(Hydrophiidae) Семейство Гидрофииды, Морские змеи, Настоящие морские змеи, Family Hydrophiidae
Жизнь животных. Том 4. Часть 2. Земноводные, пресмыкающиеся Под редакцией профессора А.Г.Банникова 1969 г.
ПОДОТРЯД ЗМЕИ (OPHIDIA, ИЛИ SERPENTES)
Змеи — одни из самых своеобразных существ на земле. Их необычный внешний вид, оригинальный способ движения, многие замечательные особенности поведения, наконец, ядовитость многих видов — все это издавна привлекает внимание и вызывает живой интерес у людей. У самых различных народов земного шара сложено множество легенд, сказок и мифов о змеях. Все эти фантазии, подкрепленные подчас безотчетным суеверным страхом перед змеями, так тесно переплелись с действительными фактами, что многие «правдивые» рассказы о змеях бывают гораздо фантастичнее любых мифов. Изучение змей постепенно разоблачает легенды и в то же время открывает новые замечательные черты в строении и образе жизни этих животных.
На первый взгляд кажется, что змей легко отличить по внешнему облику от всех других пресмыкающихся. Действительно, они имеют длинное, лишенное ног туловище, одетое чешуей, глаза их всегда покрыты прозрачной кожистой оболочкой, у них отсутствует наружное ухо. Однако все эти черты строения могут встречаться также у различных ящериц. Ящерицы и змеи — близкородственные животные, поэтому их относят лишь к разным подотрядам внутри общего о тряда чешуйчатых (Squamata). Есть такие «промежуточные» семейства, по поводу которых ученые до сих пор сомневаются, куда относить их — к змеям или к ящерицам. Около тридцати признаков внешнего и внутреннего строения отличает змей от яшериц, но почти все они «как исключение» встречаются и у последних. Таким образом, только по комплексу всех этих отличий можно достоверно разделить два подотряда чешуйчатых пресмыкающихся.
В строении черепа змей мы находим наиболее характерные и устойчивые признаки этих животных, отличающие их от ящериц. Они обеспечивают исключительную растяжимость рта змей, что позволяет им заглатывать целиком добычу, которая значительно толще их тела. Кости лицевой части черепа змей подвижно соединены между собой, а нижняя челюсть подвешена к черепу на сильно растяжимых связках. Эластичная связка соединяет также правую и левую половины нижней челюсти. Кроме того, мозг змей целиком заключен в костную капсулу, и межглазничная перегородка не развита.
Семейство Морские змеи (Hydrophidae)
Высокая специализация к водному, морскому образу жизни, вне связи с сушей, характерна для подавляющего большинства представителей этого семейства змей. Морские змеи имеют несомненные общие корни с аспидовыми, что проявляется в ряде черт их организации, но переход в водную среду наложил такой глубокий отпечаток на строение и биологию этих змей, что они заслужили выделения в особое семейство.
Облик морских змей очень характерен — маленькая голова с небольшими глазами, имеющими круглый зрачок, покрыта крупными щитками и плавно переходит в туловище. Туловище впереди вальковатое, а в задней части сплющенное с боков и переходящее в широкий и плоский ластообразный хвост. Примечательна редукция или полное отсутствие расширенных брюшных щитков, которые были важны при движении по твердому субстрату, но оказались помехой при плавании: тело большинства морских змей и сверху, и снизу покрыто одинаково мелкими чешуйками. Лишь наиболее примитивные роды (Laticauda, Aipysurus, Emydocephalus, Ephalophis), сохранившие связь с сушей, еще имеют на брюшной стороне ряд поперечно расширенных щитков.
Ноздри морских змей выдвинуты на верхнюю сторону морды, что позволяет им беспрепятственно дышать, высунув из воды лишь самый кончик носа. Кроме того, ноздри замыкаются специальными клапанами, которые защищают носовую полость от воды при нырянии.
В морской воде язык змеи уже не может выполнять свою обонятельно-осязательную роль, он у морских змей сильно укорочен и лишь раздвоенный кончик его способен высовываться наружу изо рта. Слизистая оболочка ротовой полости богата капиллярными кровеносными сосудами и может поглощать кислород непосредственно из воды. Этот дополнительный орган водного дыхания позволяет змее подолгу находиться под водой.
Парные ядовитые зубы расположены на переднем конце верхнечелюстной кости. Они сравнительно коротки, слегка изогнуты назад и имеют ядопроводящий канал. Позади ядовитых клыков на верхней челюсти находятся мелкие зубы, число которых сильно варьирует — от 1 у плоскохвостов (Laticauda) до 18 у голубого ластохвоста (Hydrophis caerulescens). Только у кольчатого эмидоцефала (Emydocephalus annulatus) позади ядовитых зубов совсем нет маленьких зубов.
Яд у многих видов морских змей в несколько раз превосходит по токсичности яды самых ядовитых наземных змей. Но при укусе морские змеи вводят в жертву очень малое количество яда — 1—20 мг в сухом весе (0,06—0,12 мл жидкого яда). Кроме того, большинство морских змей не склонно кусаться без особого побуждения, так что индийские рыбаки при попадании змей в сети часто вытаскивают их оттуда голыми руками, не подвергаясь укусам. Если же змея схвачена очень резко и ей причиняется боль, то она может укусить. Яд морских змей оказывает нейротоксическое воздействие, без опухоли и воспаления в месте укуса. Развиваются общие явления — слабость, нарушение координации движений, рвота, судороги, затруднение дыхания и далее полный паралич дыхательного центра. Если меры по оказанию первой помощи не были приняты, то через 5—10 часов человек может погибнуть. Однако процент летальных исходов при укусах невелик, благодаря тому что дозы яда, вводимые при укусе, обычно очень малы. Высокая токсичность яда у морских змей выработалась как приспособление к питанию пойкилотермными животными, сравнительно устойчивыми к ядам. Кроме того, в дебрях коралловых зарослей необходимо почти моментальное умерщвление добычи, иначе она забьется в такие щели, откуда ее уже не достать. Добавим, однако, что некоторые виды морских змей имеют весьма слабо действующий яд.
Пищей морским змеям служат различные рыбы, в первую очередь угри, в частности змеевидные угри рода Ophichthys. Помимо активной охоты, морские змеи применяют иногда своеобразную охоту на приманку. Змея неподвижно распластывается на поверхности воды и ждет, когда любопытные рыбешки соберутся вокруг нового предмета. Резкое движение — и одна из рыб оказывается схваченной.
Размножение у морских змей происходит разными способами. Немногие виды, тесно связанные с сушей, яйцекладущи. Подавляющее большинство яйцеживородящи, и у многих видов имеется примитивная плацента, связывающая зародыш с материнским организмом. Некоторые змеи рождают детенышей прямо в воде, а другие выходят на берег, чтобы принести потомство на суше. Плодовитость их удивительно низка — самки рождают всего по 1—2 детеныша. Половая зрелость наступает обычно в возрасте 1 года.
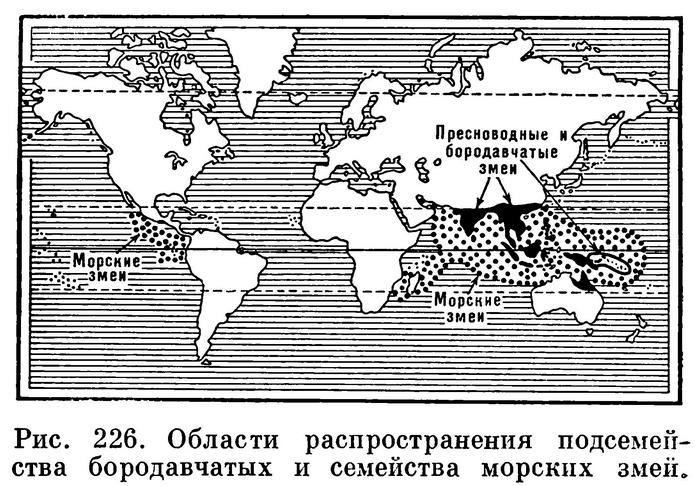
Населяют все тропические моря Тихого и Индийского океанов, от восточного побережья Африки до западных берегов Центральной Америки. На север они проникают до Японии. Наиболее многочисленны по числу видов и особей морские змеи в Южно-Китайском море и в водах Малайского архипелага. Здесь они часто встречаются большими скоплениями и десятками попадаются в рыбацкие сети. Излюбленные места обитания змей — прибрежные воды в полосе до 5—6 км от берега, особенно близ устьев крупных рек, где змеи находят изобилие пищи в виде рыб. Лишь изредка они заплывают дальше 50 км от ближайшей суши.
Известно 48 видов морских змей, объединяемых в 16 родов. Три рода (12 видов) образуют подсемейство плоскохвостых (Laticaudinae), а 13 родов (36 видов) группируются в подсемейство ластохвостых (Hydrophinae).
Плоскохвостые морские змеи еще сохранили тесную связь с сушей и нередко встречаются вдали от воды. У них хорошо развиты брюшные щитки, что позволяет им успешно передвигаться по твердому субстрату. По-видимому, большинство видов этого подсемейства яйцекладущи (установлено пока лишь у двух видов).
Центральный род подсемейства — плоскохвост (Laticauda) включает 4 вида, обитающих в западной части Тихого и восточной части Индийского океана. Плоскохвосты обычно бывают длиной не более 1 м, лишь один вид — большой плоскохвост может достигать почти 2 м длины при толщине в 7—8 см. Туловище плоскохвостов покрыто гладкой, черепицеобразно налегающей чешуей, голова одета крупными правильными щитками. Ноздри расположены на боковой поверхности головы, в отличие от остальных морских змей.
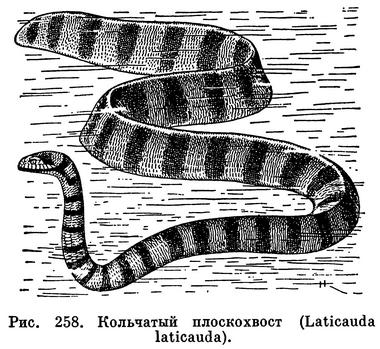
Кольчатый плоскохвост (Laticauda laticauda) — наиболее обычный вид, он широко распространен от островов Рюкю до Австралии и от Индии до Соломоновых островов. Туловище имеет яркий голубоватый фон, по которому чередуются широкие черные кольца. Брюшная сторона окрашена в более светлые голубовато-желтые тона. Эта красивая змея в больших количествах встречается на коралловых рифах, в прибрежной растительности и среди выбросов в полосе прибоя. Далеко от берега она не заплывает, зато нередко встречается на суше.
Большой плоскохвост (Laticauda semifasciata), достигающий длины почти 2 м, распространен в западной части Тихого океана, от островов Рюкю до островов Самоа. Эта крупная змея многочисленна в прибрежных водах Филиппин и издавна служит объектом промысла японских рыбаков. Они добывают змей ради шкуры, а также везут их живьем в Японию, где эти змеи идут в пищу в копченом или жареном виде.
Озерный плоскохвост (Laticauda crockeri) имеет очень ограниченную область распространения. Он встречается только в солоноватом озере Тунгано на острове Реннелл — самом южном из группы Соломоновых островов. Любопытно, что в этом озере обитает также и кольчатый плоскохвост (особый подвид).
Кроме плоскохвостов, в это подсемейство входят кольчатый эмидоцефал (Emydocephalus annulatus), распространенный от островов Рюкю до австралийского побережья, и 7 видов рода Aipysurus, населяющие воды Малайского архипелага, Юго-Восточной Азии и Северной Австралии.
Второе подсемейство морских змей — ластохвостые (Hydrophinae). Они неразрывно связаны с водной средой и насколько свободно и ловко движутся в воде, настолько же беспомощны и неуклюжи на суше. По твердому субстрату они практически не могут передвигаться из-за отсутствия расширенных брюшных щитков. Только у одного вида (Ephalophis greyi) имеются крупные брюшные щитки. Всю свою жизнь ластохвостые змеи проводят среди морских волн, и лишь некоторые виды выбираются на берег, чтобы принести потомство. Все они яйцеживородящи.
Более 20 видов этого подсемейства относится к роду ластохвостов (Hydrophis). В облике ластохвостов характерна маленькая голова и тонкая передняя часть туловища, резко отличающаяся от утолщенной задней половины и мощного ластообразного хвоста. Такое строение тела удобно при добывании пищи в толще воды, где нет опоры на твердый субстрат. Во время броска, схватывания и глотания жертвы тяжелый задний отдел служит основной опорной массой и движения передней части тела оказываются более экономными, чем при равномерном распределении массы по длине туловища. Наиболее выражен такой склад тела у ленточного ластохвоста (Hydrophis fasciatus). Брюшная часть тела у него в 4—5 раз толще, чем передняя. Спереди он черный, с овальными желтыми пятнами по бокам, а на задней части туловища по серовато-желтому фону — темные ромбы. Он водится в Индийском океане и в водах Малайского архипелага.
Полосатый ластохвост (Hydrophis cyanocinctus), окрашенный в зеленовато-серый цвет с черными поперечными полосами, достигает длины 1,5—2 м. Он обычен в Индийском океане, среди островов Малайского архипелага и проникает на север до Японии.
Спиральный ластохвост (Hydrophis spiralis), распространенный сходно с предыдущим видом, выделяется своей значительной длиной (до 2,7 м). Немного уступает ему по величине изящный ластохвост (Hydrophis elegans), обитающий в водах Северной Австралии и у островов Ару. Длина крупных особей превышает 2 м. Остальные виды ластохвостов, обитающих в Индийском океане и морях Юго-Восточной Азии, более мелких размеров (до 1—1,5 м).
Замечательно распространение лусонского ластохвоста (Hydrophis semperi), который живет в пресноводном озере Тааль на юге острова Лусон (Филиппины). Озеро имеет около 20 км в поперечнике и расположено в кратере потухшего вулкана. Лусонский ластохвост — единственная морская змея, приспособившаяся к жизни в пресной воде.
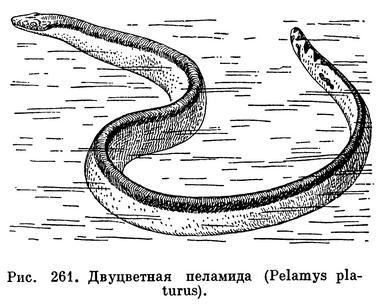
Двуцветная пеламида (Pelamis platurus) — небольшая змея, длиною до 1 м, с удлиненной, приплюснутой сверху головой и утолщенной шеей, сжатым с боков телом и плоским ластообразным хвостом. Шестиугольные прилегающие чешуи равномерно покрывают все туловище. Темно-бурая, почти черная, окраска верхней стороны обычно контрастирует со светло-желтой окраской низа, и на боках эти два цвета резко, без плавного перехода сменяют друга друга. Хвост покрыт крупными темными пятнами по светлому фону. Окраска пеламиды может варьировать, и в разных частях ее обширного ареала встречаются особи с продольными боковыми полосами или многочисленными пятнами и поперечными кольцами по всему туловищу.
Двуцветная пеламида распространена необычайно широко — от восточного побережья Африки через Индийский и Тихий океаны до западного побережья Америки. Ее ареал перекрывает области распространения всех остальных морских змей. Она заходит на юг до мыса Доброй Надежды, проникает на север до Японского моря. В нашей стране мертвый экземпляр этой змеи был найден однажды в заливе Посьета, южнее Владивостока. Вдоль побережья Америки пеламида встречается от Северо-Западной Мексики до Перу. Она не ограничивается прибрежными районами моря, заплывает далеко от суши и нередко попадается в сотнях километров от ближайшей земли. Поэтому ее можно с полным правом назвать пелагическим животным. Именно пелагический образ жизни позволил змее заселить столь обширный ареал.
В Индийском океане, от Персидского залива до Малайского архипелага, обитают два вида энгидрин (Enhydrina). Эти змеи, длиной около 1 м, очень многочисленны в восточной части ареала и часто держатся большими скоплениями. Опыты показали, что яд энгидрин намного превосходит по токсичности яд индийской кобры. Энгидрины десятками попадают в рыбацкие сети, но когда рыбаки вытаскивают их из сетей, они, как правило, даже не пытаются укусить. Известен случай содержания энгидрины в неволе. Она все время держалась на воде, не подплывая к берегам аквариума. Для отдыха прислонялась к плавающим кускам дерева, но не взбиралась на них. Оптимальная температура воды для содержания этой змеи 24°; при 20° она уже перестает принимать пищу. Вначале энгидрину кормили живой рыбой, но затем она приучилась поедать нарезанные кусочки сырого рыбного филе.
Многие морские змеи встречаются значительными скоплениями, но всех превзошла в этом отношении астроция (Astrotia stokesii), ярко-красная с черными кольцами змея длиной до 1,5 м. Одному зоологу удалось встретить в Малаккском проливе гигантское скопление астроций длиной около 100 км при средней ширине в 3 м. Плотная темная лента из многих тысяч тесно сплотившихся змеиных тел тянулась до самого горизонта. Причины подобных скоплений неясны, вероятно, это связано с периодом массового размножения.
Крайней степени приспособления к обитанию в море достигли микроцефалы. Наиболее обычен стройный микроцефал (Microcephalophis gracilis), населяющий Индийский и западные области Тихого океана. Небольшая змейка, длиной 70—80 см, с крошечной головкой, очень тонкой шеей и передней частью тела и непомерно утолщенной задней половиной. Все тело змеи покрыто одинаковыми шестиугольными чешуями с плотно прилегающими краями. Самка микроцефала приносит одного детеныша, который к моменту рождения достигает длины около 35 см (половина длины матери) и вполне способен к самостоятельной жизни в водной стихии. Хотя каждый приплод и невелик, но зато половая зрелость у микроцефалов наступает очень рано — в возрасте 6—8 месяцев.
Семейство Аспидовые змеи (Elapidae)
Обширное семейство аспидовых змей содержит около 180 видов, объединяемых в 41 род. Все виды этого семейства ядовиты. Парные ядовитые зубы помещаются на переднем конце заметно укороченной верхнечелюстной кости, они значительно крупнее остальных зубов, загнуты назад и снабжены ядопроводящим каналом. Строение этого канала в наиболее типичном варианте наглядно демонстрирует происхождение его от бороздки на передней поверхности зуба: передняя стенка канала образована как бы сомкнувшимися краями бороздки и на поверхности зуба заметен «шов», под которым расположен канал. Однако ядовитые зубы аспидовых змей еще примитивны, поскольку они располагаются в полости рта неподвижно.
У наиболее примитивных австралийских видов аспидовых змей еще по 8—15 мелких зубов расположено на верхней челюсти, у большинства аспидовых число этих зубов уменьшено до 3—5, а у африканских мамб и американских аспидов на верхней челюсти уже нет никаких зубов, кроме парных загнутых назад ядопроводящих клыков.
Обычно на каждой верхнечелюстной кости имеется по 2 таких клыка, лежащих бок о бок, но лишь один из них функционирует в данное время, а другой является «заместителем», вступающим в действие при утрате первого. Змеи периодически теряют свои ядовитые зубы, и им на смену вырастают зубы-заместители, так что змеи надежно обеспечены своим грозным оружием. Помимо верхнечелюстной кости, нёбная, птеригоидная и зубные кости также снабжены мелкими зубами.
В скелете аспидовых мы не находим уже никаких рудиментов таза и задних конечностей. Левое легкое у этих змей отсутствует.
Голова покрыта крупными щитками, причем характерно для всех аспидовых отсутствие скулового щитка (этот признак, однако, встречается у отдельных представителей других семейств). У подавляющего большинства аспидовых голова спереди закругленная, плавно, без шейного перехвата, переходящая в туловище, глаза с круглым зрачком. Лишь у отдельных уклонившихся видов (например, у австралийской смертельной змеи) голова имеет треугольную форму и отграничена резким шейным перехватом. Спинная чешуя гладкая, нижняя сторона тела змеи покрыта сильно расширенными брюшными щитками. По стройному телосложению, гладкой чешуе и большому щитку головы многие аспидовые внешне весьма сходны с ужеобразными змеями. Поэтому нередко аспидовых называют также ядовитыми ужами. Однако такое название вносит изрядную путаницу, так как среди самих ужеобразных тоже есть немало ядовитых видов.
Окраска тела довольно разнообразна, но наиболее типичны два варианта. Крупные наземные и древесные формы (кобры, мамбы и др.) имеют однородную или с неясным рисунком серую, песочную, бурую или зеленую окраску туловища. Более мелкие роющие формы (коралловые и украшенные аспиды) имеют яркий, контрастирующий рисунок тела, состоящий из чередования красных, желтых, черных колец.
Распространены в тропических и субтропических областях всех материков (исключая Европу) и достигают наибольшего богатства и разнообразия форм в Австралии и Африке. Австралию населяют наиболее древние и примитивные виды аспидовых. Поскольку на этот материк не смогли проникнуть более молодые семейства ядовитых змей — гадюковые и ямкоголовые, аспидовые заняли здесь различные экологические ниши. Эволюция аспидовых на этом материке, свободном от других ядовитых змей, привела к созданию видов, внешне очень схожих с гадюками и ямкоголовыми змеями (например, гадюкообразная смертельная змея — Acanthophis antarcticus). Этот процесс называется конвергентной адаптацией (он хорошо известен для австралийских сумчатых, которые в отсутствие высших млекопитающих образовали здесь аналогичные им формы — сумчатых волков, белок, крыс и т. п.). Большое количество родов (22) свидетельствует о длительной истории аспидовых в Австралии.
В Африке также находится древний центр распространения аспидовых, однако в сравнении с Австралией здесь обитают более молодые и прогрессивные виды. Экологически африканские аспидовые очень разнообразны (10 родов, 21 вид).
Фауна аспидовых в Азии представлена эволюционно молодыми и сравнительно специализированными формами (6 родов, 31 вид). Наибольшее число видов образуют здесь крайты и украшенные аспиды. В Азии обитает и самая крупная из всех ядовитых змей — королевская кобра. Экологическое разнообразие аспидовых здесь относительно невелико: преобладают наземные и роющие виды.
Америка была заселена аспидовыми позже других материков, и процесс видообразования здесь находится пока на первой ступени (51 вид, объединяемый всего в 3 рода). Коралловые аспиды Америки — группа очень однородная по своей морфологии и экологии. Их зубной аппарат высокоспециализирован: верхнечелюстная кость очень укорочена и на верхней челюсти имеются только парные ядовитые зубы. Все коралловые аспиды в большей или меньшей степени роющие змеи.
Питание аспидовых змей разнообразно; у многих обнаруживается пристрастие к змеям (в основном к мелким и неядовитым видам), в остальном же рацион их состоит из мелких млекопитающих, рептилий и земноводных, реже из птиц и беспозвоночных.
Яд у аспидовых змей состоит из многих компонентов различного действия и неодинаков по составу у разных видов. Однако в целом среди действующих начал в яде аспидовых резко преобладают нейротоксины, что вызывает при укусе характерную клиническую картину. Местные явления в области укуса почти не развиваются (нет ни опухания, ни красноты), зато быстро наступает смерть вследствие угнетения нервной системы, в первую очередь паралича дыхательного центра.
Преобладают яйцекладущие, но немало и яйцеживородящих видов. Живорождение свойственно главным образом роющим формам, а также большинству австралийских аспидовых. У некоторых яйцекладущих видов (например, у королевской кобры) кладка яиц охраняется самкой.
Ложные аспиды (род Aspidomorphus) — одни из самых примитивных змей этого семейства. На длинной верхнечелюстной кости у них за ядовитыми клыками расположено 8—12 мелких зубов. Семь видов этих небольших, длиною до 1 м, змей обитает в Северной и Западной Австралии, а один вид (Aspidomorphus muelleri) — на Новой Гвинее и прилежащих островах. Яд у ложных аспидов очень слабый, и соответственно своим маленьким размерам они охотятся в основном за насекомыми.
Обширный род денисоний (Denisonia) содержит 19 видов, встречающихся по всему материку.
Великолепная денисония (Denisonia superba), до 1,5 м длины, обитает в густонаселенных районах Юго-Западной Австралии и представляет известную опасность для человека и домашних животных. Самка денисонии рождает до 40 детенышей. При этом замечательно, что у нее имеется упрощенное подобие плаценты, связывающей кровеносные системы зародышей и матери.
Шесть видов коричневых змей (род Demansia) широко распространены по всей Австралии, а также проникают на Новую Гвинею и другие острова. Зубная система у коричневых змей очень примитивна — позади ядовитых клыков на удлиненной верхнечелюстной кости расположено от 7 до 15 мелких зубов. Все коричневые змеи яйцекладущи.
Самый обычный вид — сетчатая коричневая змея (Demansia textilis) достигает длины свыше 2 м и обитает в засушливых местностях по всему материку. Молодые особи имеют яркие поперечные кольца на туловище, а взрослые окрашены однотонно. Пища этой змеи состоит из ящериц и мелких млекопитающих. Самки откладывают по 15—30 яиц, из которых через 2 месяца выходят молодые.
Песчаная коричневая змея (Demansia psammophis) заметно мельче предыдущего вида, в длину не более 1,5 м. Сверху эта змея окрашена в серовато-бурый, а снизу в желтый цвет. Она населяет сухие каменистые местообитания, охотится главным образом за ящерицами и ведет дневной образ жизни. Яд у этой змеи не очень сильный — укушенные ящерицы длиной 15—18 см погибают примерно через 10 минут.
Черная змея, или черная ехидна (Pseudechis porphyriacus), распространенная по всей Восточной и Южной Австралии, достигает в длину 1,5—2 м. Блестяще-черная окраска верхней стороны тела эффектно сочетается с красноватой окраской брюха. Черная змея держится в умеренно влажных низменных местах и по долинам рек, охотно идет в воду, хорошо плавает и ныряет. Питается лягушками, ящерицами, змеями. Молодые особи предпочитают насекомых и других беспозвоночных. В неволе черная змея хорошо ест мышей. Будучи чем-либо потревожена или раздражена, черная змея слегка разводит в стороны шейные ребра, уплощая и расширяя шею. Самцы черной змеи часто вступают в турнирное единоборство между собой. Подняв голову и изогнув шею, они наступают друг на друга, пытаясь накрыть своей головой голову противника. Когда одному из соперников удается это сделать, он резким движением обвивает туловище противника своим телом. Яростно шипя и извиваясь, обе змеи сдавливают друг друга.
Вдруг, как по сигналу, они прекращают борьбу и расходятся, чтобы приготовиться к следующему поединку. Каждый из таких «раундов» длится около минуты, и они повторяются до полного изнеможения борцов. Змеи бывают так увлечены турниром, что не распутываются, даже если их поднять с земли. Причиной таких боев служит, очевидно, территориальный инстинкт в сочетании с половым возбуждением. Характерно, что во время турнира соперники не наносят друг другу укусов.
На северо-востоке Австралии и на Новой Гвинее обитает самая опасная из австралийских змей — тайпан (Oxyuranus scutellatus). Размеры тайпана очень внушительны — до 3—3,5 м, а ядовитые зубы длиной более сантиметра награждают жертву при укусе солидной дозой яда. По количеству и силе яда тайпан превосходит всех змей Австралии, лошадь от его укуса погибает через несколько минут, и несколько известных случаев укуса людей тайпаном неизменно кончались их гибелью. Тайпан очень агрессивен: при виде опасности он скручивается, уплощает тело, вибрирует концом хвоста и, высоко подняв переднюю часть туловища, делает несколько выпадов в направлении врага. К счастью местных жителей, тайпан — довольно редкая змея, встречающаяся в малонаселенных районах.
Тигровая змея (Notechis scutatus) уступает по величине тайпану, достигая всего 1,5—2 м длины, но яд ее очень силен; считается, что у тигровой змеи наиболее сильный яд среди всех наземных змей. Опасность встречи с этой змеей усугубляется еще и тем, что она широко распространена почти по всему материку, кроме самых северных областей, и населяет также Тасманию и ряд островов у южного побережья. Черное туловище змеи перехвачено нерезкими сернисто-желтыми кольцами, а брюхо имеет желтую окраску. В возбужденном состоянии тигровая змея высоко поднимает переднюю часть туловища, сильно уплощая голову и шею. Укушенные тигровой змеей мелкие животные погибают моментально, буквально не сходя с места. Подсчитано, что яда, содержащегося в железах крупной тигровой змеи, достаточно, чтобы убить 400 человек. Яйцеживородяща и приносит обильное потомство — обычно до 72 змеенышей. (Известен случай, когда у крупной самки нашли при вскрытии 109 зародышей.)
Смертельная змея (Acanthophis antarcticus) примечательна большим внешним сходством с гадюками. Ее широкая голова с выступающими скулами имеет треугольную форму с резким шейным перехватом, надглазничный щиток резко выступает вбок, чешуи на верхней стороне туловища снабжены ребрышками. Тело ее короткое и кургузое, и ведет себя она также подобно гадюке. При появлении опасности змея лежит неподвижно, не спасаясь бегством и не принимая отпугивающей позы, а полагается на свою покровительственную окраску, делающую ее незаметной. В связи с такой манерой поведения путник чаще сталкивается вплотную со змеей и бывает укушен ею. Яд смертельной змеи втрое слабее, чем тигровой змеи, и по размерам она заметно мельче. Но все-таки половина укушенных этой змеей людей погибает. Широкое распространение (Австралия, Новая Гвинея и соседние острова) усугубляет приносимый ею вред.
В Центральной и Западной Австралии распространена близкая к предыдущему виду огненная змея (Acanthophis pyrrhus), имеющая ярко-красный цвет туловища.