно; firmicutes [от лат. flrmus, крепкий, + cutis, кожа] — бактерии с толстой клеточной стенкой, окрашивающиеся грамположительно; Tenericutes [от лат. tener, нежный, + cutis, кожа] — бактерии, лишённые клеточной стенки (микоплазмы и прочие представители класса Mollicutes) и Mendosicutes [от лат. mendosus, неправильный, + cutis, кожа] — архебактерии (метан- и сульфатредуцирующие, галофильные, термофильные и архебактерии, лишённые клеточной стенки). Описание бактерий даётся по группам (секциям), в состав которых включены семейства, роды и виды; в некоторых случаях в состав групп входят классы и порядки. Патогенные для человека бактерии входят в небольшое число групп.
Группа 1 определителя Берджи. Спирохеты. Включает свободноживущие и паразитические виды; для человека патогенны представители родов Treponema, Borrelia и Leptospira.
Группа 2 определителя Берджи. Аэробные и микроаэрофильные подвижные извитые и изогнутые грамотрицательные бактерии. Патогенные для человека виды входят в роды Campylobacter, Helicobacters, Spirillum.
Группа 3 определителя Берджи. Неподвижные (редко подвижные) грамотрицательные бактерии. Не содержит патогенные виды.
Группа 4 определителя Берджи. Грамотрицательные аэробные и микроаэрофильные палочки и кокки. Патогенные для человека виды включены в состав семейств Legionellaceae, Neisseriaceae и Pseudomonadaсеае, в группу входят также патогенные и условно-патогенные бактерии родов Acinetobacter, Afipia, Alcaligenes, Bordetella, Brucella, Flavobacterium, Francisella, Kingella и Moraxella.
Группа 5 определителя Берджи. Факультативно анаэробные грамотрицательные палочки. Группа образована тремя семействами — Enterobacteriaceae, Vibrionaceae и Pasteurellaceae, каждое из которых включает патогенные виды, а также патогенные и условно-патогенные бактерии родов Calymmobaterium, Cardiobacterium, Eikenetta, Gardnerella и Streptobacillus.
Группа 6 определителя Берджи. Грамотрицательные анаэробные прямые, изогнутые и спиральные бактерии. Патогенные и условно-патогенные виды входят в состав родов Bacteroides, Fusobacterium, Porphoromonas и Prevotelta.
Группа 7 определителя Берджи. Бактерии, осуществляющие диссимиляционное восстановление сульфата или серы Не включает патогенные виды.
Группа 8 определителя Берджи. Анаэробные грамотрицательные кокки. Включает условно-патогенные бактерии poда Veillonella.
Группа 9 определителя Берджи. Риккетсии и хламидии. Три семейства — Rickettsiaceae, Bartonellaceae и Chlamydiaсеае, каждое из которых содержит патогенные для человека виды.
Группы 10 и 11 определителя Берджи включают анокси- и оксигенные фототрофные бактерии, не патогенные для человека.
Группа 12 определителя Берджи. Аэробные хемолитотрофные бактерии и родственные организмы. Объединяет серо- железо- и марганецокисляющие и нитрифицирующие бактерии, не вызывающие поражения у человека.
Группы 13 и 14 определителя Берджи включают почкующиеся и/или обладающие выростами бактерии и бактерии образующие футляры. Представлены свободноживущими видами, не патогенными для человек;
Группы 15 и 16 определителя Берджи объединяют скользящие бактерии, не образующие плодовые тела и образующие их. Группы не включают виды, патогенные для человека.
Группа 17 определителя Берджи. Грамположительные кокки. Включает условно-патогенные виды родов Enterococcus, Leuconostoc, Peptococcus, Peptostreptococcus, Sarcina, Staphylococcus, Stomatococcus, Streptococcus.
Группа 18 определителя Берджи. Спорообразующие грамположительные палочки и кокки. Включает патогенные, условно-патогенные палочки родов Clostridium и Bacillus.
Группа 19 определителя Берджи. Споронеобразующие грамположительные палочки правильной формы. Включая условно-патогенные виды родов Erysipelothrix и Listeria.
Группа 20 определителя Берджи. Споронеобразующие грамположительные палочки неправильной формы. В состав группы входят патогенные и условно-патогенные виды родов Actinomyces, Corynebacterium, Gardnerella, Mobiluncus и др.
Группа 21 определителя Берджи. Микобактерии. Включает единственный род Mycobacterium, объединяющий патогенные и условно-патогенные виды.
Группы 22-29. Актиномицеты. Среди многочисленных видов лишь нокардиоформные актиномицеты (группа 22) родов Gordona, Nocardia, Rhodococcus, Tsukamurella, Jonesia, Oerskovi и Terrabacter способны вызывать поражения у человека.
Группа 30 определителя Берджи. Микоплазмы. Патогенны для человека виды, включённые в состав рода Acholeplasma, Mycoplasma и Ureaplasma.
Остальные группы определителя Берджи — метаногенные бактерии (31), сульфатредуцируюшие бактерии (32 экстремально галофильные аэробные архебактерии (33), архебактерии, лишённые клеточно стенки (34), экстремальные термофилы и гипертермофилы, метаболизируюшие серу (35) — не содержат патогенные для человека виды.