(Monopisthocotylea) Отряд Монопистокотилы, Order Monopisthocotylea 13 семейств
Жизнь животных. Том 1. Беспозвоночные Под редакцией профессора Л.А.Зенкевича 1968 г.
КЛАСС МОНОГЕНОИДЕИ (MONOGENOIDEA)
В системе типа плоских червей моногеноидеи (моногенетические сосальщики) только в недавнее время стали рассматриваться как класс. Ранее они составляли подкласс в классе трематод наряду с другим подклассом — дигенеями.
Указанные изменения внесены в систему плоских червей в результате детального изучения биологии моногеноидей, проведенного в последнее время советским ученым Б. Е. Быховским. Он установил, что по характеру своего индивидуального развития моногеноидеи стоят ближе к другим группам плоских червей — к гирокотилидам и цестодам, чем к дигенетическим трематодам. Общим признаком, сближающим гирокотилид, цестод и моногеноидей, оказалось строение личинок этих групп животных, а именно наличие на ранних стадиях развития у всех их своеобразного хвостового отростка, снабженного зародышевыми крючьями. Личинки дигенетических трематод подобного отростка с зародышевыми крючьями не имеют.
От дигенетических трематод моногеноидеи отличаются также и рядом других существенных признаков. По-иному, прежде всего, протекает жизненный цикл моногенетических сосальщиков. В отличие от дигеней, их развитие не связано со сменой хозяев и со сменой поколений. Значительные отличия имеются в строении взрослых форм представителей этих двух групп сосальщиков. Выделительная система, например, у моногеноидей открывается наружу отверстиями, расположенными в передней части тела паразита, у дигеней же выделительное отверстие находится в задней части тела. Это важное отличие. Отмеченные особенности и послужили основанием для выделения моногеноидей из класса трематод.
С другой стороны, хотя и было установлено их родство с гирокотилидами и цестодами, они не могли быть объединены ни с одним из указанных классов. Этому препятствовало своеобразие их строения и биологии. В результате был основан новый класс.
Название класса — моногеноидеи — в переводе с латинского языка означает «имеющие одно поколение». Названием подчеркивалось отличие их от дигенетических трематод, которым свойственна смена поколений.
По-русски данный класс называют Сосальщики-многоустки.
Моногеноидеи — сравнительно небольшой класс плоских червей, он включает около 1000 видов, паразитирующих у морских и пресноводных рыб и очень редко у амфибий, рептилий и водных млекопитающих.
В отличие от других классов паразитических плоских червей — трематод и цестод, поселяющихся во внутренних органах своих хозяев, моногеноидеи — наружные паразиты (эктопаразиты). Основное место их обитания — жабры и кожа рыб.
Моногеноидеи приносят большой экономический ущерб рыбному хозяйству, так как многие представители их являются возбудителями очень серьезных заболеваний рыб. Особенно ощутимы потери от этих паразитов в прудовых хозяйствах.
Строение. Почти все моногеноидеи имеют уплощенное, вытянутое в длину тело, расширенное в задней части.
Размеры моногеноидей в большинстве случаев незначительны — длина тела колеблется в пределах 0,03—20 мм, в редких случаях несколько больше. Паразиты морских рыб обычно крупнее, чем паразиты пресноводных.
Передний конец червя с двумя — четырьмя лопастями. Иногда лопасти несут ямки или присоски.
Задний конец червя более или менее обособлен от тела и превращен в прикрепительный диск, который снабжен различного рода хитиноидными образованиями — крючьями и соединительными пластинками. На нем же могут находиться особые клапаны, ямки и выросты, способствующие более прочной фиксации паразита. Иногда весь диск преобразован в мощную присоску. Строение прикрепительных органов моногеноидей отличается большим разнообразием. Различие в строении этих органов широко используется при систематическом определении представителей класса.
Снаружи тело моногеноидей покрыто кутикулой. Под ней находятся субкутикулярный и мышечные слои, составляющие кожно-мускульный мешок. Внутренняя часть тела заполнена паренхимной тканью.
В паренхиме располагаются органы пищеварительной, выделительной, нервной и половой систем.
Пищеварительный канал начинается ротовым отверстием, расположенным на переднем конце тела. Отверстие ведет в околоротовую воронку, переходящую в мускулистую глотку, за ней следует пищевод (он имеется не у всех форм) и далее кишечная трубка, которая заканчивается слепо.
Кишечник состоит из одного или двух стволов. Они могут быть гладкие или с боковыми ответвлениями.
Выделительная система представлена сетью каналов. Главные боковые каналы открываются наружу в передней части тела двумя выделительными порами.
Нервная система состоит из парных головных: ганглиев, от которых вперед, и назад отходят 3—4 пары нервных стволов. У некоторых видов во взрослой и личиночной стадии, чаще только в личиночной, имеются особые органы чувств — «глазные пятна», находящиеся на головном конце паразита.
Половая система построена по гермафродитному типу. Мужская система состоит из одного, двух или многих семенников. Отходящие от них семявыносящие канальцы объединяются затем в общий семяпровод, оканчивающийся копулятивным органом. Яичник всегда один. Отходящий от него короткий яйцевод принимает протоки желточников и влагалища. Яйцевод расширяется в оотип, куда впадает проток скорлуповой железы (тельце Мелиса). От оотипа берет начало сравнительно короткая трубка матки. Наружу матка открывается общим с мужской половой системой гермафродитным отверстием, расположенным в передней части тела. Трубка влагалища открывается самостоятельным отверстием, моногеноидей очень разнообразны по форме, они могут быть шаровидные, овальные, пирамидальные и иной конфигурации. На одном полюсе яйца обычно имеется крышечка с отростками, на другом — ножки с филаментами. Некоторые виды моногеноидей выделяют не яйца, а личинку.
Развитие моногеноидей протекает без смены хозяев и чередования поколений. Однако вышедшая из яйца личинка не походит на взрослую форму и в процессе своего развития претерпевает довольна сложные изменения. Она имеет сигарообразную форму, покрыта полностью или частично ресничками, обладает головными железами, у большинства видов два — четыре глазка, глотка, мешковидный кишечник и слаборазвитая нервная и выделительная системы. Свободно плавает в воде.
Попав к своему хозяину, личинка прикрепляется к телу его в месте своего обитания, сбрасывает ресничный покров и, переходя к паразитическому образу жизни, начинает расти, превращаясь постепенно во взрослую форму.
Моногеноидеи отличаются резко выраженной специфичностью в отношении своих хозяев: обычно каждый вид паразита приурочен к определенному виду хозяина; случаи, когда представители одного вида поселяются в нескольких хозяевах, редки.
Классификация. Класс моногеноидей подразделяется на два подкласса (Polyonchoinea и Oligonchoinea). Каждый подкласс объединяет по три отряда. Подразделение на систематические единицы основывается главным образом на морфологических особенностях представителей класса, преимущественно на строении элементов прикрепительного аппарата.
Ниже мы расскажем о нескольких наиболее обычных видах данного класса.
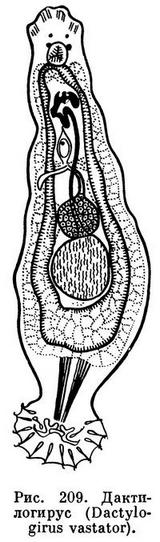
Дактилогирус вастатор (Dactylogyrus vastator) паразитирует на жабрах, редко на коже карпов, отличается большой вредоносностью, в связи с чем представляет очень серьезную опасность для прудовых хозяйств.
Тело паразита имеет длину от 0,5 до 1 мм, ширину 0,15—0,38 мм. На переднем конце его — четыре головные лопасти. Взрослые формы с четырьмя глазками.Прикрепительный диск снабжен двумя центральными большими крючьями и четырнадцатью мелкими краевыми крючочками. Рот не окружен присоской.
Размножается дактилогирус путем откладки яиц. В матке одновременно находится всего лишь одно яйцо, однако плодовитость паразита довольно высокая. Подсчитано, что за шесть часов он откладывает от 14 до 29 яиц.
Из яйца выходит личинка удлиненной формы с пятью группами ресничек и четырьмя глазками. Ее зачаточный прикрепительный диск снабжен четырнадцатью краевыми крючочками. Большие крючья отсутствуют.
Поплавав некоторое время в воде, личинка прикрепляется с помощью своих крючков к рыбе и сбрасывает реснички. Вначале она поселяется на коже карпа, но через некоторое время переползает на его жабры. Здесь личинка формируется во взрослого червя.
Дактилогирус вастатор паразитирует преимущественно у молодняка карпов. Наибольшей восприимчивостью отличаются мальки длиной от 2 до 5 см. У более крупных карпов паразит встречается в меньшем количестве и поражает меньший процент рыб.
Внешними признаками зараженности рыб дактилогирусами являются неравномерное окрашивание жабр и обильное выделение слизи. Зараженная рыба делается беспокойной, устремляется к притоку воды, поднимается к поверхности, заглатывает воздух.
При дальнейшем развитии болезни, если рыба заражена большим количеством паразитов, наблюдается омертвление пораженных участков жабр с последующим разрушением их.
Заболевание, вызываемое дактилогирусами, достигает наибольшей интенсивности в прудовых хозяйствах обычно в июле — августе. Особенно сильно болезнь развивается при густой посадке рыб в пруду, при плохом питании и неблагоприятных для рыбы условиях окружающей среды.
Для лечения больных рыб в прудовых хозяйствах применяют ванны с растворами химических веществ, губительно действующих на паразитов и существенно не отражающихся на состоянии их хозяев. Чаще других применяют растворы поваренной соли и медного купороса.
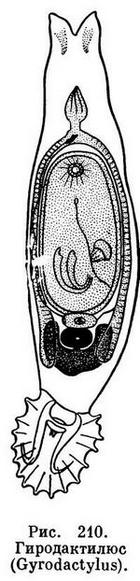
Гиродактилюс элеганс (Gyrodactylus elegans) паразитирует на коже и плавниках карпов и некоторых других близкородственных им рыб. Это один из наиболее опасных паразитов в прудовых хозяйствах.
Длина взрослой особи гиродактилюса часто колеблется от 0,5 до 1 мм при ширине 0,12-0,15 мм. Передний конец с двумя лопастями, способными сильно сокращаться. На вершинах лопастей открываются отверстия головных железок. Глазные пятна отсутствуют. Прикрепительный диск, расположенный на заднем конце, вооружен двумя большими центральными крючьями и шестнадцатью мелкими краевыми. Между большими крючьями имеются две пластинки. Ротовое отверстие помещается на брюшной стороне в передней трети тела. Кишечник подразделяется на два ствола.
Гиродактилюс принадлежит к группе живородящих моногеноидей.
Формирующийся внутри тела паразита зародыш отличается крупными размерами. Зародыш имеет два больших крючка. Особенно интересно, что внутри его помещается другой зародыш несколько меньших размеров и также с двумя крючками, а в нем в свою очередь находится третий зародыш гиродактилюса.
Долгое время не могли разгадать, каким образом осуществляется подобное развитие зародышей. Тайна процесса была раскрыта только в последнее время. Происходит это следующим образом. Оплодотворенное яйцо делится пополам. Из одной половинки образуется зародыш. Он обхватывает собой другую половинку яйца. Эта половинка в свою очередь снова делится пополам. Одна из образовавшихся частей дает начало новому зародышу, который также обхватывает собой оставшуюся часть яйца. Эта четвертая часть яйца опять-таки превращается в нового зародыша. Таким образом возникают три зародыша, как бы вложенных друг в друга.
Выделяемый гиродактилюсом зародыш представляет собой вполне развитую личинку. Долго существовать как свободноживущий организм он не может. Встретившись с хозяином, личинка прикрепляется к поверхности тела и к плавникам рыбы, переходит к паразитическому образу жизни и начинает развиваться во взрослого червя.
У рыб, сильно зараженных гиродактилюсами, тело покрывается голубовато-серым налетом, состоящим из обильно выделяющейся слизи, которой и питаются паразиты. При дальнейшем развитии болезни участки кожи могут подвергаться разрушению и омертвлению. В некоторых случаях на поверхности кожи появляются небольшие изъязвления. Плавники, на которых паразитируют гиродактилюсы, могут разрушаться настолько сильно, что от них остаются только свободно торчащие отдельные лучи.
Заражаются гиродактилюсами главным образом мальки и годовики рыб.
Для лечения больных рыб применяют ванны с раствором веществ, которые убивают паразитов и от которых не гибнут рыбы. Обычно для этого используют поваренную соль. Как меру предохранения от заноса паразита в пруды, где выращивается молодь, применяют обязательное использование солевых ванн для производителей карпов перед посадкой их на нерест.
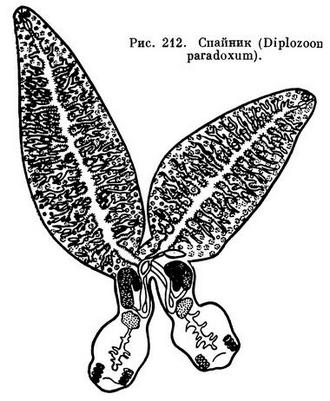
Спайник парадоксальный (Diplozoon paradoxum) — это, как показывает название животного, несколько необычный представитель класса. Своеобразие его заключается в том, что паразиты живут попарно. Соединяются они в молодом возрасте. При этом брюшная присоска одной особи охватывает специальный спинной бугорок другой, и наоборот. В местах соединения животные полностью срастаются, оставаясь в таком состоянии всю жизнь. В месте срастания мужской проток одного червя соединяется с женским протоком другого: этим обеспечивается перекрестное оплодотворение. Если молодой спайник не встретится с другой особью, он погибает.
Спайник парадоксальный паразитирует на жабрах леща. Хозяевами его могут быть и некоторые другие виды карповых рыб. Однако в последнее время высказано мнение, что спайники, паразитирующие не на лещах, принадлежат к другим видам данного рода.
По внешнему виду спайник представляет собой крестообразную фигуру. Тело каждого червя из сросшейся пары довольно четко подразделяется на две части: переднюю — листообразную и заднюю — значительно меньшую, цилиндрическую по форме. В передней части располагаются желточники и основная часть кишечника, в задней находятся органы половой системы и прикрепительный аппарат, состоящий из восьми ущемляющих клапанов и срединных крючочков. Длина особи спайника 3,8—6,7 мм, ширина 0,7—1,7 мм.
Размножается спайник путем откладки яиц, из которых в воде выходят личинки. Прикрепившись к жабрам леща, личинки затем соединяются попарно и вырастают во взрослых паразитов.