(Anthozoa Ehrenberg, 1834) Класс Антозои, Коралловые полипы, Class Anthozoa Ehrenberg, 1834 (Stony corals, Soft corals, Sea anemones) 10 отрядов
Жизнь животных. Том 1. Беспозвоночные Под редакцией профессора Л.А.Зенкевича 1968 г.
Коралловые полипы (Anthozoa).
Тело кораллового полипа обычно имеет цилиндрическую форму и не подразделяется на туловище и ножку. У колониальных форм нижний конец тела полипа прикреплен к колонии, а у одиночных полипов снабжен прикрепительной подошвой. Щупальца у коралловых полипов расположены в один или несколько тесно сближенных венчиков. Различают две большие группы коралловых полипов: восьмилучевые (Octocorallia) и шестилучевые (Нехасоrallia). У первых всегда 8 щупалец, и они снабжены по краям маленькими выростами — пиннулами, у вторых число щупалец обычно бывает довольно большим и, как правило, кратно шести. Щупальца шестилучевых кораллов гладкие, без пиннул.
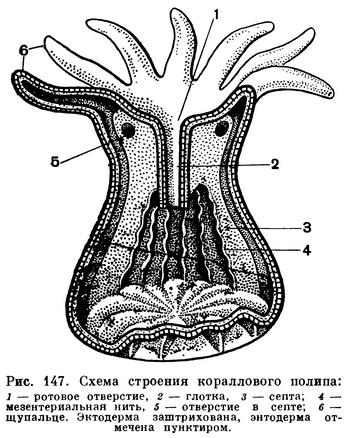
Верхняя часть полипа, между щупальцами, называется ротовым диском. В его середине расположено щелевидное ротовое отверстие. Рот ведет в глотку, выстланную эктодермой. Один из краев ротовой щели и нисходящей от него глотки получили название сифоноглифа. Эктодерма сифоноглифа покрыта эпителиальными клетками с очень крупными ресничками, которые находятся в беспрерывном движении и гонят воду внутрь кишечной полости полипа. Кишечная полость кораллового полипа разделена продольными энтодермальными перегородками (септами) на камеры.
В верхней части тела полипа перегородки прирастают одним краем к стенке тела, а другим к глотке. В нижней части полипа, ниже глотки, септы прикрепляются лишь к стенке тела, вследствие чего центральная часть гастральной полости — желудок — остается неразделенной. Число септ соответствует числу щупалец. По каждой септе, вдоль одной из ее сторон, проходит мускульный валик. Свободные края септ утолщены и получили названия мезентериальных нитей. Две из этих нитей, расположенных на паре соседних септ, противостоящих сифоноглифу, покрыты особыми клетками, несущими длинные реснички. Реснички находятся в постоянном движении и гонят воду из гастральной полости наружу. Совместная работа ресничного эпителия этих двух мезентериальных нитей и сифоноглифа обеспечивает постоянную смену воды в гастральной полости. Благодаря им в кишечную полость все время поступает свежая, богатая кислородом вода. Виды, питающиеся мельчайшими планктонными организмами, получают при этом также и пищу. Остальные мезентериальные нити играют важную роль в пищеварении, так как образованы железистыми энтодермальными клетками, выделяющими пищеварительные соки.
Половые железы развиваются в энтодерме септ. У коралловых полипов очень большую роль играет скелет.
Восьмилучевые кораллы имеют скелет, состоящий из отдельных известковых иголочек — спикул, находящихся в мезоглее. Иногда спикулы соединяются между собой, сливаясь или же объединяясь органическим рогоподобным веществом.
Среди шестилучевых кораллов имеются бесскелетные формы, такие, как актинии. Чаще, однако, скелет у них имеется, причем он может быть либо внутренним — в виде стержня из рогоподобного вещества, либо наружным — известковым.
Особенно большой сложности достигает скелет у представителей группы мадрепоровых. Он выделяется эктодермой полипов и на первых порах имеет вид пластинки или низкой чашечки, в которой сидит сам полип. Далее скелет начинает разрастаться, на нем возникают радиальные ребра, соответствующие септам полипа. Вскоре полип оказывается как бы насаженным на скелетную основу, которая глубоко вдается снизу в его тело, хотя и отграничена всюду эктодермой. Скелет мадрепоровых кораллов получает очень сильное развитие: мягкие ткани покрывают его в виде тонкой пленки.
Скелет кишечнополостных играет роль опорной системы, а вместе со стрекательным аппаратом представляет мощную защиту от врагов, что способствовало их существованию в течение длительных геологических периодов.
Для коралловых полипов характерно только полипоидное состояние, медуз они не образуют.
ЖИЗНЕННЫЙ ЦИКЛ КИШЕЧНОПОЛОСТНЫХ, ЧЕРЕДОВАНИЕ ПОКОЛЕНИЙ
Из современных нам кишечнополостных животных наиболее просто завершается жизненный цикл коралловых полипов. Оплодотворенное яйцо начинает дробиться. Сначала оно делится надвое, затем каждая из образовавшихся клеточек в свою очередь также разделяется, и так далее. В результате образуется большое количество мелких клеток, расположенных в один слой и имеющих вид маленького полого шарика. Вслед за этим часть клеток начинает погружаться внутрь, в результате чего получается двуслойный зародыш. Из его внутреннего слоя впоследствии формируется энтодерма, а из наружного — эктодерма будущего полипа. Эктодерма покрыта многочисленными мелкими ресничками, при помощи которых зародыш получает способность плавать; с этого момента он превращается в личинку, называемую планулой. Планула не способна питаться и размножаться. Она некоторое время плавает в толще воды, затем садится на дно, прикрепляясь к нему передним концом. Вскоре после этого на заднем (теперь уже верхнем) конце планулы прорывается ротовое отверстие и образуется венчик щупалец. Так возникает первый полип. У колониальных форм этот полип вскоре выпочковывает на себе других полипов, те в свою очередь следующих и т. д. Возникает колония. По достижении колонией определенной степени развития составляющие ее полипы начинают размножаться также и половым путем, образуя яйца. На этом цикл замыкается.
КЛАСС КОРАЛЛОВЫЕ ПОЛИПЫ (ANTHOZOA)
ПОДКЛАСС ВОСЬМИЛУЧЕВЫЕ КОРАЛЛЫ (OCTOCORALL1A)
ОТРЯД ГОРГОНОВЫЕ КОРАЛЛЫ (GORGONARIA)
Горгонарии, или роговые кораллы, внешне очень похожи на колонии гидроидных полипов. Их скелет состоит из образующихся в толще мезоглеального слоя известковых спикул в виде покрытых бугорками палочек и чешуек.
Но для горгонарий более характерен плотный осевой скелет, пронизывающий ствол и ветви колоний. В его образовании в простейшем случае участвуют те же известковые спикулы, соединенные между собой промежуточным веществом. У большинства же горгонарий осевой скелет слагается из концентрических роговых пластинок, образующихся в мезоглеальном слое. Эти роговые пластинки обычно пропитаны известью. У многих горгонарий спикулы, из которых состоит скелет отдельных полипов, скапливаются вокруг ротового диска, образуя венчик торчащих наружу защитных игл и чешуек.
Колонии горгонарий прочно прикрепляются к твердому субстрату при помощи округлого расширения основания. Лишь немногие виды способны развиваться на мягких песчаных или илистых грунтах. Тогда в базальной части колонии образуются корневые выросты. На мелководье, основном месте обитания горгонарий, они особенно многочисленны и разнообразны лишь там, где находят необходимый для своего укрепления скалистый грунт. На соседних участках, где дно песчаное или илистое, можно встретить единичные, к тому же обычно чахлые и недоразвитые колонии горгонарий. Однако у глубоководных представителей корневидные выросты основания колоний — более обычное образование. У некоторых глубоководных видов, приспособившихся к обитанию на илистых грунтах, нижняя часть ствола колонии разрастается в ширину и принимает характер лопатовидной пластинки, свободно втыкающейся в грунт.
Горгонарии распространены всесветно от полярных районов до экватора. Но особенно многочисленны и разнообразны они в водах тропической зоны. К обитанию в холодных водах полярных районов приспособились очень немногие виды. Так, в водах Арктической области встречено только 5 видов горгонарий, а в Антарктике их не более 30. Общее же число видов роговых кораллов достигает 1200. На мелководье обитает около 70% всех известных видов горгонарий. Только 16% их опускается на глубину более 1000 м. В абиссаль Мирового океана проникло всего лишь 11 видов горгонарий. Подавляющее большинство роговых кораллов может жить только в воде с нормальной морской соленостью, и лишь несколько видов способно переносить незначительное опреснение. Поэтому в таких морях, как Черное или Балтийское, горгонарии не встречаются.
Способ питания и состав пищи горгонарий остаются до сих пор неизвестными. В гастральных полостях почти никогда не удается найти остатки проглоченной пищи. Возможно, что пищей горгонариям служат мельчайшие частицы органического вещества животного происхождения, взвешенные в морской воде. В таком случае остатки мелких ракообразных, которые бывали изредка обнаружены в гастральных полостях, очевидно, явились случайным приловом для полипов-фильтраторов.
ОТРЯД МЯГКИЕ КОРАЛЛЫ (ALCYONARIA)
Слово «коралл» в нашем представлении всегда связано с чем-то твердым, хрупким. Таковы в действительности многие кораллы, но имеются и мягкие кораллы — альционарии.
К альционариям относятся наиболее просто организованные колониальные коралловые полипы, в колониях которых никогда не образуется центральный, или осевой, скелет. Многочисленные склериты или беспорядочно разбросаны в толще мезоглеального слоя, или образуют корковый слой непосредственно под эктодермальным слоем клеток. В более редких случаях склериты, сливаясь между собой, образуют вокруг полипов изящные трубочки. Дольчатые, стелющиеся, грибовидные и шаровидные колонии альционарий прочно прикрепляются к субстрату мясистым основанием. Но некоторые альционарии способны укрепляться и на мягких илистых грунтах. Различно устроенные выросты основания служат тогда колониям хорошими якорями.
Альционарии — наиболее многочисленный отряд восьмилучевых кораллов. Они широко распространены в морях и океанах, встречаясь от полярных районов до экватора и от литорали до больших глубин. Однако в Мировом океане альционарии распределены весьма неравномерно. Более 80% всех известных в настоящее время видов обитает на мелководье тропических морей Малайского архипелага и вдоль северных берегов Австралии. В холодные воды полярных районов северного и южного полушарий проникает всего несколько видов альционарий. Не более 2% видов встречается на глубине свыше 1000 м, а в абиссаль Мирового океана (на глубину более 3 тыс. м) проникло всего лишь шесть видов альционарий.
В отряде Alcyonaria различают до 40 родов и 1200 видов мягких кораллов, неравномерно распределяющихся между тремя подотрядами — Stolonifera, Telestacea и Alcyonacea.
ОТРЯД МОРСКИЕ ПЕРЬЯ (PENNATULARIA)
Из всех кишечнополостных самые фантастические по форме колонии образуют морские перья, относящиеся к восьмилучевым кораллам.
В отличие от других колониальных кишечнополостных морские перья никогда не прикрепляются к какому-нибудь субстрату. Их нижний конец, имеющий обычно мускулистое расширение, свободно втыкается в мягкий грунт, выстилающий дно. Хотя морские перья ведут неподвижный образ жизни, они могут в случае необходимости передвигаться по грунту и вновь укрепляться на новом месте.
Морские перья образуют неразветвленные, часто довольно крупные и ярко окрашенные колонии. Они состоят из мясистого ствола, представляющего собой разросшееся и сильно видоизмененное тело первичного полипа. Остальные полипы вырастают на стенках ствола или на его выростах. Для всех морских перьев характерен диморфизм вторичных полипов. Имеются крупные кормящие полипы — аутозоиды — и мелкие недоразвитые полипы — сифонозоиды. Каждая колония морского пера делится на два отдела: верхний, или полипоидный, и нижний — стебель. На полипоидной части колонии образуются аутозоиды и сифонозоиды. Стебель свободен от полипов, хотя у некоторых видов и здесь развиваются мелкие сифонозоиды.
Для морских перьев характерна высокоорганизованная система каналов, связывающая различные отделы колонии. Она состоит из центральной и периферической систем. Центральная система представлена четырьмя основными продольными каналами, пронизывающими всю колонию. Два из них являются продолжением радиальных карманов гастральной полости первичного полипа. Два других канала — вторичные образования. Периферическая система состоит из сложной густой сети каналов, связывающих между собой гастральные полости отдельных полипов. Каналы периферической системы открываются в основные продольные каналы колонии.
Скелет морских перьев слагается из многочисленных известковых спикул в виде палочек и игл, обычно имеющих три продольных ребрышка. Кроме спикул, образующих, как правило, корковый слой тела колонии, имеется и осевой скелет, образующийся, так же как и у роговых кораллов, из концентрических роговых пластинок, пропитанных известью.
Морским перьям свойственна хорошо развитая мышечная система. В ней различают мощную кольцевую мускулатуру, отражающую продольные каналы, слой продольных мышечных тяжей и слой косых мускулов, прикрепляющихся одним концом к стержню осевого скелета. Косые мускулы обеспечивают произвольное изгибание колонии.
Нервная система морских перьев достигает высокого для кишечнополостных совершенства. Основная черта ее прогресса заключается в образовании общеколониальной нервной системы, связывающей между собой все отделы колонии. Однако специальные органы чувств отсутствуют и у морских перьев.
ПОДКЛАСС ШЕСТИЛУЧЕВЫЕ КОРАЛЛЫ (HEXACORALLIA)
ОТРЯД АКТИНИИ, ИЛИ МОРСКИЕ ЦВЕТЫ (ACTINIARIA)
Если колонии гидроидов и горгонарий похожи на причудливые кусты и деревья, то крупные коралловые полипы актинии (Actiniaria) напоминают фантастические цветы. На многих языках они и называются морскими анемонами (см. цветную таблицу 9).
К отряду актиний относятся одиночные, лишь изредка колониальные животные, ведущие подвижный образ жизни. Только немногие глубоководные виды неподвижно прикреплены к субстрату. Актинии имеют цилиндрическую форму тела с уплощенным верхним (ротовой диск) и нижним концом (подошва). Но у некоторых актиний, в основном у тех, которые ведут роющий образ жизни, подошва может и не образовываться.
Число гастральных перегородок у большинства актиний по меньшей мере равно шести парам или кратно шести. Образование новых пар перегородок происходит почти всегда в промежуточных гастральных камерах. Однако от такого расположения перегородок встречаются отклонения, при которых число перегородок оказывается равным восьми или кратным восьми или десяти. Обычно такие отклонения особенно характерны для наиболее примитивных актиний. Известно, что в процессе индивидуального развития все актинии проходят стадию четырехлучевой симметрии, что, возможно, указывает на родство актиний с восьмилучевыми коралловыми полипами. Наибольшее сходство с современными восьмилучевыми кораллами у актиний из родa Edwardsia. Эти актинии ведут роющий образ жизни, обитая в илистых песчаных грунтах прибрежного мелководья. Их тело, на поверхности которого различимы восемь продольных валиков, имеет вытянутую червеобразную форму. Углубления между ними соответствуют восьми гастральным перегородкам. Кроме восьми полных перегородок, у старых экземпляров Edwardsia в верхней части тела образуются еще четыре, но уже неполные перегородки. Валики продольных мышечных тяжей лежат у этих актиний, так же как и у восьмилучевых кораллов, на вентральных сторонах перегородок. Восемь полных и восемь неполных перегородок образуются и у другой архаичной актинии Gonactinia. Наиболее хорошо известный европейский вид гонактинии Gonactinia prolifera имеет вид маленького, 2—3 мм в длину и 1—2 мм в ширину, прозрачного столбика с нежно-розовой или красной окраской. Ротовой диск актинии окружен шестнадцатью нежными щупальцами, расположенными в два ряда. Глотка ее столь коротка, что при открытом рте легко различимы в ее гастральной полости восемь основных радиальных перегородок. Гонактинии прикрепляются своей подошвой к субстрату, чаще всего к раковинам моллюсков, а иногда даже к стволам гидроидных полипов.
Число перегородок, кратное десяти, наблюдается у представителей семейства Myniadidae, весьма своеобразных актиний, перешедших к свободноплавающему образу жизни. Их поддерживает в воде особая воздушная камера, аналогичная пневматофору сифонофор, называемая пневмоцистой. Она образуется в результате сильного впячивания подошвы. Края подошвы при этом сближаются и смыкаются над центром углубления диска. Актиния поэтому плавает у поверхности воды ртом вниз. Как многие другие плавающие кишечнополостные, Myniadidae окрашены в голубой цвет. У остальных актиний число перегородок, как уже говорилось, равно шести парам или кратно шести.
Свободные края гастральных перегородок имеют мезентериальные нити, богатые железистыми и стрекательными клетками. У некоторых актиний образуются еще и особые нити — аконции, на которых стрекательные капсулы особенно многочисленны. Для защиты от нападения эти нити выбрасываются актиниями через рот или через особые отверстия в стенках тела или щупалец. Ротовой диск актиний окружен щупальцами. В зависимости от числа щупалец они располагаются в один или два и даже более концентрических рядов. В каждом круге щупальца бывают одного размера и формы, но щупальца, лежащие в разных кругах, часто довольно сильно отличаются друг от друга. Как правило, щупальца соответствуют пространствам между гастральными перегородками.Обычно щупальца имеют простую коническую форму, но иногда наблюдаются и значительные отклонения от нее. У некоторых видов на концах щупалец образуются вздутия благодаря тому, что там развиваются многочисленные батареи стрекательных капсул. У некоторых тропических мелководных актиний образуются ветвящиеся или перистые щупальца. На их концах образуются одна или две пары, служащие дополнительным средством для быстрого опорожнения полости тела.
Ротовое отверстие высших актиний имеет овальную или щелевидную форму. Глотка сильно сжата с боков и имеет два сифоноглифа. Только у описанных примитивных видов имеется лишь один слаборазвитый сифоноглиф или он вообще отсутствует. Биением ресничек сифоноглифа создаются два тока воды: один — направленный внутрь гастральной полости и приносящий кислород (у некоторых актиний— и частицы пищи), и другой — движущийся в обратном направлении и выносящий углекислый газ и продукты выделения.
Мышечная система актиний достигает высокого для кишечнополостных животных развития. Эктодермальная система складывается из продольных волокон, лежащих в щупальцах, и радиальных волокон вокруг ротового отверстия. Энтодермальная система состоит из кольцевой мускулатуры щупалец, ротового диска, глотки, стенок тела и ножного диска. На гастральных перегородках лежат продольные мышечные валики.
Нервная система актиний складывается из имеющихся во всех отделах тела эктодермальной сети нервных клеток и слаборазвитой энтодермальной сети, охватывающей только гастральные перегородки. Особенно много нервных клеток сконцентрировано у оснований щупалец и на ротовом диске. Однако это не приводит к образованию околоротового нервного кольца, так как нервные клетки расположены здесь очень рыхло. Другое скопление нервных клеток имеется около подошвы. Интересно отметить, что разные отделы тела оказываются особенно чувствительными к определенным раздражениям. Подошва, например, чувствительна к механическим раздражениям и не воспринимает химических. Ротовой диск, наоборот, весьма чувствителен к химическим раздражениям и почти не реагирует на механические. Пожалуй, лишь стенки тела и щупалец реагируют на механические, химические и электрические раздражения, но щупальца оказываются гораздо чувствительнее к ним, чем стенки тела.
ОТРЯД ЦЕРИАНТАРИИ (CERIANTHARIA)
Известным внешним сходством с актиниями обладают представители отряда цериантарий.
Цериантарии — малочисленная и сравнительно плохо изученная группа шестилучевых кораллов. К ним относятся исключительно одиночные бесскелетные кишечнополостные, ведущие закапывающийся образ жизни. Длинное цилиндрическое тело цериантарий мускулистое и совершенно гладкое. На переднем его конце имеется расширенный ротовой диск, задний конец заострен и снабжен порой, не играющей, однако, роли порошицы. Ротовой диск окружен двумя группами щупалец, между которыми остается небольшое свободное пространство. В каждой группе щупальца могут располагаться в одном или четырех концентрических кругах. Размеры щупалец в пределах каждой группы одинаковые, но могут быть весьма различными в разных группах. Чаще всего более длинные щупальца образуются в наружной группе. В целом внешний вид цериантарий почти не отличается от внешнего вида многих актиний, ведущих роющий образ жизни. По внутреннему же строению цериантарии резко отличаются от актиний.
На поперечном разрезе животного на уровне глотки отчетливо видны многочисленные и всегда полные гастральные перегородки, расположенные на равном расстоянии друг от друга. Однако радиальная симметрия нарушается сдавленной с боков глоткой с одним сифоноглифом, лежащим не на брюшной, как у остальных коралловых полипов, а на спинной стороне. Валики продольной мускулатуры перегородок выражены слабо, тем не менее видно, что они лежат на их вентральной стороне. Все новые перегородки всегда образуются на вентральной стороне тела.
Свободные края перегородок, кроме направляющих, заканчиваются мезентериальной нитью, снабженной железистыми клетками и стрекательными капсулами.
Мышечный слой цериантарий сильно развит и лежит сразу под эктодермой. Мезоглеальный слой представлен тонкой гомогенной пластинкой.
Цериантарии не имеют скелета. Обыкновенно они сидят более или менее отвесно в грунте, в трубках, образованных из затвердевающей слизи, выделяемой многочисленными железистыми клетками эктодермального слоя. Внутренняя поверхность трубки совершенно гладкая и довольно скользкая, наружная же поверхность становится грубой и шероховатой на ощупь из-за обилия различных посторонних включений — песчинок, обломков раковин моллюсков, игл губок и раковинок фораминифер. Трубки цериантарий несколько возвышаются над грунтом. В активном состоянии животное высовывает из трубки свой верхний конец. Щупальца наружной группы обычно расправляются и лежат в одной плоскости. Более короткие щупальца внутренней группы поднимаются вверх. При малейшем раздражении животное быстро втягивается внутрь своего домика. Длина трубок у разных видов весьма изменчива — от нескольких до 30—40 см. Но известны трубки, достигавшие 1 м в длину. Цериантарии не способны втягивать щупальца внутрь своего тела. Прячась в трубку, животное складывает щупальца над своим ротовым диском.
Все цериантарии — гермафродиты, однако мужские и женские половые клетки образуются у них в разное время. Как правило, этим животным свойствен протандрический гермафродитизм. Как протекает размножение цериантарий, до сих пор неизвестно.
Бесполое размножение у цериантарий наблюдается редко, но зато эти животные обладают исключительной способностью к регенерации.
Цериантарии обитают во всех морях с нормальной соленостью. В таких морях, как Черное и Балтийское, эти животные не встречаются. Большинство из 50 известных в настоящее время видов цериантарий обитает в теплых водах тропического пояса и в субтропиках. Лишь немногие виды проникают в холодные воды приполярных районов. Диапазон вертикального распределения цериантарий довольно широк.
Образ жизни цериантарий изучен мало. Многие планктонные личинки их питаются мелкими рачками — копеподами. Взрослые же животные используют в пищу органические частицы, взвешенные в придонном слое воды.
ОТРЯД ЗОАНТАРИИ (ZOANTHARIA)
Зоантарии представляют собой очень небольшую группу кишечнополостных, иногда одиночных, иногда колониальных, лишенных скелета и напоминающих внешним своим видом мелких актиний.
У колониальных зоантарий отдельные полипы объединяются, как у альционарий, или мембрановидной, стелющейся по субстрату пластинкой, или при помощи сети коротких отростков.
Расположение гастральных перегородок отличается от их расположения у всех современных коралловых полипов. Однако внутреннее строение зоантарий отдаленно напоминает строение вымерших четырехлучевых кораллов. Все полные перегородки расположены парами. Между полными перегородками образуются короткие неполные перегородки. Имеются две пары направляющих перегородок, из которых дорзальная состоит из неполных сект, достигающих глотки. В каждой паре гастральных перегородок мышечные валики продольной мускулатуры обращены друг к другу, за исключением направляющих, где они лежат на противолежащих сторонах.
Зоантарии не имеют собственного скелета. Эктодермальные клетки поверхности тела способны выделять лишь довольно толстую кутикулу. Любопытно, что эти животные способны укреплять кутикулу при помощи всевозможных мелких чужеродных предметов — песчинок, игл губок, раковин фораминифер и т. д.
Мышечная система зоантарий развита очень слабо, особенно в базальной части полипов. О нервной системе до сих пор почти ничего не известно.
Эти животные чаще раздельнополы. В таких случаях и колонии слагаются только из мужских или только из женских особей. Но имеются и гермафродитные особи зоантарий. Развитие зоантарий также еще полностью не прослежено, известны лишь их своеобразные плавающие личинки.
Зоантарии распространены всесветно, однако это сравнительно редкие животные и попадаются они нечасто. Из 300 известных в настоящее время видов большинство обитает в тропической зоне. Только отдельные представители зоантарий проникли далеко на север и юг, встречаясь в холодных водах Арктики и Антарктики. Среди зоантарий имеются обитатели прибрежного мелководья и больших глубин океана. Однако в абиссаль океана на глубину более 3 тыс. м они не смогли проникнуть.
Зоантарии ведут исключительно донный образ жизни. Одиночные формы живут на песчаном или заиленном грунте, погружая в него задний конец своего тела.
ОТРЯД АНТИПАТАРИИ (ANTIPATARIA)
Имеется еще одна группа кишечнополостных, внешне напоминающих растения, — антипатарии.
Это изящные перистые, вееровидные, древовидные колонии и колонии, напоминающие ершики. Подобно роговым кораллам, они имеют осевой роговой скелет, но, в отличие от роговых кораллов, полностью лишенный извести.
По главному стволу скелета и по его боковым отросткам тянутся каналы, а поверхность его сплошь покрыта коническими шипами, иногда достигающими значительной высоты. Шипы могут отсутствовать у основания колонии или на концах молодых ветвей. Все ветви колонии равномерно покрыты тонким, лишенным окраски ценосарком, в котором образуются мелкие невзрачные, небольшого размера, удивительно некрасивые и всегда бесцветные полипы. Вокруг их ротового отверстия образуется всего лишь шесть коротких толстых щупалец. Только у представителей одного рода — Dendribranchia — образуется восемь к тому же перистых щупалец. У некоторых антипатарий полипы как бы расстилаются вдоль осевых стержней колонии, и тогда они подразделяются на три отдела: в нижнем и верхнем обычно развиваются половые продукты, в середине среднего отдела образуется ротовой конус с ротовым отверстием на его вершине. Каждому отделу соответствует одна пара коротких щупалец. Гастральных перегородок в теле полипа может быть 6, 10 или 12. Чаще бывает 10 перегородок. Шесть перегородок первичные, остальные представляют собой вторичное образование. Глотка овальной формы. Все перегородки полные, т. е. достигают глотки. Полости щупалец являются продолжением боковых камер полости тела, образованных первичными перегородками.
Тонкая цененхима, а иногда и тело полипов часто пронизывается почти насквозь острыми шипами скелета.
Мускулатура у антипатарий почти совсем не развита. Ничего не известно о строении нервной системы. Полипы антипатарий раздельнополы, но на одной и той же колонии могут развиваться как женские, так и мужские особи.
Питание антипатарий совершенно не изучено. В их гастральной полости не удавалось найти следов схваченной и переваренной пищи. Однако эктодермальный слой тела полипов имеет многочисленные реснитчатые клетки, и это позволяет предполагать, что антипатарии питаются органическими частицами, взвешенными в морской воде и переносимыми в глотку биением ресничек. Биология антипатарий почти совсем неизвестна.
Из 120 видов антипатарий около 100 видов обитает на небольших глубинах тропического и субтропического поясов океана. Очень немногие виды смогли проникнуть в умеренные воды и на большие глубины океана. Глубже 2 тыс. м встречается 5 видов антипатарий и глубже 3 тыс. м — только три вида. Глубоководные виды антипатарий распространены во впадинах всего Мирового океана.
ОТРЯД МАДРЕПОРОВЫЕ КОРАЛЛЫ (MADREPORARIA)
Мадрепоровые кораллы — самая обширная группа кишечнополостных. Она насчитывает свыше 2500 видов, обитающих исключительно в море. Среди них имеются как одиночные, так и колониальные формы, но последних подавляющее большинство. Самая характерная черта кораллов этого отряда — наличие мощного известкового скелета, образуемого эктодермой. Скелет мадрепоровых кораллов сплошной, т. е. он не состоит из отдельных элементов — спикул или игл. Подошва полипа образует скелетную пластинку, от которой поднимаются вверх (внутри септ) радиальные скелетные перегородки. Толща скелета постоянно наращивается, и у сформированного полипа живые мягкие ткани (по сравнению с толстым скелетом) выглядят как полупрозрачная пленочка. У колониальных форм скелетные чашечки отдельных полипов сливаются в единый скелет колонии. Полипы одиночных видов мадрепоровых кораллов могут достигать довольно большой величины. Так, диаметр подошвы некоторых видов грибовидных кораллов Fungia у наиболее крупных особей бывает до 25 см. Напротив, полипы колониальных форм обычно очень маленькие, как правило, всего 1—5 мм в диаметре. Форма колоний, образуемых мадрепоровыми кораллами, может быть очень разнообразной: одни виды образуют кустики, другие — плоские решетчатые плиты, третьи — похожи на цветную капусту, имеются и шаровидные колонии. Во многом форма колонии этих кораллов зависит главным образом от силы прибоя, течений, заиленности и температуры воды.
Многие колониальные мадрепоровые кораллы проявляют отчетливую тенденцию к объединению колонии в единый организм путем слияния полипов. Рассмотреть это явление можно на примере некоторых шаровидных колоний. Так, колония фавии (Favia) имеет вид полушария с ячеистой поверхностью. Каждая ячейка — это чашечка полица. Полип сидит внутри ее и похож на маленькую актинию — в его центре имеется рот, окруженный венчиком щупалец. У некоторых видов цейлорий (Coeloria) в чашечке сидит уже не один, а два-три полипа. Такая чашечка имеет не круглую, а вытянутую форму, рты полипов расположены в один ряд и окружены все вместе одним венчиком щупалец. У этого коралла уже трудно установить, какое щупальце принадлежит какому полипу. У лобофиллий (Lobophyllia) в ряд сидят уже десятки полипов, причем границы между отдельными полипами утрачиваются еще больше, так как у некоторых соседних особей рты сливаются в общую щель. Наконец, у кораллов из рода меандрин (Meandrivia) все полипы сливаются воедино. Колония имеет вид полушария, по поверхности которого проходят причудливо извитые борозды, что придает кораллу вид мозга (он и называется мозговиком). По центру борозды проходит сплошная щель, образовавшаяся из слившихся ртов полипов, а по сторонам этого единого рта колонии сидят рядами щупальца. У мозговиков вообще невозможно установить границы полипов — вся колония превратилась в единый организм.
Мадрепоровые кораллы обитают во всех зонах Мирового океана и на различных глубинах; отдельные виды встречаются в полярных и умеренных поясах. Но наибольшего разнообразия эти кораллы достигают только в тропиках, где они составляют основу знаменитых коралловых рифов.
КОРАЛЛОВЫЕ ПОСТРОЙКИ
Коралловыми рифами называют массовые поселения (заросли) морских организмов, обладающих твердыми известковыми скелетами. Основу такого рифа составляют мадрепоровые кораллы, но далеко не всегда они являются его единственными создателями. Наряду с мадрепоровыми здесь могут быть и мягкие кораллы — альционарии, и гидрокораллы, и мшанки, и даже известковые водоросли.
Кроме того, существует большое количество прикрепленных полихет, моллюсков и усоногих раков, также обладающих мощным известковым скелетом и участвующих в построении рифа. Однако главенствующая роль все же остается за мадрепоровыми кораллами.
Они являются не только самыми многочисленными, но, что гораздо более важно, непременными компонентами кораллового рифа, тогда как представители любой из других перечисленных выше групп животных могут отсутствовать или встречаться в небольшом числе экземпляров. В связи с этим процветание жизни на коралловом рифе в первую очередь зависит от процветания самих мадрепоровых кораллов.
Отдельные виды мадрепоровых кораллов встречаются в различных частях Мирового океана и на различных глубинах, но далеко не везде кораллы образуют обширные поселения, которые получили название рифов. Для роста и процветания кораллового рифа необходимо наличие целого ряда благоприятных условий. Все рифообразующие кораллы нуждаются в морской воде с нормальной океанической соленостью, т. е. с содержанием в ней около 3,5% солей. Кораллы могут вынести некоторое повышение солености (так, они хорошо развиваются в водах Красного моря, соленость которого достигает 3,8—4%), но всякое уменьшение количества солей в морской воде действует на мадрепоровые кораллы губительно. В связи с этим они не поселяются вблизи устьев крупных рек, где вода распреснена. Во время сильных дождей, принимающих характер стихийного бедствия, когда речной сток усиливается и соленость в прибрежных частях моря падает, наблюдается массовая гибель мадрепоровых кораллов. Особенно боятся кораллы прямого попадания на них дождевой воды, что иногда случается, если дождь идет во время отлива. Известно много случаев, когда обильные дожди вызывали гибель мадрепоровых кораллов, а вместе с ними и всего населения рифа. Последний раз массовая гибель мадрепоровых кораллов в результате сильнейших ливней наблюдалась в 1956 г. на некоторых участках восточного побережья Австралии. Гибель кораллов влечет за собой серьезные последствия для различных обитателей моря. Разлагающиеся мягкие ткани кораллов отравляют воду и вызывают смерть тех морских животных, которые уцелели при распреснении. Даже после того как море похоронит их трупы и начисто вымоет скелеты кораллов, еще долго такие участки остаются безжизненными. Большинство животных, населяющих коралловый риф, либо питается кораллами, либо находит в их зарослях свое убежище, либо получает и то и другое. Поэтому, пока заново не разрастутся кораллы, жизнь здесь замирает.
Другим не менее важным условием для жизни рифообразующих кораллов является высокая и постоянная температура морской воды. Установлено, что коралловые рифы встречаются лишь там, где температура воды никогда не падает ниже 20,5° С. В связи с этим распространение коралловых рифов ограничено тропическими частями Тихого, Индийского и Атлантического океанов. На карте распространения коралловых рифов (см. цветную таблицу 10) почти полностью совпадают границы зоны, где встречаются коралловые рифы, с изотермой 20° С. Обращает на себя внимание то обстоятельство, что в Атлантическом океане зона рифов у берегов Америки значительно больше, чем на африканском побережье. То же самое наблюдается и в Тихом океане. В его западной части зона рифов тянется от юга Японии до Австралии и включает всю северную половину ее береговой линии, тогда как на тихоокеанском побережье Америки эта зона расположена в относительно короткой полосе между тропиком Рака и экватором и совершенно не заходит в южное полушарие. Это обстоятельство не случайно, оно объясняется особенностями океанических течений. Вдоль экватора с востока на запад круглый год движутся воды поверхностного экваториального течения. Они сильно нагреваются под лучами тропического солнца. Конечно, тропическое течение не имеет вида цельного кольца: оно разбивается на три дуги материками Африки, Америки и островами, расположенными между Азией и Австралией. Дойдя до суши, дуга течения разбивается на два рукава, один из которых несет теплые воды на север, а другой на юг. Это происходит и в Тихом, и в Индийском, и в Атлантическом океанах. Теплые течения обогревают восточные берега всех материков, в то же время их западное побережье охлаждается. Это происходит вследствие подхода к экватору с севера и с юга вдоль западных берегов более холодных струй, восполняющих убыль воды, уносимой экваториальным течением. Разница между поселениями кораллов на западном и восточном побережье Индийского океана выражена меньше, так как в этом океане нет притока холодных вод с севера.
Температура воды и недостаток освещения ограничивают распространение кораллов также и по вертикали. В связи с этим рифообразующие кораллы не заходят на глубину более 50 м.
Вертикальное распределение многих мадрепоровых кораллов ограничивается также и условиями освещенности. В теле большинства таких кораллов поселяются микроскопические одноклеточные жгутиковые из отряда перидиней (Peridinea), так называемые зооксантеллы, окрашивающие живые ткани кораллов в зеленоватый или желтоватый цвет. До последнего времени предполагали, что кораллы могут получать дополнительное питание, переваривая эти водоросли. Какое преимущество получают водоросли, поселяющиеся в теле кораллов, оставалось совершенно неясным. Известно, что такие же одноклеточные водоросли поселяются также в теле некоторых других тропических морских животных, в частности они всегда имеются в тканях двустворчатых моллюсков тридакна (Tridacna), обитающих среди зарослей кораллов. Большинство кораллов образует с набивающими их ткани зооксантеллами очень сложное физиологическое целое, загадка которого еще не разрешена полностью.
Недавно шотландский зоолог Йонг (С. М. Yonge, 1958) дал более полное объяснение причин симбиоза между кораллами и зооксантеллами. Несомненно, что зооксантеллы оказывают существенное общее стимулирующее действие на метаболизм своих хозяев-кораллов и для некоторых из них имеют очень большое значение в процессе питания. Шестилучевые кораллы, которые, как и все кишечнополостные, являются хищниками, совершенно не способны усваивать пищу растительного происхождения. При любых неблагоприятных условиях (голодании, недостатке кислорода или понижении температуры внешней среды) в их тканях происходят серьезные болезненные изменения, при этом зооксантеллы гибнут и изгоняются из тела коралла. Вначале они попадают в кишечную полость, а затем через ротовое отверстие выводятся наружу. Однако даже в случае заболевания, вызванного продолжительным голоданием, зооксантеллы совершенно не усваиваются кораллами, хотя и попадают в кишечную полость, в которой происходит пищеварение. Если поместить коралл в темноту, то зооксантеллы также гибнут, хотя некоторые кораллы при этом продолжают нормально развиваться.
Установлено также, что альциониевые кораллы (Alcyonaria), обильно заселенные зооксантеллами, утрачивают свободное питание, не реагируют на животную пищу, некоторые части их пищеварительной полости испытывают редукцию, и в темноте они погибают. Видимо, зооксантеллы служат для них источником питания.
На основании этих наблюдений Йонг делает следующий вывод. Зооксантеллы потому поселяются в теле кораллов (и некоторых других тропических морских животных), что они получают здесь углекислоту (как продукт дыхания кораллов), которую используют в процессе фотосинтеза углеводов. Еще более важной следует считать возможность использования зооксантеллами других продуктов обмена, образующихся в результате жизнедеятельности кораллов, в первую очередь производных азота и фосфора. Эти вещества зооксантеллы используют при синтезе белков — протеинов. Таким образом, зооксантеллы как бы заменяют кораллам органы выделения, аккумулируя в себе продукты распада. В свою очередь кораллы благодаря наличию в их теле зооксантелл получают в дневные часы дополнительный кислород, образующийся в результате фотосинтеза. Йонг считает, что отдельные колонии кораллов вполне могут существовать и без зооксантелл, но наличие этих симбионтов совершенно необходимо для образования мощных зарослей — рифов, так как избыток кислорода и интенсивное удаление продуктов обмена из тела кораллов дают им преимущества для быстрого роста и развития мощного скелета.
Зооксантеллы могут развиваться только при достаточном освещении. Поэтому мадрепоровые кораллы даже при благоприятной температуре не опускаются на такую глубину, куда проникает мало света; они редко поселяются также в затененных местах — нишах и щелях. Однако виды рифообразующих мадрепоровых кораллов, в теле которых нет зооксантелл, напротив, предпочитают укрытые, затененные места рифа, таков киноварно-красный коралл Dendrophyllia.
В последнее время в связи с возможностью применения новых методов изучения процессов, происходящих в организме, выдвинуто новое объяснение роли зооксантелл для кораллов.
Горо (Goreau, 1963) с помощью радиоактивного изотопа углерода исследовал биохимическую сторону механизма отложения извести кораллами. Им была установлена прямая зависимость между фотосинтезом зооксантелл и скоростью отложения извести. Оказалось, что на свету отложение извести идет в 9 раз быстрее, чем в темноте. Уэнрайт (Wainwright, 1963) добавляет, что зооксантеллы вырабатывают хитиноподобное вещество, которое служит основой для отлагающейся извести скелета. Горо считает, что кораллы, поселяющиеся в затененных местах и на некоторой глубине, куда попадает меньше света, потому обладают более нежным скелетом, что фотосинтез здесь замедлен.
Следующим, важнейшим условием, способствующим нормальной жизнедеятельности рифообразующих кораллов, является чистота и прозрачность морской воды. Мадрепоровые кораллы предпочитают прозрачную воду не только потому, что она лучше пропускает солнечные лучи, несущие тепло и свет, но и потому, что они не выносят попадания на свое тело частиц ила. Особенно это касается видов, способных образовывать сплошные густые заросли. Для них заиление не менее губительно, чем понижение температуры (ниже 20°) или распреснение морской воды. Тем не менее в ряде случаев и в заиленных местах (в бухтах, лагунах) встречаются значительные поселения мадрепоровых кораллов. Однако они никогда не образуют здесь густых зарослей: обычно в таких случаях колонии кораллов отстоят друг от друга на известном расстоянии, их видовой состав значительно отличается от видового состава кораллов, обитающих в незамутненной воде.
Если колонию мадрепоровых кораллов поместить в сосуд с чистой теплой морской водой, то она все равно через короткий срок неизбежно погибнет вследствие недостатка кислорода. Дело в том, что с повышением температуры растворимость газов в воде понижается; кораллы же очень быстро поглощают из воды растворенный кислород и, если воду не сменить, погибают. Вот почему в природе наибольшего развития кораллы достигают в прибойных участках моря, где обогащение воды кислородом облегчено, а сами кораллы постоянно омываются все новыми порциями воды.
Наконец, для роста кораллов, как и всех других животных, требуется пища. Мадрепоровые кораллы питаются микроскопическими животными, населяющими морской планктон: простейшими и различными мелкими ракообразными. Новые порции пищи также приносятся к рифу морем.
Казалось бы, немного можно найти мест, где все эти условия были бы в наличии одновременно. И тем не менее это не так. Значительные пространства тропической части Мирового океана оказываются вполне благоприятными для процветания мадрепоровых кораллов. Общая площадь всех коралловых сооружений превышает 27 млн. кв. км. Площадь одних только островов и обнажающихся при отливе рифов равна приблизительно 8 млн. кв. км, т. е. она несколько более площади Австралии, равной 7,7 млн. кв. км. Уже одно это говорит о большом значении коралловых поселений, требующих их всестороннего изучения.
Коралловые рифы представляют собой как бы насыщенные жизнью оазисы среди малопродуктивных вод тропического океана. Вычисления показали, что они дают 1500—3500 г первичной продукции углерода на 1 кв. м в год. Это в 100 раз больше, чем в окружающем океане, и в 4—8 раз больше, чем в морях умеренной зоны (не считая ее самой прибрежной части).
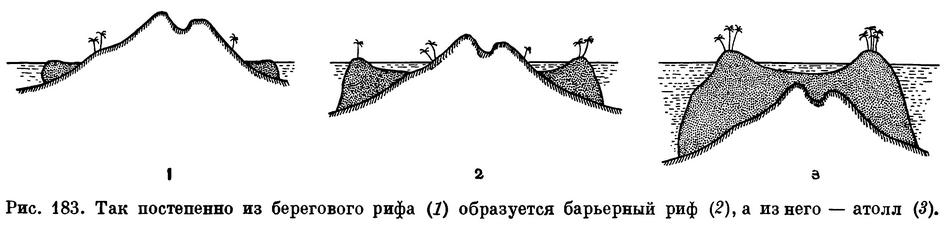
Все многообразие построек, создаваемых кораллами и другими рифообразующими организмами, можно разделить на несколько основных типов. Различают береговые рифы, расположенные непосредственно по берегам островов или материков, барьерные рифы, отстоящие от берега на некотором расстоянии, и атоллы — кольцеобразные коралловые острова.
Наконец, имеются также значительные коралловые банки, которые возникают на отмелях, и густые коралловые поселения в лагунах и бухтах. Процесс формирования всех этих коралловых построек интересовал геологов и зоологов уже очень давно, особенно непонятным казалось происхождение кольцевидных островов — атоллов. Было предложено несколько теорий, объясняющих образование этих островов, причем многие из них довольно наивны. Так, до середины прошлого века господствовало предположение, что атоллы являются коралловыми обрастаниями кратеров подводных вулканов.
Первая убедительная теория происхождения коралловых построек различных типов была выдвинута крупнейшим натуралистом прошлого века Чарлзом Дарвином. В своей книге «Строение и распространение коралловых рифов», вышедшей в 1842 г., Дарвин дал не только подробнейшее описание различных коралловых построек, но и показал, как один тип коралловых поселений по мере развития переходит в другой.
Дарвин собрал огромный материал, касающийся особенностей жизнедеятельности организмов, образующих коралловый риф, их отношения к условиям внешней среды, интенсивности роста и распространения в Мировом океане. Часть сведений он получил от капитанов кораблей, плававших в тропических морях, и от ученых, занимающихся изучением кораллов. Ценнейшие наблюдения провел сам во время кругосветного путешествия на «Бигле».
По мнению Дарвина, первой стадией образования коралловых островов является окаймляющий береговой риф. Кораллы в этом случае используют берега островов в качестве опоры, или, как говорят специалисты, субстрата. Если условия благоприятствуют развитию кораллов, а остров не испытывает подъема или опускания, риф так и остается окаймляющим береговым рифом. В тех случаях, когда дно моря в результате процессов, происходящих в земной коре, начинает подниматься и остров как бы выходит из воды, окаймляющий риф разрастается вдоль его новой береговой линии. Участки рифа, которые оказываются поднятыми над уровнем моря, погибают, а с морской стороны риф нарастает и увеличивается. Но общая картина при этом не меняется.
Совсем иначе обстоит дело в тех случаях, когда морское дно опускается и остров погружается в воду. Как уже указывалось выше, рифообразующие организмы требуют для своего развития много пищи и чистой морской воды, богатой кислородом. Благодаря этому нарастание рифа всегда идет по его периферии, омываемой морем. Вследствие этого между растущим внешним краем берегового рифа и погружающимся островом вскоре образуется затопленное водой пространство, на котором кораллы растут менее интенсивно. Так возникает барьерный риф. Чем дольше длится этот процесс, тем дальше барьер отступает от острова. Наконец, может наступить такой момент, когда остров окончательно погружается в море, а барьерный риф превращается в атолл — кольцевой остров с заключенной внутри его лагуной.
Современные коралловые постройки дают возможность проследить все стадии постепенного превращения берегового рифа в атолл. По мере дальнейшего опускания морского дна кольцо атолла постепенно расширяется, причем с его наружной стороны сразу начинается крутой свал. Внутренняя лагуна обычно бывает мелководной, так как она все время зарастает менее прихотливыми, но и медленно растущими видами кораллов. Скорость роста различных кораллов неодинакова. Массивные виды, как, например, мозговики, нарастают в год на 1—2 см; ветвистые формы, если ничто не препятствует их развитию, дают за это же время ветки длиной 20—30 см. Вероятно, внутренние возможности роста кораллов более значительны, чем это допускается внешними условиями. Так, рост рифа вверх резко ограничивается уровнем моря; но как бы быстро ни опускалось морское дно, кораллы всегда успевают разрастись до той верхней границы, при которой они могут существовать. Часть современных коралловых островов возвышается со дна моря на очень значительную высоту. Попытки бурить, с тем чтобы определить, как глубоко идет известковая порода, показали, что даже на глубине 50—70 м коралловые острова сложены из известняка.
Позднее некоторые исследователи выдвинули новые теории происхождения коралловых островов. Дело в том, что теория Дарвина базируется на одном непременном условии — опускании морского дна. Между тем противники этой теории утверждают, что отношения между поднятиями и опусканиями морского дна в действительности несравненно сложнее, чем это полагал Дарвин.
В связи с этим в 1880 г. Джон Мэррей (Murray) выдвинул новое предположение о происхождении атоллов. Мэррей полагал, что над подводными возвышенностями (которые могут возникнуть благодаря вулканической деятельности) жизнь гораздо богаче. Здесь на отмелях поселяются различные донные морские организмы, а в воде над ними развивается более богатый планктон. Отмирая, все эти животные оставляют много скелетных остатков, благодаря чему напластование осадков на такой возвышенности идет значительно быстрее, чем на более глубоких участках морского дна. Когда вершина такого подводного бугра приблизится к поверхности моря на 50 м, на ней начинают поселяться мадрепоровые кораллы. Они растут вверх и образуют коралловую банку. Последняя нарастает по периферии, а ее внутренние части разрушаются, благодаря чему и образуется кольцо атолла. Хотя теория Мэррея на некоторый срок получила признание, но она не объясняла происхождения барьерных рифов и была забыта.
Позднейшие исследователи опять вернулись к теории Дарвина, но пытались внести в нее некоторые дополнения. Очень остроумное решение вопроса предложил американский ученый Дели (Dely, 1915).
Он совершенно правильно рассудил, что для теории Дарвина нет особой разницы, опускается ли морское дно, или повышается общий уровень воды в океане. Предложенная Дели «теория ледникового контроля» основывается на том, что в последнюю ледниковую эпоху значительные массы воды вышли из кругооборота и остались на континентах в виде льда. В это время наблюдалось общее похолодание на всем земном шаре и условий для произрастания рифообразующих кораллов не было. С наступлением потепления в тропических частях моря появились и теплолюбивые кораллы, которые селились на отмелях и вокруг островов. Потепление вызвало таяние ледников, что повлекло за собой повышение уровня Мирового океана примерно на 70 м. Именно на эту глубину залегают достоверные остатки скелетов мадрепоровых кораллов.
Теория Дели, поддержанная известным исследователем коралловых рифов Девисом (Davis, 1928), в значительной мере укрепляет теорию Дарвина, хотя и вносит в нее существенные изменения. Однако и «ледниковый контроль» не дает исчерпывающего объяснения происхождению коралловых островов. В последнее время появилось много новых данных, касающихся геоморфологии, океанографии, палеонтологии и биологии современных рифообразующих животных. Все они используются для уточнения дарвиновской теории, но в основе наших современных представлений о происхождении и развитии коралловых сооружений остается гениальное творение Чарлза Дарвина.
Совсем недавно теория Ч. Дарвина о происхождении и развитии коралловых островов получила новое подтверждение.
Известный английский исследователь кишечнополостных Вильям Рисс (Riss, 1962) сообщил, что проведенные сейсмические исследования четырех атоллов полностью подтвердили все основные положения дарвиновской теории.
Внешне все коралловые острова очень сходны между собой. При подходе к такому острову уже издали становятся видны ряды кокосовых пальм, белая полоса прибрежного пляжа и буруны на краю рифа.
Коралловые острова обычно очень незначительно поднимаются над уровнем моря, их растительность довольно однообразна: кроме кокосовых пальм, здесь растут высокие кусты пандануса. Листья этого растения широкие и длинные, снабженные по краям многочисленными очень острыми зазубринами-шипами. На кустах висят плоды, похожие по цвету, величине и форме на ананас. Ближе к берегу можно видеть высокую жесткую траву и мясистые опунции, похожие на некоторые виды кактусов. Вся эта растительность удовлетворяется скудной почвой и может обходиться минимальным количеством пресной воды, которая выпадает во время редких дождей.
Пляж резко отграничен от полосы зеленых растений, он состоит почти исключительно из кораллового песка — перемолотых волнами скелетов мадрепоровых кораллов, но здесь же имеются и раковины морских простейших фораминифер, и обломки раковин моллюсков. Часто попадаются целые, обмытые морем раковины, многие из них имеют очень красивую форму, ярко окрашены, блестят перламутром. На поверхности песка бегают жуки-скакуны, в самом песке роют норы крабы оципода (Ocypoda). Это очень осторожные животные, укрывающиеся в нору при малейшей опасности. Если такого краба застать вдали от норы, он стремительно несется на высоко поднятых ногах по пляжу и с разбегу смело бросается в набегающую волну. Остановившись на мелководье, оципода высовывает из воды глаза, сидящие на длинных стебельках, как бы выполняющих роль перископа.
Ширина пляжа обычно не превышает 10—20 м, его верхняя часть только во время самого сильного прилива обдается брызгами морской воды, тогда как нижняя часть пляжа заливается приливной волной очень регулярно. В приливо-отливной части пляжа поселяются различные роющие двустворчатые моллюски: донаксы (Donax) и атактодея (Atactodea), здесь живут также ползающие брюхоногие моллюски клипеоморус (Clypeomorus), планаксис (Planaxis), мелкие раки-отшельники и некоторые виды полихет.
Ниже расположен отмерший участок рифа, состоящий из кораллового известняка, покрытого многочисленными мелкими и крупными обломками кораллов. Здесь в щелях и под кустами мертвых полипняков поселяются змеехвостки макрофиотриксы (Macrophiotrix), гигантские полихеты эунице (Eunice aphroditoides), достигающие более 1 м в длину. В толще известняка обитают животные, сверлящие и разрушающие скелет мертвых кораллов. Среди них наиболее активны небольшие двустворчатые моллюски литофага (Lithophaga) и гастрохена (Gastrochaena), а также маленькие морские ежи эхинометра (Echinometra matthaei). Отмерший участок рифа покрыт различными водорослями, которые служат пищей некоторым его обитателям. Гигантские полихеты эунице далеко высовываются из своих убежищ и пожирают водоросли. При малейшей опасности они быстро скрываются в щели.
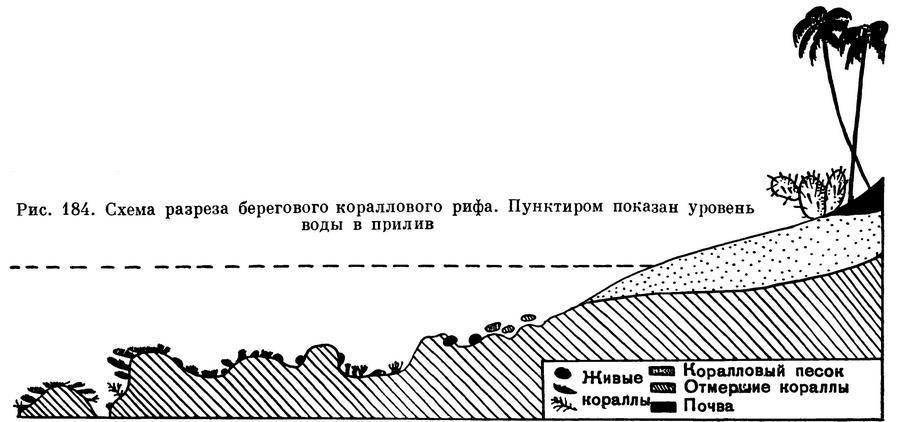
Если продвигаться далее в сторону моря, можно увидеть первые колонии живых мадрепоровых кораллов. Ближе всего к берегу селятся буро-зеленые массивные шаровидные или уплощенные колонии поритес (Porites), фавитес (Favites) и орбицелла (Orbicella). Они относятся к разным семействам, но имеют одну общую особенность: полипы этих видов могут глубоко втягиваться внутрь массивных колоний и потому способны переносить непродолжительное обсыхание (этот участок рифа во время отлива на некоторый период остается не покрытым водой). Отдельные виды живущих здесь кораллов не выносят обсыхания, в связи с чем у них выработалась своеобразная форма колонии, позволяющая запасать воду на время отлива. Внешне такая колония больше всего похожа на чашу или бокал. Когда уровень воды при отливе понижается, в полости чаши остается вода. Все полипы таких колоний обращены внутрь и потому постоянно находятся в водной среде. Это приспособление выработалось у некоторых видов турбинарий (Turbinaria) и подобаций (Podobacia). Колонии этих же видов при поселении в более глубоких местах, где они не подвергаются обсыханию, имеют плоскую или даже несколько выпуклую форму. В этой зоне рифа обитают многочисленные брюхоногие моллюски — ципрея (Cypraea) и конус (Conus), много змеехвосток, голотурий, раков-отшельников и крабов. Иногда попадаются большие брюхоногие моллюски птероцера (Pterocera), несущие на раковине длинные изогнутые шипы.
На краю рифа, где даже во время самого низкого отлива бьются волны прибоя, кораллы образуют сплошные многоярусные заросли. Волны наката достигают здесь огромной силы, они обрушиваются на риф и разбиваются о него в белую пену. Действие постоянно движущихся масс воды откладывает на все коралловое поселение края рифа глубокий отпечаток. Кроме волн, здесь все прочно и неподвижно. Обломки кораллов, раковины мертвых моллюсков прочно заклинены в щелях: все подвижные предметы неминуемо уносятся волнами. Они либо выбрасываются на более высокие участки рифа, либо смываются в море. Колонии кораллов здесь плотно срастаются с основой рифа и между собой. Одни из них имеют вид гигантских шаров, другие комковатые, есть колонии плоские, как лепешки. Для этой части рифа наиболее типичны кораллы-мозговики (Coeloria и Meandra), шаровидные гладкие пориты (Porites), орбицеллы (Orbicella), колючие галаксеи (Galaxea), симфиллия (Symphyllia), лобофиллия (Lobophyllia). Ветвистые формы имеют короткие толстые ветви. Все они не боятся разрушительного действия волн, так как обладают толстым и очень прочным известковым скелетом. Другие виды кораллов, например некоторые представители рода акропор (Acropora), приспособились к жизни в прибойной полосе иначе. Они имеют вид огромных округлых плит и похожи на гигантские грибы-трутовики. Эти кораллы растут в несколько ярусов по наружному склону рифа. Диаметр таких плит достигает 2—5 м. Вся толща плиты пронизана многочисленными отверстиями и напоминает собой решето. Набегающие и уходящие волны свободно проходят сквозь отверстия, не разрушая колонии. Все колонии кораллов, поселяющиеся на краю рифа, окрашены в яркие, сочные цвета. Благодаря наличию в тканях полипов симбиотических водорослей зооксантелл большинство видов кораллов желтоватой или зеленой окраски. Цвет, особенно зеленый, очень сочный, похожий на цвет молодой весенней травы. Края ситовидных плит акропор, как более молодые участки колонии, окрашены светлее. Между бурыми и зелеными кораллами яркими пятнами выделяются карминно-красные, иногда вишнево-красные колонии поциллопор (Pocillopora). Здесь же поселяются коричневые с голубым или сиреневым налетом Montipora foliosa, колония которых по форме напоминает цветную капусту. В темных щелях сидят маленькие колонии коралла Dendrophyllia киноварно-красного цвета.
Если опуститься в воду по внешнему краю рифа на глубину 3—4 м, взору представится совершенно необыкновенный подводный лес. На этой глубине сила прибойных волн несколько ослабевает, поэтому здесь поселяются более нежные ветвящиеся колонии, причем преобладают представители многообразного рода акропор (Acropora). Здесь есть колонии, похожие по форме на елочки, густые колючие кусты, грибы-трутовики, имеются и различные кораллы, причудливая форма которых трудно поддается описанию. Преобладают яркие краски: лимонно-желтая, изумрудно-зеленая , светло-коричневая, малиновая. Очень часто кончики ветвей несут голубой налет мелких молодых полипов. В ряде случаев к мадрепоровым кораллам прибавляются мягкие рифообразующие кораллы альционарии (Alcyonaria). Чаще других встречаются здесь виды рода саркофитум (Sarcophytum). Колонии саркофитум образуют обширные скользкие ковры площадью в несколько квадратных метров, сплошь покрытые сосковидными выростами. Когда идешь по такому ковру, ноги скользят и погружаются в податливую колонию. Цвет мягких кораллов обычно серый, мутно-зеленый или серо-фиолетовый. Некоторые виды, разрастаясь, ниже уровня отлива приобретают причудливую форму граммофонных труб с волнистыми краями, гигантских воронок, ваз или чаш. В прибойной части рифа поселяются также некоторые гидрокораллы, среди которых главное место занимает миллепора (Millepora), образующая густые сплетения кремово-желтых ветвей. Здесь же попадаются и ярко-голубые солнечные кораллы (Heliopora).
Очень богато и разнообразно население прибойной части рифа. В густых зарослях кораллов находят себе убежище и пищу многочисленные черви, моллюски, ракообразные, иглокожие, рыбы и многие другие животные. Часть из них всю жизнь прячется внутри колонии, иногда они просверливают ее скелет, в ряде случаев колония обрастает такое животное со всех сторон и оно оказывается навсегда замурованным в толщу коралла, получая пищу через небольшое отверстие. Другие укрываются в заросли только при опасности, третьи постоянно ползают по поверхности колоний или держатся вблизи от них. Приспособления животных к жизни на рифе также многообразны. Многие имеют толстый скелет или раковину, защищающую их от ударов волн. Такие организмы прочно держатся за риф при помощи крючковатых конечностей, присосок, острых выростов, крепких шипов или же просто прирастают к кораллам. Если эти животные подвижны, то они всегда медлительны. Их тело имеет обтекаемую форму, и вода не смывает их с рифа. Из крупных прикрепленных обитателей рифа нужно отметить двустворчатых моллюсков тридакна (Tridacna). Разные виды этого рода укрепляются на рифе различно. Некоторые из них прирастают к рифу при помощи мощного пучка биссуса — особых волокон, выделяемых специальной биссусовой железой. Раковина таких видов имеет очень толстые стенки и обычно достигает значительной величины. Живущая на рифах Индийского океана и Красного моря гигантская тридакна (Tridacna gigas) имеет раковину до полутора метров длиной при весе свыше 250 кг. В спокойном состоянии створки раковины приоткрыты и наружу выступает ярко окрашенный край мантии. Тридакна непрерывно пропускает через свои сифоны воду и улавливает пищу, которой ей служат планктонные организмы. Потревоженная тридакна быстро закрывает створки раковины. Ее мускул-замыкатель обладает очень большой силой, и случайное попадание руки или ноги ныряльщика в щель между створками грозит ему увечьем или гибелью. Маленькая, всего в несколько сантиметров длиной, тридакна кроцея (Tridacna сгосеа) живет в толще кораллового известняка и сообщается с внешней средой через узкую щель, из которой наружу выступают только края раковины и складки мантии. Очень любопытен брюхоногий моллюск верметус (Vermetus). Его раковина имеет вид длинной извитой трубки, приросшей к поверхности кораллов. Иногда колония обрастает вокруг всю раковину и наружу выступает только край устья. Верметус улавливает добычу, выделяя вокруг себя большое количество слизи. Время от времени моллюск поедает эту слизь вместе с прилипшими к ней мелкими организмами.
Подвижные моллюски, живущие на прибойной части рифа, имеют обычно крупные размеры и толстую раковину. Они прочно присасываются к поверхности рифа, а их тело даже при движении полностью покрыто раковиной, из-под которой выдаются лишь кончики щупалец. Если прибойная волна все же оторвет такого моллюска от рифа, он или глубоко втягивает мягкие ткани внутрь (так ведет себя пятнистый трохус — Trochus maculatus), или же закрывает вход в устье толстой сферической известковой крышечкой до 200 г весом (мраморный турбо — Turbo marmoratus). В самых прибойных местах рифа живет очень интересный морской еж гетероцентротус (Heterocentrotus mammilatus). Его тело имеет овальную форму, позволяющую ежу забираться в довольно узкие щели и небольшие полости рифа. Часть игл на спине животного сильно утолщена и служит для удерживания тела в расселинах. Основания игл снабжены мощной мускулатурой, позволяющей ежу прочно упираться в стенки своего убежища. Здесь же между ветвями и вдоль щелей медленно ползают округлые коротконогие крабы калаппа (Calappa) и атергатес (Atergates). Имеется много других крабов, моллюсков, иглокожих, обладающих толстыми защитными скелетными образованиями.
Другой способ защиты от разрушительного действия волн заключается в том, что тело приобретает предельную гибкость, оно изгибается и бьется вместе с волной, благодаря чему сохраняется его целость. Таковы поселяющиеся на прибойных рифах иглокожие животные морские лилии. Маленькое тело их прикреплено к рифу при помощи цирр — корневидных выростов нижней стороны тела, а длинные «руки» этого животного, имеющие членистое строение, свободно извиваются вместе с движущейся водой.
Все обитатели кораллового рифа ярко и пестро окрашены; это надежно скрывает их на разноцветном фоне кораллов.
Особой яркости достигают коралловые рыбки. Эти быстрые рыбки поодиночке и стайками «бродят» между кораллами, отщипывая полипов, которыми питаются. Некоторые из них обладают мощными челюстями, позволяющими им отгрызать и измельчать куски коралла. Если под водой разбить на мелкие куски колонию ветвистого коралла, то вскоре вокруг обломков собирается много рыбок, от крошечных до крупных в 20—30 см длиной. Все они набрасываются на обломки коралла и быстро растаскивают их в разные стороны. Большинство коралловых рыбок очень пугливы, они постоянно живут среди неподвижных кораллов и всякий шевелящийся предмет заставляет их искать убежище в зарослях.
Окраска и форма коралловых рыбок чрезвычайно разнообразны: здесь есть и красные, и зеленые, и голубые, и желтые, и черные. Почти все они имеют яркие пятна или полосы. В коралловых лесах укрываются хищные мурены, иногда достигающие 3 м в длину. Змеевидное тело мурены скрыто в щели или под нависающим кораллом, наружу выставлена маленькая голова с приоткрытым ртом, усаженным острыми, как бритва, зубами. Заметив добычу, мурена покидает засаду и стремительно нападает на свою жертву. В глубине пасти мурены имеются ядовитые зубы; попадание яда в кровь ускоряет гибель животного.
Совсем иначе выглядят коралловые поселения в бухтах и лагунах, защищенных от действия прибоя. В таких местах кораллы не образуют сплошного монолита, здесь они растут отдельными деревьями, кустами или лежат на дне, не прикрепившись к нему. Отсутствие волн способствует развитию нежных ветвистых колоний. Интересно отметить, что одни и те же виды при поселении на прибойной части рифа и в лагуне образуют колонии различной формы. Широко распространенная на всех коралловых островах поциллопора оленьи рога (Pocillopora damicornis) в прибойной зоне образует приземистые колонии с короткими толстыми веточками, а при поселении в лагунах этот вид образует редковетвистые колонии с тонкими длинными ветвями. Изменяется и цвет кораллов. Лагунные формы всегда окрашены менее интенсивно, здесь преобладают желтоватые, бурые и грязно-зеленые тона колоний. Виды, которые на прибойном рифе имеют интенсивный красный цвет, в лагунах окрашены в розовато-желтый.
Многие виды кораллов, обитающие на прибойном рифе, никогда не встречаются в лагунах; зато здесь имеются свои лагунные виды, которые могут жить только в спокойной воде. Очень своеобразны полушаровидные колонии эуфиллий (Euphyllia), иногда достигающие 1 м в диаметре. Каждая такая колония состоит из тесно расположенных ветвей — секторов, отходящих от центра полушария. Основания ветвей старых колоний разрушаются, после чего ветви теряют связь между собой. Большинство ветвистых колоний акропоры (Acropora), стилофоры (Stylophora) и других кораллов имеет длинные корневидные выросты, которые внедряются в ил, благодаря чему колония удерживается на месте. Веточки этих видов тонкие, чрезвычайно ломкие. Даже слабые волны обламывают кончики таких ветвей, однако поврежденные участки колонии быстро зарастают. В лагунах поселяются ячеистые колонии павона (Pavona), а также одиночные (не образующие колоний) грибовидные фунгии (Fungia). Молодые фунгии снабжены ножкой, на которой растет тело коралла. Оно похоже на шляпку сыроежки, обращенную пластинками вверх. Когда тело фунгии достигает в диаметре 2—3 см, ножка обламывается и коралл падает на дно, где продолжает расти. На ножке тем временем развивается новый коралл. Некоторые экземпляры достигают 10 см в диаметре, а вытянутая в длину колючая фунгия (Fungia echinata) вырастает до 30—35 см. Цвет фунгий обычно охристый или нежно-желтый, некоторые виды имеют розовый край.
Население лагунного рифа отличается своеобразием, здесь обитает много животных со слаборазвитым скелетом, с длинными ломкими придатками, а также мягкие, бесскелетные организмы. Встречается много плавающих и вообще быстро двигающихся животных. Живущие в лагунах крабы проворно плавают, их задние конечности уплощены и служат веслами. Плавают также и некоторые моллюски. Небольшой двустворчатый моллюск лима (Lima) передвигается, быстро хлопая створками, широкая мантия и длинные щупальца придают плавающей лиме внешнее сходство с медузой. Другой двустворчатый моллюск — гребешок (Chlammis), набрав воды в мантийную полость, выбрасывает ее резким толчком, при этом сам моллюск силой обратного толчка делает большой скачок. Многие моллюски, например молоточек (Malleus) и кренатула (Crenatula), подвешиваются на ветвистые колонии при помощи биссуса. На дне лагун попадаются жемчужницы (Pinctada), в илистом грунте живет большое количество различных закапывающихся двустворок. Среди них наиболее интересна пинна (Pinna), достигающая иногда 40 см. Раковина пинны, имеющая форму наконечника копья, погружена в ил на всю свою длину, причем нижний конец ее при помощи нитей биссуса прикреплен к обломку коралла. Наружу выступает лишь самый верхний край раковины. Брюхоногих моллюсков в лагунах обычно мало, однако нельзя не отметить попадающихся здесь различных ципрей (Cypraea), имеющих красивую глянцевую раковину. В ячейках, образованных колониями кораллов павона (Pavona), укрываются маленькие осьминоги.
В лагунах живет много разнообразных иглокожих — голотурий, морских ежей и звезд. Лагунные морские ежи имеют тонкий, ломкий панцирь. Встречаются также мягкие ежи, например токсопустес (Toxopneustes). Этот еж, достигающий величины кокосового ореха, сильно ядовит. «Укус» щипчиков (педициллярий), во множестве сидящих на теле ежа, вызывает сильнейшую боль и продолжительное сердцебиение.
Ядовиты и ежи диадемы (Diadema setosum). Величина тела такого ежа достигает размеров антоновского яблока, со всех сторон он покрыт почти полуметровыми тонкими иглами. Эти черные ежи сотнями и тысячами лежат на прогалинах между кораллами. Случайный укол их иглой также вызывает сильную боль, зависящую от попадания в ранку ядовитой слизи. В бухтах массами живут безвредные короткоиглые морские ежи трипнеустесы (Tripneustes), окраска которых подвержена значительной индивидуальной изменчивости. Встречаются и белые экземпляры, и особи с красным или оранжевым звездообразным рисунком. Лагунные морские звезды обычно достигают величины в размахе лучей 20—30 см. Они медленно ползают между кораллами, поедая моллюсков и других обитателей лагуны. Наиболее характерны для лагунного рифа неуклюжие кульциты (Culcita), похожие по форме, величине и цвету на круглый хлеб; очень обычны серо-зеленые с розовыми шипами звезды ореастеры (Oreaster) и коричневые, также шиповатые, протореастеры (Protoreaster).
В лагунах живут бурые очень крупные актинии Stoichactis kentii, диаметр ротового диска которых достигает полуметра. Вблизи каждой актинии постоянно держится пара маленьких креветок — самец и самка. В случае опасности креветки прячутся между щупальцами актинии.
И на прибойных рифах, и в лагунах, кроме животных, обитающих в зарослях кораллов и поселяющихся между их колониями, живет большое число организмов, еще более тесно связанных с мадрепоровыми кораллами. К ним относятся преимущественно мелкие животные: черви, иглокожие, моллюски и главным образом ракообразные. Все они обладают крючками, большими клешнями, выростами тела и другими приспособлениями, позволяющими им прочно удерживаться на ветвях кораллов. Окраска таких животных обычно бывает покровительственной. Так, все крабики и рачки, обитающие в красных колониях поциллопоры, сами окрашены в красноватый цвет. На бурых и коричневых кораллах поселяются темноокрашенные виды ракообразных.
В ячейках полипов некоторых видов мадрепоровых кораллов поселяются личинки усоногих рачков из рода пиргома (Pyrgoma). Полип вскоре погибает, а ячейка по мере развития рачка увеличивается в размерах, сохраняя основные пропорции. Такие уродливые ячейки похожи на галлы, вызываемые у растений насекомыми и клещами.
В толще полипняков поселяются разрушители кораллов — двустворчатые моллюски. Морские финики (Lithophaga) имеют почти цилиндрическую форму. Оны проделывают в полипняке трубчатые ходы, растворяя известь выделениями специальной железы. Раковина самого морского финика, также состоящая из извести, защищена от действия кислоты слоем органического вещества — конхиалина. Другие двустворчатые моллюски — фоласы (Pholas) протачивают коралл своей раковиной, поверхность которой покрыта многочисленными крепкими зубцами и похожа на напильник.
ПРАКТИЧЕСКОЕ ИСПОЛЬЗОВАНИЕ КОРАЛЛОВ
Как уже отмечалось, живые ткани мадрепоровых кораллов составляют незначительную часть веса этих животных, тогда как главная масса приходится на долю мощного известкового скелета. Скелет отлагается эктодермой кораллов, образующая его известь имеет особую некристаллическую форму (арагонит).
Для населения тропических стран коралловые рифы имеют чрезвычайно важное значение. Все обширное пространство, занятое коралловыми рифами, является огромной природной фабрикой извести. Изо дня в день, из года в год крошечные полипы мадрепоровых кораллов, а также другие морские животные извлекают известь из морской воды и откладывают ее в своем теле в виде скелета. Высокая температура воды в поверхностных слоях тропических морей способствует образованию массивного известкового скелета. Встречаются массивные колонии мадрепорового коралла Coeloria до 3—6 м в диаметре, а некоторые виды рода Pontes образуют колонии с диаметром 8—9 м. В тропической зоне моря известковый скелет отмерших организмов не подвергается разрушению. Напротив, в холодных: водах морские организмы, как правило, лишены массивного известкового скелета, который целиком или частично заменяется скелетом кремниевым или хитиноидным. Известь скелета отмерших организмов в холодной морской воде быстро разрушается. Известно, что мадрепоровые кораллы являются древними животными, и на местах их поселения в течение многих тысячелетий из скелетов отмерших колоний образовались значительные отложения извести, которые и в настоящее время продолжают увеличиваться. Благодаря тому что рифообразующие мадрепоровые кораллы поселяются у поверхности моря, т. е. по берегам островов, либо сами образуют острова, эта известь легко доступна для разработки, а запасы ее практически неограниченны, хотя и располагаются пятнами на местах, где были или имеются коралловые рифы.
Скелет мадрепоровых кораллов имеет довольно широкое хозяйственное применение. Во многих приморских тропических странах его используют как хороший строительный материал для постройки домов, молов, набережных, мощения улиц и т. д. Очень велика ценность мадрепоровых кораллов как сырья для получения извести. Кроме того, скелет мадрепоровых кораллов употребляют иногда для заполнения водопроводных фильтров, для полировки и шлифовки деревянных и металлических изделий, а также для изготовления лекарственных препаратов. Некоторые мадрепоровые кораллы вследствие красивой формы их колонии употребляются для устройства искусственных скал и гротов в садах и парках и в аквариумах, а также используются в качестве комнатных украшений.
Способы добычи кораллов в значительной мере зависят от места их обитания. В лагунах и бухтах, где дно обычно бывает заилено, колонии кораллов имеют форму кустов с редкими ломкими ветвями. Они не прикрепляются ко дну, а держатся на нем при помощи корневидных выростов, либо свободно лежат на поверхности ила. В мелководных лагунах, поросших кораллами, промысел их ведется с лодок. Кораллы собирают со дна при помощи трезубца на длинном древке. Вовремя прилива добытые кораллы подвозят возможно ближе к берегу и сваливают там в воду, а затем, когда наступает отлив, транспортируют на телегах к месту дальнейшей переработки.
Гораздо сложнее добывать кораллы из прибойных рифов, где они образуют массивные колонии, или на мертвых участках рифа, где из скелетов отмерших кораллов образовался известняк. Здесь материал добывают откалыванием кусков при помощи железных клиньев, ломов, молотов и других инструментов.
Массивные колонии мадрепоровых кораллов употребляют для постройки различных сооружений: изгородей, дамб, набережных, пристаней и т. д. Обработка строительного материала весьма несложная: края шаровидных колоний обкалывают, придавая им форму куба или бруса. Величина брусьев строго не определена, чаще она колеблется между такими размерами: 20—25—30 см и 25—30—35 см. Такие грубо обработанные брусья коралла укладывают прямой либо косой кладкой. С наружной, обычно необработанной поверхности стены хорошо различима структура колоний, часто позволяющая судить о родовой и даже видовой принадлежности коралла. В большинстве случаев отдельные брусья в кладке ничем не связаны и стена удерживается вследствие плотного прилегания их шероховатых поверхностей. Однако для придания постройке большей прочности глыбы коралла иногда скреплены раствором коралловой извести, о производстве и употреблении которой будет сказано несколько ниже.
Строительные брусья делают из массивных колоний кораллов, относящихся к родам Favia, Galaxea, Symphyllia, Coeloria, Porites, Goniastrea и другим.
Свежие и недавно отмершие кораллы — вполне подходящий материал для постройки различных сооружений на берегу моря (дамбы, пирсы, волноломы, набережные): они не разрушаются под химическим воздействием морской воды. Однако эти кораллы никогда не употребляют для постройки зданий. Причина этого заключается в значительной пористости материала, подверженного разрушению на воздухе и в результате действия пресной дождевой воды. Для наземных построек успешно применяют известняк, образовавшийся из отмерших кораллов. Для такого типа строений коралловый известняк колется на сравнительно небольшие узкие плитки (в среднем 8х15х20 см), которые укладывают друг на друга подобно кирпичам и связывают известковым раствором. Плитки, употребляемые для строительства зданий, почти лишены следов структуры колонии. Стена, сложенная из таких плиток, снаружи имеет приятную для глаза поверхность и не требует штукатурных и других отделочных работ. Изнутри же стены штукатурят.
Скелет всех мадрепоровых кораллов с успехом можно употреблять для обжига на известь. Сырьем служат как перечисленные выше массивные, так и ветвистые (Acropora, Pocillopora, Stylophora), ячеистые (Pavona) колонии и даже одиночные формы (Fungia), а также обломки любых кораллов. Вероятно, мадрепоровые кораллы обжигались на известь с древнейших времен, причем во многих местах сохранились очень примитивные методы обжига.
Химический состав кораллового известняка очень благоприятен для обжига его на известь. В высушенных мадрепоровых кораллах углекислый кальции составляет 90—97%, соединения фосфора и кремния содержатся в очень небольшом количестве (0,3—2,6%). Последнее очень важно, так как примесь солей кремниевой кислоты снижает качество извести.
Известно несколько типов печей для обжига кораллов. В наиболее примитивном случае печь складывают из самого обжигаемого материала, т. е. из ничем не связанных между собой и не обработанных колоний мадрепоровых кораллов. Стены печи укладывают в форме широкого кольца. Сверху печь открыта. Обычно диаметр такой печи около 10 м, высота стен 2 м. Стены печи для предохранения их от разваливания подпирают снаружи деревянными кольями. Вследствие неплотного прилегания кусков коралла, образующих стену печи, между ними остаются большие отверстия, через которые при обжиге поступает воздух. Если поддув оказывается избыточным (при сильном ветре), стены печи прикрываются плотным плетнем и циновками из листьев кокосовой пальмы. Печь загружают дровами и засыпают несколькими тоннами разбитого коралла (один кусок обычно не превышает по весу 1—1,5 кг). Разжигают печь сверху через отверстие в засыпанных кораллах. Обжиг производится в течение четырех-пяти суток. Затем около трех суток печь остывает. После остывания ее разгружают, для чего разбирают часть стены и выносят известь через образовавшийся проход. Затем обожженный материал просеивают на грохоте. Такие примитивные печи устанавливают поблизости от места добычи коралла и после окончания разработок разрушают.
Более широко используют маленькие постоянные печи. Такая печь имеет форму полого, открытого сверху цилиндра или усеченного конуса, обращенного вверх основанием. Печь ставят на кирпичном фундаменте, она имеет снизу поддувало. Под печи снабжен колосниками. Стены складывают из ничем не связанного кирпича и для предохранения от рассыпания оплетают снаружи гибкими ветвями, придающими печи сходство с гигантской корзиной около 2 м в диаметре. На дно печи укладывают слой топлива, затем слой мелко битого коралла, далее снова топливо и т. д. Всего кладется 3—4 слоя кораллов. Процесс обжига длится около 15 часов. Обожженные кораллы извлекают из печи через широкое отверстие в ее стене.
Гораздо более продуктивны печи непрерывного действия. Такая печь имеет вид высокой башни и построена из кирпича. Топливо и коралловый известняк загружают через окно в верхней части печи. В качестве топлива используют каменный уголь. Выем обожженной извести производят через окна в нижней части печи. Интенсивность горения регулируют искусственным поддувом от механической воздуходувки. Для усиления тяги на верху печи установлена труба. По мере выема извести слои горящего угля и обжигаемых кораллов под влиянием своего веса опускаются из верхней части печи в среднюю, где и происходит обжиг. В нижней части печи обожженная масса остывает. Отсюда она и извлекается, а через верхнее окно в печь добавляют новые порции топлива и сырья. Печь работает непрерывно.
При использовании в качестве топлива каменного угля обожженная известь получается более твердой, чем при топке печи дровами. Поэтому ее приходится размалывать на шаровой мельнице.
Готовую известь используют в строительном деле для приготовления связывающих растворов и штукатурных работ. Качество ее очень высокое. Готовый продукт имеет вид чисто-белого жирного на ощупь порошка, состоящего почти из одной окиси кальция.
Как видно из сказанного, мадрепоровые кораллы используют главным образом для получения строительных материалов, однако этим применение скелета кораллов не ограничивается.
Благодаря пористому строению скелет мадрепоров оказался очень удобным материалом для заполнения водопроводных фильтров. Теперь в большинстве стран, расположенных у берегов тропических морей, фильтры заполняют не коксом, как это делали раньше, а хорошо промытыми кусками кораллов.
Известь, из которой построен скелет мадрепоровых, очень легко измельчить в тонкий порошок, который с успехом применяют как полировочный материал. Куском коралла, как напильником, можно обрабатывать деревянные изделия.
На островах Самоа из пластинок кораллового известняка изготовляют блесны для ловли рыбы.
Как уже было сказано, в тропиках имеется немало островов, возникших из отмерших коралловых рифов. На них вообще нет и не может быть природного камня, а потому здесь кораллы употребляются как тяжелые предметы. При их помощи раздавливают плоды для получения сока, перетирают семена. Куски коралла употребляют как балласт в лодках и как груз в различных механических приспособлениях.
Издавна не только в тропиках, но и в Европе мадрепоровым кораллам приписывали магические свойства, из них делали амулеты, будто бы предохранявшие от «дурного глаза» и колдовства.
В старину в Европе скелет мадрепоровых кораллов под названием «кораллиум альбум» служил для приготовления некоторых лечебных препаратов. В некоторых странах Юго-Восточной Азии и сейчас толченый скелет этих кораллов можно найти в любой аптеке.
Довольно большое значение имеет употребление мадрепоровых кораллов в качестве украшений и сувениров. Для этой цели используют преимущественно ветвистые колонии (Acropora, Pocillopora), похожие на причудливое растение тридакофиллию (Tridacophyllia), а также на одиночные грибовидные кораллы (Fungia).
Ловцы собирают особенно красивые колонии, выбирая их среди зарослей. Отобранную колонию осторожно отделяют от рифа и затем удаляют из нее мягкие ткани. Известно несколько способов отделения мягких тканей от скелета. Если на судне имеется водяная помпа, то проще всего разложить собранные кораллы на палубе и поливать их из шланга струей воды под большим напором. При этом нежные ткани легко смываются и остается белоснежный известковый скелет, который затем высушивается на тропическом солнце. Более простой и, вероятно, самый древний способ отбеливания кораллов состоит в том, что собранные колонии привязывают одну за другой на длинную веревку и тянут ее за парусной лодкой. При этом мягкие ткани также смываются струями воды. Нужно только следить, чтобы кораллы не опускались на дно, где они неминуемо поломаются. Когда лодка пройдет несколько миль, веревку с отбеленными кораллами вытаскивают на борт. Наконец, мягкие ткани могут быть удалены продолжительным вымыванием кораллов в пресной воде. Выбеленные и просушенные скелеты мадрепоровых кораллов очень твердые, хотя и хрупкие. Они звенят, как хороший фарфор, да и внешне кажутся сделанными из чистейшего неглазированного фарфора.
В качестве украшений и сувениров кораллы наравне с раковинами тропических моллюсков охотно покупаются и местным населением и приезжими. В Китае и Вьетнаме из мадрепоровых кораллов делают декоративные горки и помещают их в садах и парках. Иногда перед продажей белые скелеты окрашивают яркими красками.





